
Characterization and Control of Dendrobium officinale Bud Blight Disease

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, Pathogens Isolation and Purification
2.2. Pathogenicity Tests of Isolates
2.3. Identification of Pathogens
2.3.1. Morphological Observation
2.3.2. DNA Extraction, Amplification and Sequencing
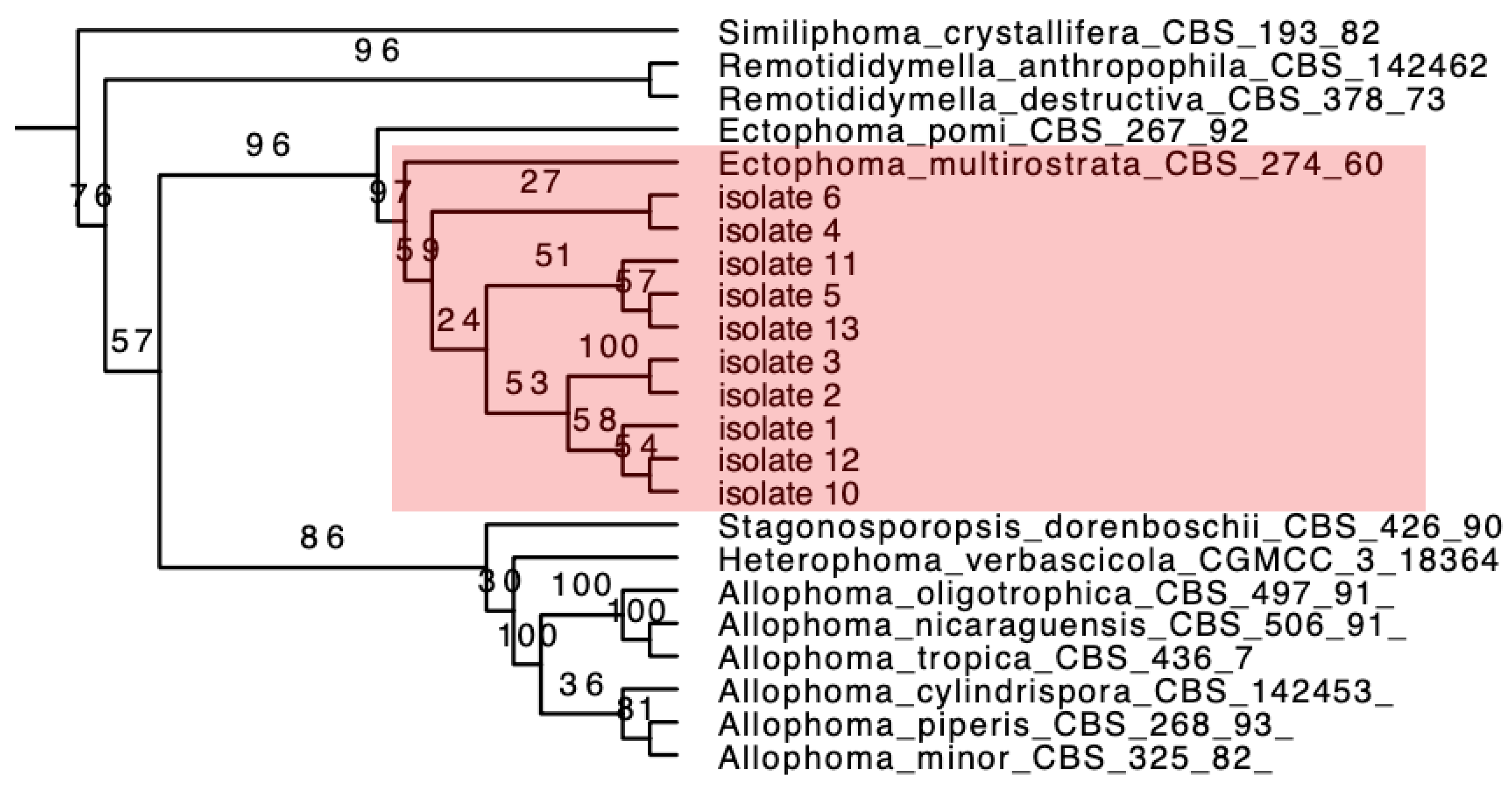
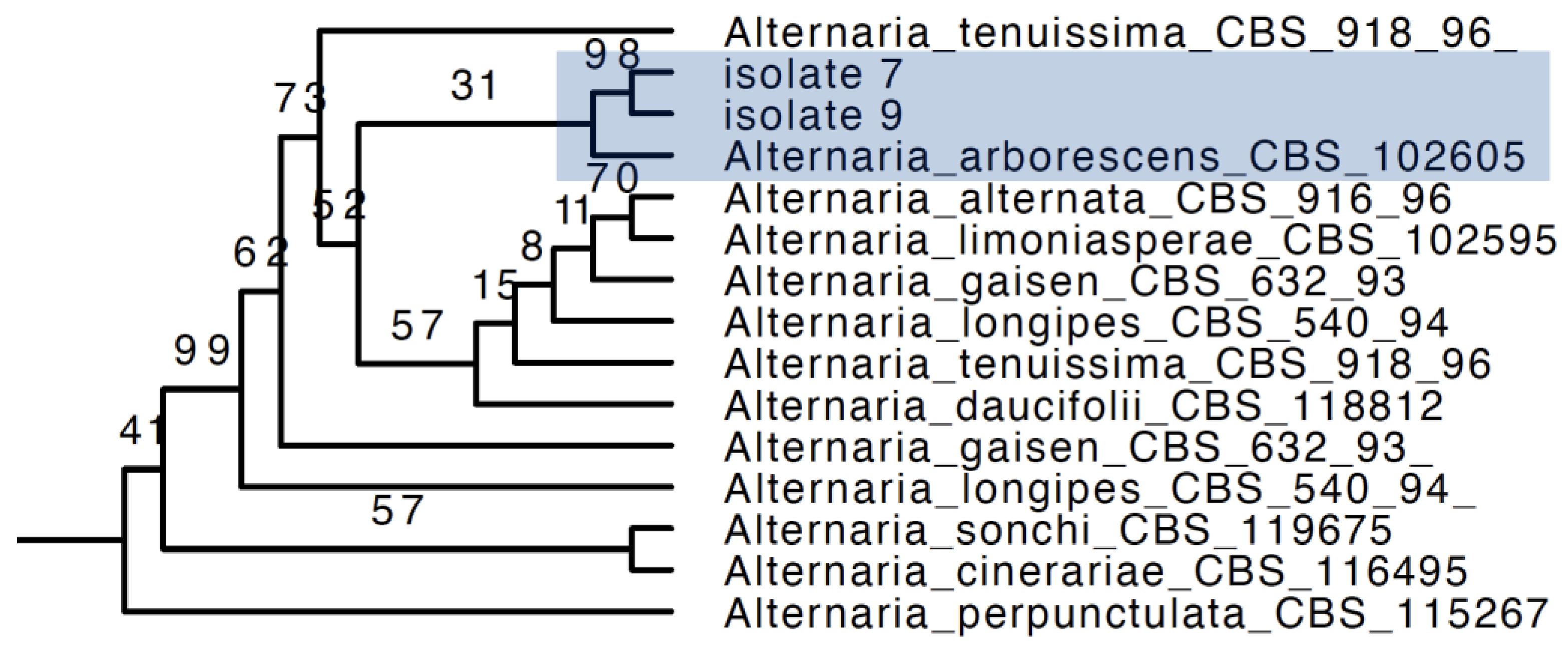
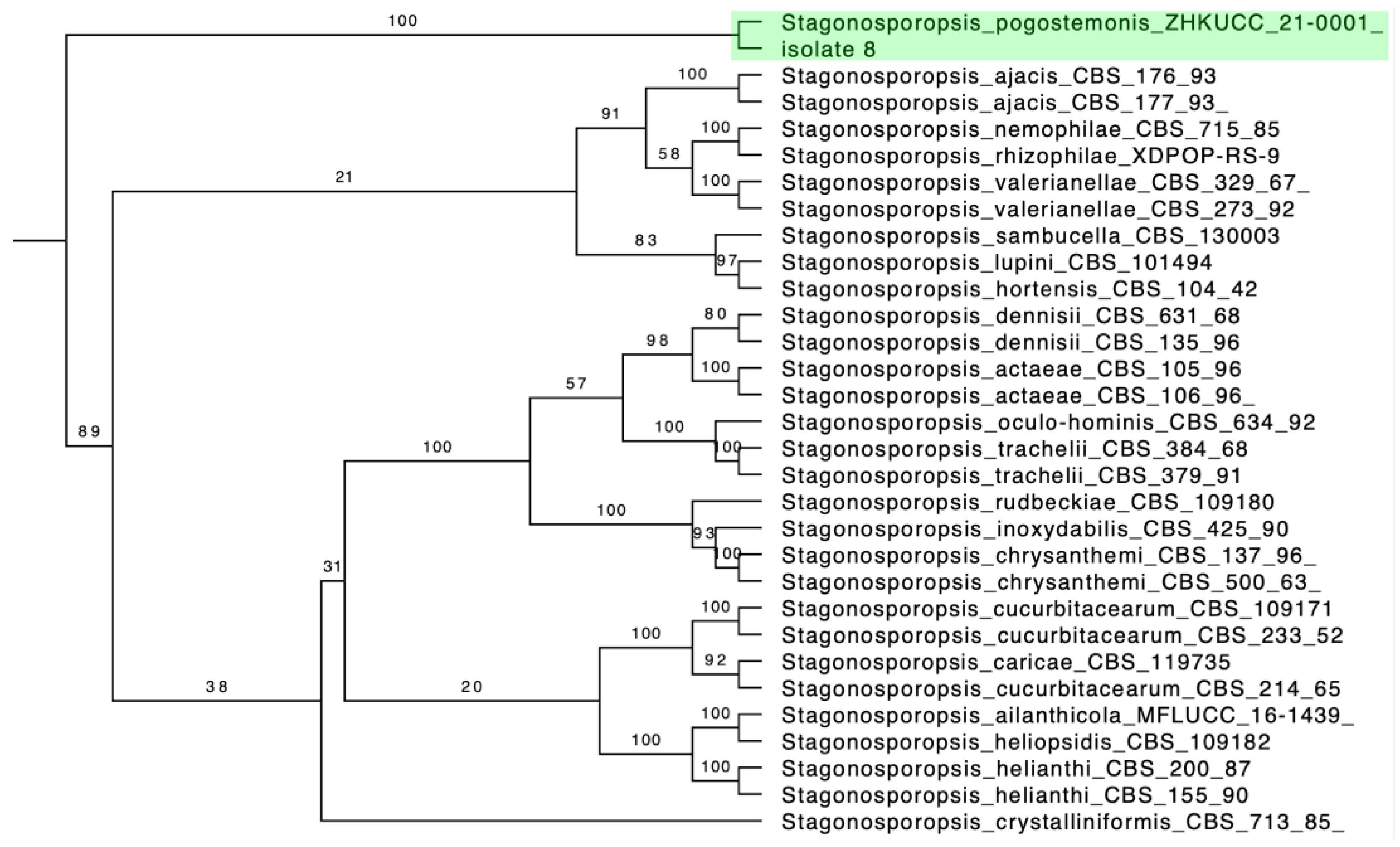
2.3.3. Phylogenetic Analysis
2.4. Fungicides Testing for the Control of the Disease Caused by E. multirostrata
2.4.1. Sensitivity Assessment In Vitro
2.4.2. Control Test In Vivo
3. Results
3.1. Field Observation of Disease
3.2. Grouping of Isolates and Phylogenetic Analysis
3.3. Pathogenicity Assessment
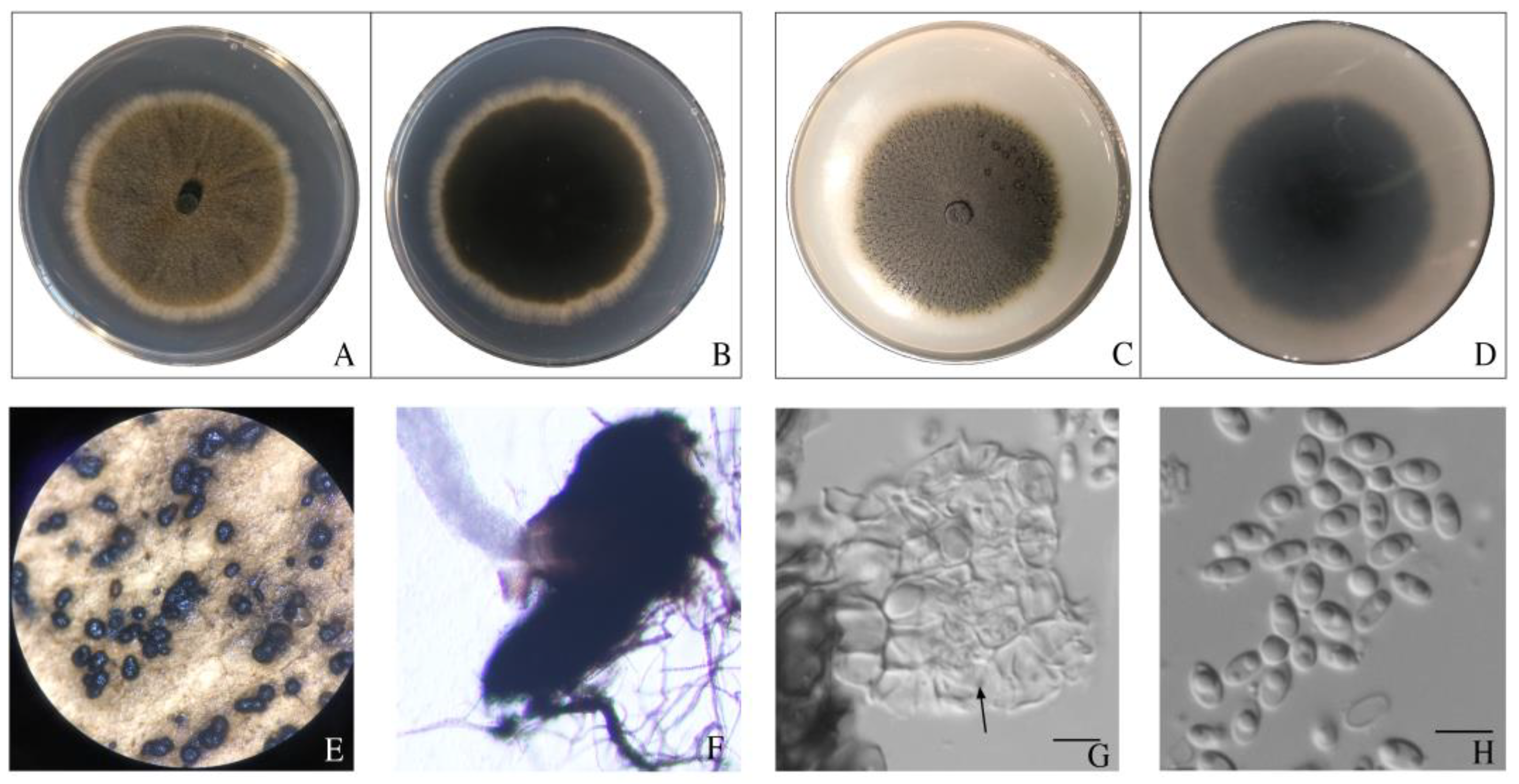
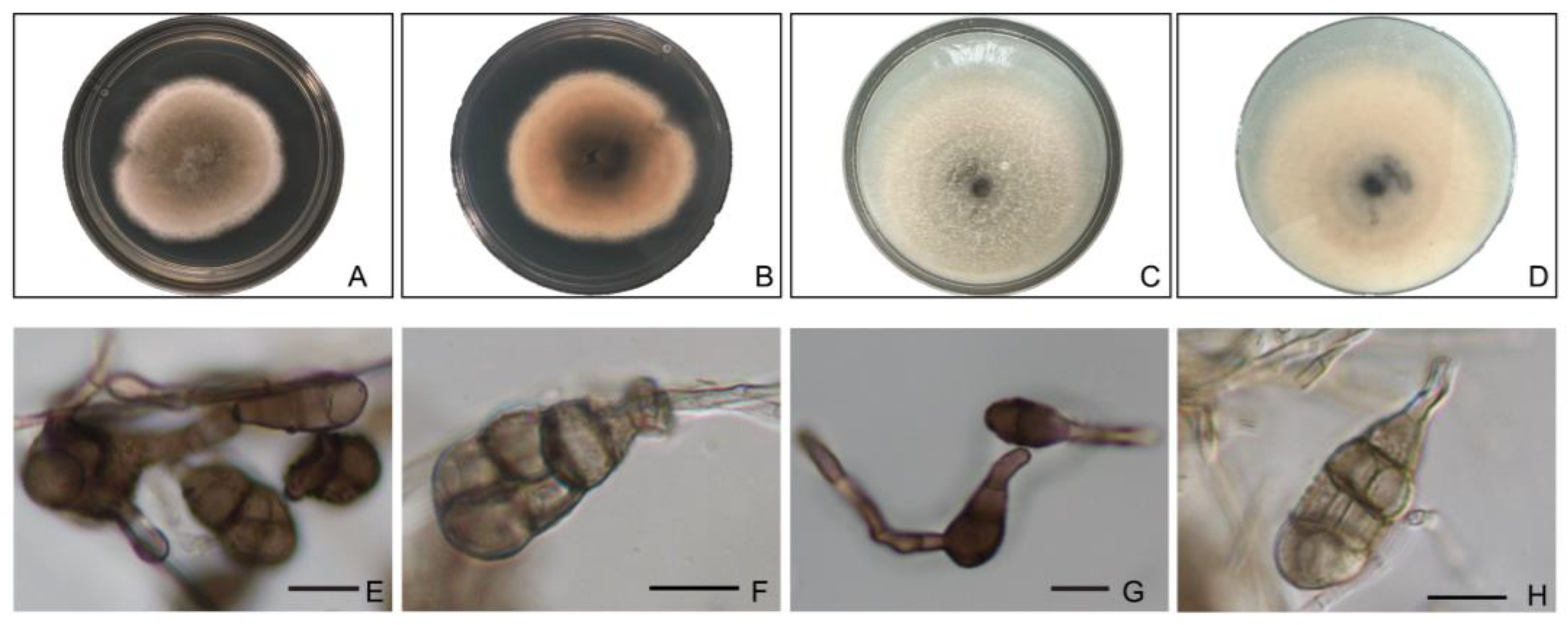
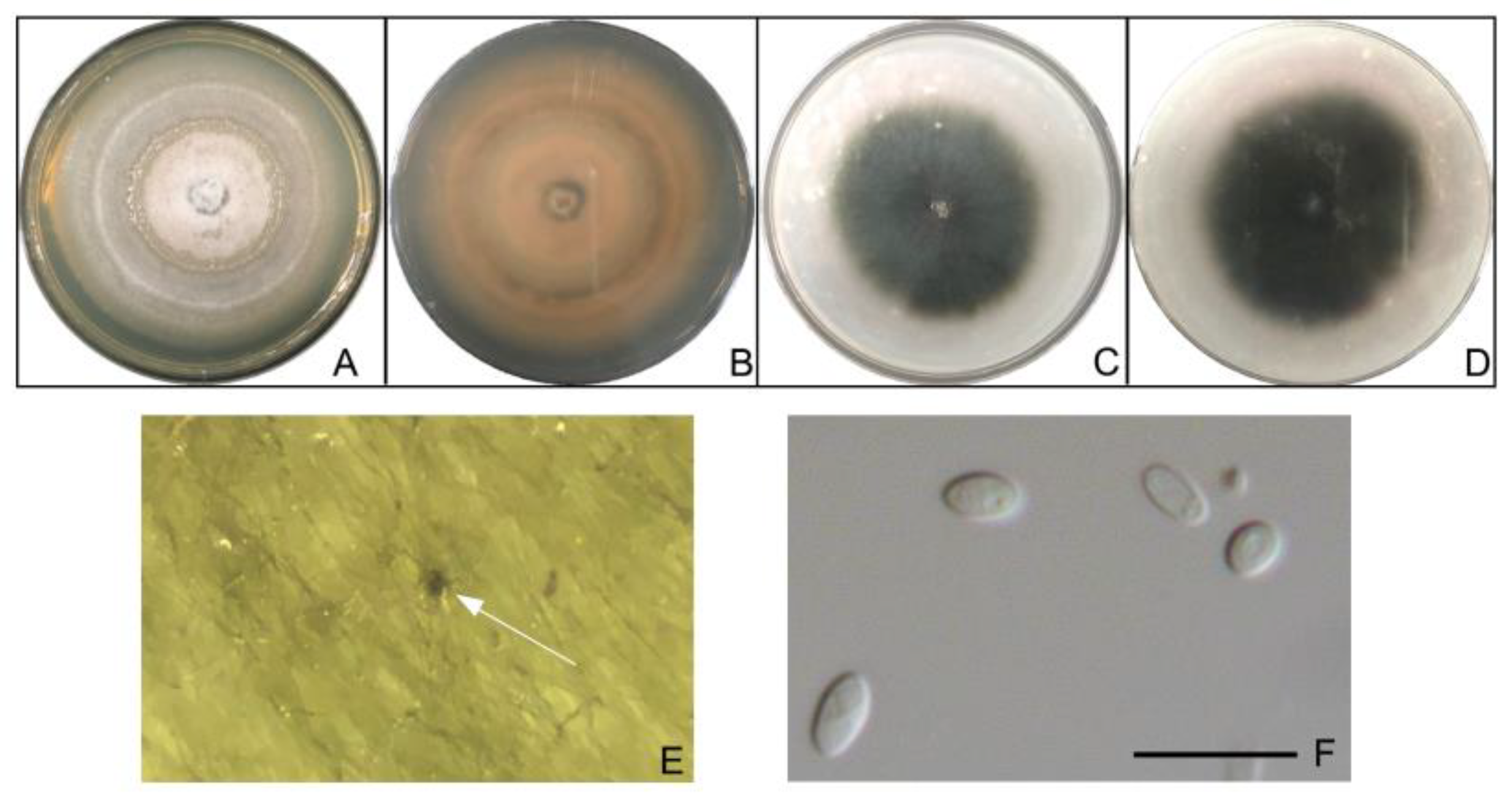
3.4. Morphological Observation of Pathogens
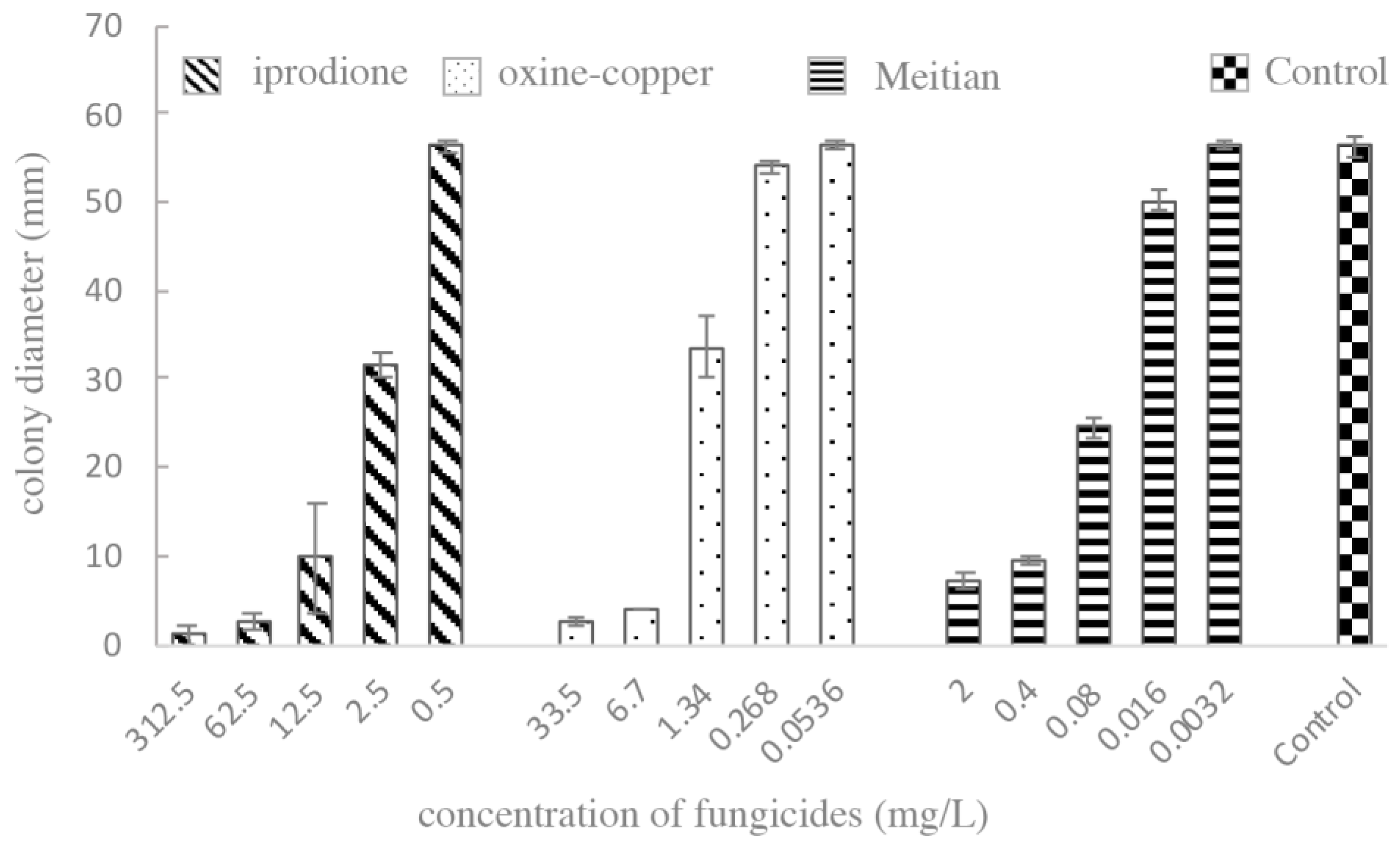
3.5. Sensitivity Assessment of Pathogens to Fungicides In Vitro
3.6. Pot Trial of Meitian against Bud Blight Disease
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Loci | Primer | Sequence |
|---|---|---|
| ITS | ITS5 | GGAAGTAAAAGTCGTAACAAGG |
| ITS4 | TCCTCCGCTTATTGATATGC | |
| LSU | LROR | ACCCGCTGAACTTAAGC |
| LR5 | TCCTGAGGGAAACTTCG | |
| tub2 | BT2A | GGTAACCAAATCGGTGCTGCTTTC |
| BT2B | ACCCTCAGTGTAGTGACCCTTGGC | |
| rpb2 | RPB2-5F | GAYGAYMGWGATCAYTTYGG |
| RPB2-7cR | CCCATRGCTTGYTTRCCCAT |
References
- Zhang, Y. Screening of Anthracnose Resistant Germplasm Resources of Dendrobium officinale and Seed Production and Application of Resistant Strains. Master’s Thesis, Zhejiang University, Hangzhou, China, 2016. (In Chinese). [Google Scholar]
- Luo, Q.; Tang, Z.; Zhang, X.; Zhong, Y.; Yao, S.; Wang, L.; Lin, C.; Luo, X. Chemical Properties and Antioxidant Activity of a Water-Soluble Polysaccharide from Dendrobium Officinale. Int. J. Biol. Macromol. 2016, 89, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Li, H.; Chen, J.; He, L.; Du, X.; Zhou, L.; Xiong, Q.; Lai, X.; Yang, Y.; Huang, S. Dendrobium officinale polysaccharides alleviate colon tumorigenesis via restoring intestinal barrier function and enhancing anti-tumor immune response. Pharmacol. Res. 2019, 148, 104417. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ding, X.; Chu, B.; Zhou, Q.; Ding, G.; Gu, S. Genetic diversity analysis and conservation of the endangered Chinese endemic herb Dendrobium Officinale Kimura et Migo (Orchidaceae) based on AFLP. Genetica 2008, 133, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zheng, X. Identification of pathogenic bacteria and cytological study on the infection process of Dendrobium officinale black spot. Acta Phytopathol. Sin. 2004, 34, 92–94. (In Chinese) [Google Scholar]
- Xiao, R.; Guan, C.-L.; Kong, D.-D.; Li, O.; Hu, X.-F. Black spot on the medicinal orchid Dendrobium Officinale caused by Cladosporium oxysporum in China. Can. J. Plant Pathol. 2018, 40, 100–104. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, J.; Liu, Y.; Dai, D. First report of black spot in Dendrobium officinale caused by A. alternata in Zhejiang Province, China. Plant Dis. 2018, 102, 824. [Google Scholar] [CrossRef]
- Zhao, L. Pathogen identification and control measures of three diseases of Dendrobium officinale. Master’s Thesis, Zhejiang University, Hangzhou, China, 2020. (In Chinese). [Google Scholar]
- Wang, L.; Fang, L.; Xie, J.; Wang, H. Preliminary study on etiology of gray mold of Dendrobium officinale. Zhejiang J. Agric. Sci. 2017, 58, 990–992. (In Chinese) [Google Scholar]
- Xie, Y.; Wang, L.; Fang, L.; Wang, H. First report of leaf spot caused by Phoma multirostrata var. microspora on Dendrobium officinale in Zhejiang Province of China. Plant Dis. 2018, 102, 1655. [Google Scholar] [CrossRef]
- Cao, P.; Fang, Y.; Zheng, Z.; Han, X.; Zou, H.; Yan, X. Occurrence of Neopestalotiopsis clavispora causing leaf spot on Dendrobium officinale in China. Plant Dis. 2022, 106, 1761. [Google Scholar] [CrossRef]
- Sun, C.; Wang, T.; Shen, X.; Wang, G.; Gao, Q.; Lou, B.; Shao, Y. First report of leaf spot caused by Cladosporium cladosporioides on Dendrobium officinale in China. Plant Dis. 2017, 101, 1055. [Google Scholar] [CrossRef]
- Liu, G.; Zhao, N.; Zhao, G. Study on Pathogenic bacteria and pathogenesis of Dendrobium officinale dieback disease. J. Southwest For. Univ. 2017, 37, 138–143. (In Chinese) [Google Scholar]
- Mirghasempour, S.A.; Michailides, T.; Chen, W.; Mao, B. Fusarium Spp. associated with Dendrobium officinale dieback disease in China. J. Fungi 2022, 8, 919. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Li, B.; Wang, R.; Liu, P.; Chen, Q. Occurrence of dieback disease caused by Fusarium equiseti on Dendrobium officinale in China. Crop Prot. 2020, 137, 105209. [Google Scholar] [CrossRef]
- Zhao, G.; Liu, G.; Zhao, N. Pathogen identification of stem rot from Dendrobium officinale of China. Jiangsu J. Agric. Sci. 2016, 32, 780–785. (In Chinese) [Google Scholar]
- Cao, P.; Zheng, Z.; Fang, Y.; Han, X.; Zou, H.; Yan, X. First report of stem rot caused by Fusarium kyushuense on Dendrobium officinale in China. Plant Dis. 2022, 106, 2257. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Lin, J.; Wang, W.; Zou, H.; Dai, Y. Isolation, identification and screening of bacteriostatic agents of Dendrobium officinale. Fujian J. Agric. Sci. 2015, 30, 901–904. (In Chinese) [Google Scholar]
- Kong, Q.; Yuan, S.; Guo, J.; Xue, C.; Li, X.; Wang, Y.; Wang, T. Identification and biological characteristics of pathogenic bacteria from Dendrobium officinale root rot. Chin. Med. 2018, 7, 1566–1570. (In Chinese) [Google Scholar]
- Zhang, Y.; Lin, B.; Zou, M.; Liang, J.; Hu, H. First report of Fusarium wilt of Dendrobium officinale caused by Fusarium oxysporum in China. Plant Dis. 2017, 101, 1039. [Google Scholar] [CrossRef]
- Shen, W.; Zhao, L.; Wang, G.; Qiu, C.; Wang, X.; Lou, B. Identification of ring rot of Dendrobium officinale and screening of fungicides. J. Plant Prot. 2018, 45, 886–892. [Google Scholar]
- Xiao, C.; Li, R.; Song, X.; Tian, X.; Zhao, Q. First report of soft rot on Dendrobium officinale caused by Epicoccum sorghinum in China. Plant Dis. 2022, 106, 1063. [Google Scholar] [CrossRef]
- Xiao, C.; Li, R. Detection and control of Fusarium oxysporum from soft rot in Dendrobium officinale by loop-mediated isothermal amplification assays. Biology 2021, 10, 1136. [Google Scholar] [CrossRef] [PubMed]
- Saberi Riseh, R.; Skorik, Y.A.; Thakur, V.K.; Moradi Pour, M.; Tamanadar, E.; Noghabi, S.S. Encapsulation of plant biocontrol bacteria with alginate as a main polymer material. Int. J. Mol. Sci. 2021, 22, 11165. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, N.; Fujimura, M.; Oshima, M.; Motoyama, T.; Ichiishi, A.; Yamada-Okabe, H.; Yamaguchi, I. Effects of iprodione and fludioxonil on glycerol synthesis and hyphal development in candida albicans. Biosci. Biotechnol. Biochem. 2002, 66, 2209–2215. [Google Scholar] [CrossRef] [PubMed]
- Nicoletti, G.; Domalewska, E.; Borland, R. Fungitoxicity of oxine and copper oxinate: Activity spectrum, development of resistance and synergy. Mycol. Res. 1999, 103, 1073–1084. [Google Scholar] [CrossRef]
- Avenot, H.F.; Michailides, T.J. Progress in understanding molecular mechanisms and evolution of resistance to succinate dehydrogenase inhibiting (SDHI) fungicides in phytopathogenic fungi. Crop Prot. 2010, 29, 643–651. [Google Scholar] [CrossRef]
- Li, Y.; Ma, X.; Lu, G. Systematic investigation of the toxic mechanism of difenoconazole on protein by spectroscopic and molecular modeling. Pestic. Biochem. Physiol. 2013, 105, 155–160. [Google Scholar] [CrossRef]
- Crous, P.W.; Lombard, L.; Sandoval-Denis, M.; Seifert, K.; Schroers, H.-J.; Chaverri, P.; Gené, J.; Guarro, J.; Hirooka, Y.; Bensch, K. Fusarium: More than a node or a foot-shaped basal cell. Stud. Mycol. 2021, 98, 100116. [Google Scholar] [CrossRef]
- Massicotte, H.; Melville, L.; Peterson, R. Structural characteristics of root fungal interactions for five ericaceous species in Eastern Canada. Can. J. Bot. 2005, 83, 1057–1064. [Google Scholar] [CrossRef]
- Guo, L.; Hyde, K.; Liew, E. A method to promote sporulation in palm endophytic fungi. Fungal Divers. 1998, 1, 109–113. [Google Scholar]
- Pan, L.; Cui, C.; Wang, B. A rapid DNA extraction method for filamentous fungi for PCR amplification. J. Microbiol. 2010, 3, 450–453. (In Chinese) [Google Scholar]
- Valenzuela-Lopez, N.; Cano-Lira, J.; Guarro, J.; Sutton, D.A.; Wiederhold, N.; Crous, P.; Stchigel, A. Coelomycetous Dothideomycetes with emphasis on the families Cucurbitariaceae and Didymellaceae. Stud. Mycol. 2018, 90, 1–69. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- De Hoog, G.; Van den Ended, A. Molecular diagnostics of clinical strains of filamentous basidiomycetes. Mycoses 1998, 41, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Rehner, S.A.; Samuels, G.J. Taxonomy and phylogeny of gliocladium analysed from nuclear large subunit ribosomal DNA sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Sung, G.-H.; Sung, J.-M.; Hywel-Jones, N.L.; Spatafora, J.W. A multi-gene phylogeny of Clavicipitaceae (Ascomycota, fungi): Identification of localized incongruence using a combinational bootstrap approach. Mol. Phylogenet. Evol. 2007, 44, 1204–1223. [Google Scholar] [CrossRef]
- Chen, Q.; Jiang, J.; Zhang, G.; Cai, L.; Crous, P.W. Resolving the Phoma enigma. Stud. Mycol. 2015, 82, 137–217. [Google Scholar] [CrossRef]
- Hou, L.; Groenewald, J.; Pfenning, L.; Yarden, O.; Crous, P.W.; Cai, L. The Phoma-like dilemma. Stud. Mycol. 2020, 96, 309–396. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Wang, Q.-H.; Fan, K.; Li, D.-W.; Han, C.-M.; Qu, Y.-Y.; Qi, Y.-K.; Wu, X.-Q. Identification, virulence and fungicide sensitivity of Colletotrichum gloeosporioides ss responsible for walnut anthracnose disease in China. Plant Dis. 2020, 104, 1358–1368. [Google Scholar] [CrossRef]
- Qi, J. Identification of Pathogenic Bacteria of Dendrobium Officinale Stem Rot and Study on Prevention and Growth of Bacillus velezensis D-12. Master’s Thesis, Zhejiang University, Hangzhou, China, 2021. (In Chinese). [Google Scholar]
- Borman, A.M.; Johnson, E.M. Sequence-based identification and classification of fungi. Trends Syst. Bact. Fungi 2021, 108, 198–216. [Google Scholar] [CrossRef]
- Glaeser, S.P.; Kämpfer, P. Multilocus sequence analysis (MLSA) in prokaryotic taxonomy. Syst. Appl. Microbiol. 2015, 38, 237–245. [Google Scholar] [CrossRef]
- Kashyap, A.S.; Manzar, N.; Ahamad, F.; Tilgam, J.; Sharma, P.K.; Saxena, A.K. First report of root rot disease in green gram (Vigna radiata) caused by Ectophoma multirostrata in India. Plant Dis. 2022, 106, 2256. [Google Scholar] [CrossRef]
- Harteveld, D.; Akinsanmi, O.; Drenth, A. Multiple Alternaria species groups are associated with leaf blotch and fruit spot diseases of apple in Australia. Plant Pathol. 2013, 62, 289–297. [Google Scholar] [CrossRef]
- Ali, S.; Abbasi, P.; Rehman, S.; Ellouze, W. First report of moldy core of sweet tango apples from New Zealand caused by Alternaria arborescens. Plant Dis. 2021, 105, 2719. [Google Scholar] [CrossRef]
- Serdani, M.; Kang, J.-C.; Andersen, B.; Crous, P.W. Characterisation of Alternaria species-groups associated with core rot of apples in South Africa. Mycol. Res. 2002, 106, 561–569. [Google Scholar] [CrossRef]
- Ntasiou, P.; Myresiotis, C.; Konstantinou, S.; Papadopoulou-Mourkidou, E.; Karaoglanidis, G.S. Identification, Characterization and mycotoxigenic ability of Alternaria spp. causing core rot of apple fruit in Greece. Int. J. Food Microbiol. 2015, 197, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Matić, S.; Tabone, G.; Garibaldi, A.; Gullino, M.L. Alternaria leaf spot caused by Alternaria species: An emerging problem on ornamental plants in Italy. Plant Dis. 2020, 104, 2275–2287. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, N.; Bashir, U.; Mushtaq, S. First report of leaf spot of rice caused by Alternaria arborescens in Pakistan. Plant Dis. 2014, 98, 846. [Google Scholar] [CrossRef] [PubMed]
- Akram, W.; Li, G.; Ahmad, A.; Anjum, T.; Ali, B.; Luo, W.; Guo, J.; Xie, D.; Wang, Q. Leaf spot disease caused by Alternaria arborescens, A. tenuissima, and A. infectoria on Brassica rapa subsp. parachinensis in China. Plant Dis. 2019, 103, 2480. [Google Scholar] [CrossRef]
- Garibaldi, A.; Bertetti, D.; Matić, S.; Luongo, I.; Gullino, M. First report of leaf spots caused by Alternaria arborescens on symphyotrichum novi-belgii in Italy. Plant Dis. 2020, 104, 281. [Google Scholar] [CrossRef]
- Razak, N.J.; Abass, M.H. First report of Alternaria arborescens causing early blight on tomato in Iraq: Short notes. Basrah J. Agric. Sci. 2021, 34, 230–232. [Google Scholar] [CrossRef]
- Aloi, F.; Riolo, M.; Sanzani, S.M.; Mincuzzi, A.; Ippolito, A.; Siciliano, I.; Pane, A.; Gullino, M.L.; Cacciola, S.O. Characterization of Alternaria species associated with heart rot of pomegranate fruit. J. Fungi 2021, 7, 172. [Google Scholar] [CrossRef]
- Luo, Y.; Hou, L.; Förster, H.; Pryor, B.; Adaskaveg, J. Identification of Alternaria species causing heart rot of pomegranates in California. Plant Dis. 2017, 101, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Luo, J.; Li, B.; Song, Y.; Mu, W.; Liu, F. Bioactivity, physiological characteristics and efficacy of the SDHI fungicide pydiflumetofen against Sclerotinia sclerotiorum. Pestic. Biochem. Physiol. 2019, 160, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Hu, H.; Lai, R.; Wu, R. Field control effect of 18% flumazoylhydroxylamine phenoxymethyclozole suspension on citrus scab. Pesticide 2018, 57, 690–692.35. (In Chinese) [Google Scholar]
- Zhou, Y.; Han, X.; Guo, Z.; Wang, S.; Li, J.; Zhang, S. Evaluation of field control effect of 200g/L fluzoylhydroxylamine and phenoxymethyclozole suspension on cucumber powdery mildew. J. Chang. Veg. 2021, 10, 73–75. (In Chinese) [Google Scholar]











| Species | Strain Number | GenBank Accession Numbers | |||
|---|---|---|---|---|---|
| ITS | LSU | tub2 | rpb2 | ||
| Ectophoma sp. | 1 | OQ073676 | OQ096504 | OQ271767 | OQ271782 |
| Ectophoma sp. | 2 | OQ073677 | OQ096505 | OQ271768 | OQ271783 |
| Ectophoma sp. | 3 | OQ073678 | OQ096506 | OQ271769 | OQ271784 |
| Ectophoma sp. | 4 | OQ073679 | OQ096507 | OQ271770 | OQ271785 |
| Ectophoma sp. | 5 | OQ073680 | OQ096508 | OQ271771 | OQ271786 |
| Ectophoma sp. | 6 | OQ073681 | OQ096509 | OQ271772 | OQ271787 |
| Alternaria sp. | 7 | OQ073682 | OQ096510 | OQ271773 | OQ271788 |
| Stagonosporopsis sp. | 8 | OQ073683 | OQ096511 | OQ271774 | OQ271789 |
| Alternaria sp. | 9 | OQ073684 | OQ096512 | OQ271775 | OQ271790 |
| Ectophoma sp. | 10 | OQ073685 | OQ096513 | OQ271776 | OQ271791 |
| Ectophoma sp. | 11 | OQ073686 | OQ096514 | OQ271777 | OQ271792 |
| Ectophoma sp. | 12 | OQ073687 | OQ096515 | OQ271778 | OQ271793 |
| Ectophoma sp. | 13 | OQ073688 | OQ096516 | OQ271779 | OQ271794 |
| Geographic Origin | Species | Number of Isolates | Isolate Frequency (%) |
|---|---|---|---|
| Yueqing City (Zhejiang Province) | Ectophoma spp. | 91 | 71.6 |
| Alternaria spp. | 27 | 21.3 | |
| Stagonosporopsis spp. | 9 | 7.1 | |
| Total | 127 | 100 |
| E. multirostrata | Iprodione (mg/L) | Oxine-Copper (mg/L) | Meitian (mg/L) |
|---|---|---|---|
| EC50 | 2.10 | 1.78 | 0.09 |
| 95% CI | 1.36–3.05 | 1.43–2.24 | 0.07–0.13 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Sha, H.; Chen, W.; Mao, B. Characterization and Control of Dendrobium officinale Bud Blight Disease. Pathogens 2023, 12, 621. https://doi.org/10.3390/pathogens12040621
Zhang J, Sha H, Chen W, Mao B. Characterization and Control of Dendrobium officinale Bud Blight Disease. Pathogens. 2023; 12(4):621. https://doi.org/10.3390/pathogens12040621
Chicago/Turabian StyleZhang, Jinzhao, Haodong Sha, Weiliang Chen, and Bizeng Mao. 2023. "Characterization and Control of Dendrobium officinale Bud Blight Disease" Pathogens 12, no. 4: 621. https://doi.org/10.3390/pathogens12040621
APA StyleZhang, J., Sha, H., Chen, W., & Mao, B. (2023). Characterization and Control of Dendrobium officinale Bud Blight Disease. Pathogens, 12(4), 621. https://doi.org/10.3390/pathogens12040621