

Survey, Detection, Characterization of Papaya Ringspot Virus from Southern India and Management of Papaya Ringspot Disease

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Survey and Collection of Virus Isolates
2.2. Detection and Sequencing of PRSV
2.3. PRSV Complete Genome Comparison, Phylogenetic, and Recombination Analysis
2.4. Integrated Management of PRSV Disease under Field Conditions
3. Results
3.1. Survey, Incidence, and Symptomatology of PRSD
3.2. Molecular Characterization of PRSV
3.3. Comparison of the Complete Genome of PRSV-BGK Isolate with Other PRSV Sequences
3.4. Phylogenetic Analysis of PRSV
3.5. Recombination Analysis of PRSV
3.6. Integrated Management of PRSD under Field Conditions
3.6.1. Effect of Insecticides and Biorationals on PRSD Incidence, Growth, and Yield
Parameters of Papaya
Disease Incidence
Growth, Yield, and Yield Parameters
3.6.2. Integrated Disease Management Modules for the Management of PRSD
Disease Incidence
Growth, Yield, and Yield Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carvalho, F.A.; Renner, S.S. Molecular phylogenetics and evolution a dated phylogeny of the papaya family (Caricaceae) reveals the crop’s closest relatives and the family’s biogeographic history. Mol. Phylogenet. Evol. 2012, 65, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Niklas, K.J.; Marler, T.E. Carica papaya (Caricaceae): A case study into the effects of domestication on plant vegetative growth and reproduction. Am. J. Bot. 2007, 94, 999–1002. [Google Scholar] [CrossRef]
- Evans, E.A.; Ballen, F.H. An Overview of Global Papaya Production, Trade, and Consumption: FE913/FE913, 9/2012; EDIS 2012; University of Florida: Gainesville, FL, USA, 2012. [Google Scholar]
- Huerta-Ocampo, J.Á.; Osuna-Castro, J.A.; Lino-López, G.J.; Barrera-Pacheco, A.; Mendoza-Hernández, G.; De León-Rodríguez, A.; de la Rosa, A.P.B. Proteomic analysis of differentially accumulated proteins during ripening and in response to 1-MCP in papaya fruit. J. Proteom. 2012, 75, 2160–2169. [Google Scholar] [CrossRef]
- Anonymous. FAOSTAT Data Base. 2020. Available online: http://nhb.gov.in/database (accessed on 1 September 2022).
- Akhter, M.S.; Akanda, A.M. Effect of papaya ringspot virus on growth and yield of papaya. Bangladesh J. Crop. Sci. 2008, 19, 203–208. [Google Scholar]
- Mishra, R.; Patil, S.; Patil, A.; Patil, B.L. Sequence diversity studies of papaya ringspot virus isolates in South India reveal higher variability and recombination in the 5′-terminal gene sequences. Virus Dis. 2019, 30, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Premchand, U.; Mesta, R.K.; Basavarajappa, M.P.; Venkataravanappa, V.; Devappa, V.; Narasimha Reddy, L.R.C.; Shankarappa, K.S. Identification of novel begomoviruses associated with leaf curl disease of papaya (Carica papaya L.) in India. Agronomy 2023, 13, 3. [Google Scholar] [CrossRef]
- Alcala-Briseno, R.I.; Casarrubias-Castillo, K.; Lopez-Ley, D.; Garrett, K.A.; Silva-Rosales, L. Network analysis of the papaya orchard virome from two agroecological regions of Chiapas, Mexico. Msystems 2020, 5, e00423-19. [Google Scholar] [CrossRef] [Green Version]
- Alabi, O.J.; Al Rwahnih, M.; Brown, J.K.; Idris, A.M.; Gregg, L.; Kmieciak, E.; Setamou, M.; Jifon, J.L. First report of papaya (Carica papaya) naturally infected with the introduced tomato yellow leaf curl virus-Israel. Plant Dis. 2016, 100, 1959. [Google Scholar] [CrossRef]
- Jain, R.K.; Nasiruddin, K.M.; Sharma, J.; Pant, R.P.; Varma, A. First report of occurrence of papaya ringspot virus infecting papaya in Bangladesh. Plant Dis. 2004, 88, 221. [Google Scholar] [CrossRef]
- Kunkalikar, S.R.; Poojari, S.; Arun, B.M.; Rajagopalan, P.A.; Chen, T.-C.; Yeh, S.-D.; Naidu, R.A.; Zehr, U.B.; Ravi, K.S. Importance and genetic diversity of vegetable-infecting tospoviruses in India. Phytopathology 2011, 101, 367–376. [Google Scholar] [CrossRef] [Green Version]
- Lokhande, N.M.; Moghe, P.G.; Matte, A.D.; Hiware, B.J. Occurrence of papaya ringspot virus (PRSV) in Vidharbha regions of Maharashtra. J. Soils Crop. 1992, 2, 36–39. [Google Scholar]
- Hussain, S.; Varma, A. Occurrence of papaya ringspot virus from Amritsar (Punjab) India. J. Phytopathol. Res. 1994, 7, 77–78. [Google Scholar]
- Purcifull, D.E. Papaya ringspot virus. CMI/AAB descriptions of plant viruses. Assoc. Appl. Biol. 1984, 7, 292. [Google Scholar] [CrossRef]
- Gonsalves, D.; Vegas, A.; Prasartsee, V.; Drew, R.A.; Suzuki, J.Y.; Tripathi, S. Developing papaya to control papaya ringspot virus by transgenic resistance, intergeneric hybridization, and tolerance breeding. Plant Breed. Rev. 2010, 26, 35–73. [Google Scholar]
- Tripathi, S.; Suzuki, J.Y.; Ferreira, S.A.; Gonsalves, D. Papaya ringspot virus-P: Characteristics, pathogenicity, sequence variability and control. Mol. Plant Pathol. 2008, 9, 269–280. [Google Scholar] [CrossRef]
- Roy, G.; Jain, R.K.; Bhat, A.I.; Varma, A. Comparative host range and serological studies of papaya ringspot potyvirus isolates. Indian Phytopathol. 1999, 52, 14–17. [Google Scholar]
- Webb, R.E.; Scott, H.A. Isolation and identification of watermelon mosaic viruses 1 and 2. Phytopathology 1965, 55, 895–900. [Google Scholar]
- Yeh, S.-D.; Jan, F.-J.; Chiang, C.-H.; Doong, T.-J.; Chen, M.-C.; Chung, P.-H.; Bau, H.-J. Complete nucleotide sequence and genetic organization of papaya ringspot virus RNA. J. Gen. Virol. 1992, 73, 2531–2541. [Google Scholar] [CrossRef]
- Huang, C.-H.; Tai, C.-H.; Lin, R.-S.; Chang, C.-J.; Jan, F.-J. Biological, pathological, and molecular characteristics of a new potyvirus, dendrobium chlorotic mosaic virus, infecting Dendrobium orchid. Plant Dis. 2019, 103, 1605–1612. [Google Scholar] [CrossRef]
- Parris, G.K. A New Disease of Papaya in Hawaii. Proc. Am. Soc. Hortic. Sci. 1938, 36, 263–265. [Google Scholar]
- Lindner, R.C.; Jensen, D.D.; Ikeda, W. Ringspot: New papaya plunderer. Hawaii Farm. Home 1945, 8, 10–12. [Google Scholar]
- Jensen, D.D. Papaya virus diseases with special reference to papaya ringspot. Phytopathology 1949, 39, 191–211. [Google Scholar]
- Capoor, S.P.; Varma, P.M. A mosaic disease of Carica papaya L. in the Bombay province. Curr. Sci. 1948, 17, 265–266. [Google Scholar]
- Byadgi, A.S.; Anahosur, K.H.; Kulakarni, M.S. Ringspot virus in papaya. Hindu 1995, 118, 28. [Google Scholar]
- Van Loon, L.C.; Bakker, P.; Pieterse, C.M.J. Systemic resistance induced by rhizosphere bacteria. Annu. Rev. Phytopathol. 1998, 36, 453–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Ani, R.A.; Adhab, M.A. Bean yellow mosaic virus (BYMV) on broadbean:Characterization and resistance induced by Rhizobium leguminosarum. J. Pure Appl. Microbiol. 2013, 7, 135–142. [Google Scholar]
- Al-Ani, R.A.; Athab, M.A.; Matny, O.N. Management of potato virus Y (PVY) in potato by some biocontrol agents under field conditions. Adv. Environ. Biol. 2013, 7, 441–444. [Google Scholar]
- Acosta-Leal, R.; Duffy, S.; Xiong, Z.; Hammond, R.W.; Elena, S.F. Advances in plant virus evolution: Translating evolutionary insights into better disease management. Phytopathology 2011, 101, 1136–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elena, S.F.; Fraile, A.; García-Arenal, F. Evolution and Emergence of Plant Viruses. Adv. Virus Res. 2014, 88, 161–191. [Google Scholar]
- Jia, R.; Zhao, H.; Huang, J.; Kong, H.; Zhang, Y.; Guo, J.; Huang, Q.; Guo, Y.; Wei, Q.; Zuo, J.; et al. Use of RNAi technology to develop a PRSV-resistant transgenic papaya. Sci. Rep. 2017, 7, 12636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendoza, E.M.; Laurena, A.C.; Botella, J.R. Recent advances in the development of transgenic papaya technology. Biotechnol. Annu. Rev. 2008, 14, 423–462. [Google Scholar]
- Chen, G.; Ye, C.M.; Huang, J.C.; Yu, M.; Li, B.J. Cloning of the papaya ringspot virus (PRSV) replicase gene and generation of PRSV-resistant papayas through the introduction of the PRSV replicase gene. Plant Cell. Rep. 2001, 20, 272–277. [Google Scholar] [CrossRef]
- Fang, J.; Lin, A.; Qiu, W.; Cai, H.; Umar, M.; Chen, R.; Ming, R. Transcriptome profiling revealed stress-induced and disease resistance genes up-regulated in PRSV resistant transgenic papaya. Front. Plant Sci. 2016, 7, 855. [Google Scholar] [CrossRef] [Green Version]
- Kung, Y.-J.; You, B.-J.; Raja, J.A.J.; Chen, K.-C.; Huang, C.-H.; Bau, H.-J.; Yang, C.-F.; Huang, C.-H.; Chang, C.-P.; Yeh, S.-D. Nucleotide sequence-homology-independent breakdown of transgenic resistance by more virulent virus strains and a potential solution. Sci. Rep. 2015, 5, 9804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitra, S.K.; Dinesh, M.R. Papaya breeding in India- achievements and future thrust. Acta Hortic. 2019, 1250, 23–28. [Google Scholar] [CrossRef]
- Younas, M.; Zou, H.; Laraib, T.; Rajpoot, N.A.; Khan, N.A.; Zaidi, A.A.; Ayaz Kachelo, G.; Akhtar, M.W.; Hayat, S.; Al-Sadi, A.M. The Impact of insecticides and plant extracts on the suppression of insect vector (Bemisia Tabaci) of mungbean yellow mosaic virus (MYMV). PLoS ONE 2021, 16, e0256449. [Google Scholar] [CrossRef]
- El-Bilawy, E.H.; Al-Mansori, A.-N.A.; Alotibi, F.O.; Al-Askar, A.A.; Arishi, A.A.; Teiba, I.I.; Sabry, A.E.-N.; Elsharkawy, M.M.; Heflish, A.A.; Behiry, S.I.; et al. Antiviral and antifungal of Ulva fasciata extract: HPLC analysis of polyphenolic compounds. Sustainability 2022, 14, 12799. [Google Scholar] [CrossRef]
- El-Bilawy, E.H.; Al-Mansori, A.-N.A.; Soliman, S.A.; Alotibi, F.O.; Al-Askar, A.A.; Arishi, A.A.; Sabry, A.E.-N.; Elsharkawy, M.M.; Heflish, A.A.; Behiry, S.I.; et al. Antifungal, antiviral, and HPLC analysis of phenolic and flavonoid compounds of Amphiroa Anceps extract. Sustainability 2022, 14, 12253. [Google Scholar] [CrossRef]
- Elsharkawy, M.M.; El-Okkiah, S.; Elsadany, A.Y.; Bedier, M.Y.; Omara, R.I.; Behiry, S.I.; Hassan, S.; Abdelkhalek, A. Systemic resistance induction of tomato plants against tomato mosaic virus by microalgae. Egypt. J. Biol. Pest. Control. 2022, 32, 37. [Google Scholar] [CrossRef]
- Hemmati, F.; Behjatnia, S.A.-A.; Moghadam, A.; Afsharifar, A. Induction of systemic resistance against cucumber mosaic virus (CMV) and tomato yellow leaf curl virus (TYLCV) in tomato. Int. J. Pest. Manag. 2023, 69, 1–14. [Google Scholar] [CrossRef]
- Vathshalyan, N.; Hariharan, G.; Karunarathna, B. Potentials of seaweed extracts in managing phytopathogens. In Environment Sustenance and Food Safety: Need for More Vibrant Policy Initiatives for Sri Lanka; National Science and Technology Commission: Battaramulla, Sri Lanka, 2022; pp. 151–173. [Google Scholar]
- Jaastad, G. Late Dormant Rapeseed Oil Treatment against black cherry aphid and cherry fruit moth in sweet cherries. J. Appl. Entomol. 2007, 131, 284–288. [Google Scholar] [CrossRef]
- Jiménez, E.; Dorta, F.; Medina, C.; Ramírez, A.; Ramírez, I.; Peña-Cortés, H. Anti-phytopathogenic activities of macro-algae extracts. marine drugs. Mar. Drugs 2011, 9, 739–756. [Google Scholar] [CrossRef] [Green Version]
- Yasmin, M.S.; Bachchu, M.A.A.; Hossain, M.A. Toxic and repellent effects of three botanical oils against adult Aphis Craccivora Koch (Homoptera: Aphididae) under laboratory conditions. Univ. J. Zool. Rajshahi. Univ. 2017, 36, 39–48. [Google Scholar]
- Pushpa, R.N.; Shantamma; Anil, P.; Manjunath, B.; Bhose, S.; Sawan, K.; Rangaswamy, K.T.; Girish, T.R.; Nagaraju, N. Molecular characterization, epidemiology and management of the papaya ringspot virus (PRSV) in papaya under southern Indian conditions. Int. J. Agric. Sci. 2018, 10, 5029–5038. [Google Scholar]
- Singh, S.K.; Jha, P.K.; Ray, P.K. Integrated management of papaya ringspot virus (PRSV) in agro ecological conditions of Bihar. In Proceedings of the II International Symposium on Papaya, Madurai, India, 9–12 December 2008; Volume 851, pp. 487–494. [Google Scholar]
- Kalleshwaraswamy, C.M.; Krishna Kumar, N.K.; Dinesh, M.R.; Chandrashekhar, K.N.; Manjunatha, M. Evaluation of insecticides and oils on aphid vectors for the management of papaya ringspot virus (PRSV). Karnataka J. Agri. Sci. 2010, 22, 552–553. [Google Scholar]
- Hansen, L.M.; Nielsen, S.L. Efficacy of mineral oil combined with insecticides for the control of aphid virus vectors to reduce potato virus Y infections in seed potatoes (Solanum tuberosum). Acta Agric. Scand. B Soil Plant Sci. 2012, 62, 132–137. [Google Scholar] [CrossRef]
- Kavyashri, V.V. Induced Defense against cucumber mosaic virus (CMV) Disease in Chilli (Capsicum annuum L.); University of Agricultural Sciences, GKVK: Bangalore, India, 2019. [Google Scholar]
- Patel, D.; Ahlawat, T.R.; Jena, S.; Chaudhary, A. Effect of silicon and seaweed extract on physical and sensory quality of papaya cv. red lady. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 504–510. [Google Scholar] [CrossRef]
- Sugandhika, M.G.G.; Pakeerathan, K.; Fernando, W.M.K. Efficacy of seaweed extract on chilli leaf curl virus. J. Agro-Technol. Rural. Sci. 2021, 1, 1. [Google Scholar] [CrossRef]
- Bateson, M.F.; Henderson, J.; Chaleeprom, W.; Gibbs, A.J.; Dale, J.L. Papaya ringspot potyvirus: Isolate variability and the origin of PRSV Type P (Australia). J. Gen. Virol. 1994, 75, 3547–3553. [Google Scholar] [CrossRef] [PubMed]
- Lal, A.; Kil, E.; Thuy, V.T.B.; Fadhila, C.; Ho, P.T.; Byun, H.; Dao, H.T.; Kim, J.; Lee, S. Milk vetch dwarf virus infection in the Solanaceae and Caricaceae families in Southeast Asia. Plant Pathol. 2020, 69, 1026–1033. [Google Scholar] [CrossRef]
- Tuo, D.; Shen, W.; Yang, Y.; Yan, P.; Li, X.; Zhou, P. Development and Validation of a Multiplex Reverse Transcription PCR assay for simultaneous detection of three papaya viruses. Viruses 2014, 6, 3893–3906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villamor, D.E.V.; Ardales, E.Y. Cloning and sequence analysis of the coat protein gene of zucchini yellow mosaic potyvirus. Former. Philipp. Phytopathol. 2003, 39, 16–35. [Google Scholar]
- Ortiz-Rojas, L.Y.; Chaves-Bedoya, G. Molecular characterization of two papaya ringspot virus isolates that cause devastating symptoms in Norte de Santander, Colombia. Eur. J. Plant Pathol. 2017, 148, 883–894. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989; ISBN 0-87969-309-6. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. In Nucleic Acids Symposium Series; c1979-c2000; Information Retrieval Ltd.: London, UK, 1999; Volume 41, pp. 95–98. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X Version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Martin, D.P.; Varsani, A.; Roumagnac, P.; Botha, G.; Maslamoney, S.; Schwab, T.; Kelz, Z.; Kumar, V.; Murrell, B. RDP5: A computer program for analyzing recombination in, and removing signals of recombination from, nucleotide sequence datasets. Virus Evol. 2021, 7, veaa087. [Google Scholar] [CrossRef]
- Anonymous. Package of Practice: Papaya; University of Horticultural Sciences: Bagalkot, India, 2013; ISBN 978-81-922104-1-4. Available online: http://uhsbagalkot.edu.in/downloads/Horticulture_POP.pdf (accessed on 10 May 2019).
- Sheoran, O.P.; Tonk, D.S.; Kaushik, L.S.; Hasija, R.C.; Pannu, R.S. Statistical Software Package for Agricultural Research Workers. Recent Advances in Information Theory, Statistics & Computer Applications by DS Hooda & RC Hasija Department of Mathematics Statistics; CCS HAU: Hisar, India, 1998; pp. 139–143. [Google Scholar]
- Azad, M.; Kalam, A.; Amin, L.; Sidik, N.M. Gene Technology for papaya ringspot virus disease management. Sci. World J. 2014, 2014, 768038. [Google Scholar] [CrossRef]
- Ventura, J.A.; Costa, H.; da Tatagiba, J.S. Papaya diseases and integrated control. In Diseases of Fruits and Vegetables: Volume II; Naqvi, S.A.M.H., Ed.; Springer: Dordrecht, The Netherlands, 2004; pp. 201–268. [Google Scholar] [CrossRef]
- Sharma, S.K.; Tripathi, S. Papaya ringspot virus-P: Overcoming limitations of resistance breeding in Carica papaya L. In Plant Virus–Host Interaction; Academic Press: Cambridge, MA, USA, 2014; pp. 177–194. [Google Scholar] [CrossRef]
- Castillo, X.O.; Fermin, G.; Tabima, J.; Rojas, Y.; Tennant, P.F.; Fuchs, M.; Sierra, R.; Bernal, A.J.; Restrepo, S. Phylogeography and molecular epidemiology of papaya ringspot virus. Virus Res. 2011, 159, 132–140. [Google Scholar] [CrossRef]
- Babu, K.S.; Banerjee, A. Biological and molecular evidence of papaya ringspot virus pathotype P from mid-hills of Meghalaya, India. Indian Phytopathol. 2018, 71, 611–620. [Google Scholar] [CrossRef]
- Maina, S.; Barbetti, M.J.; Edwards, O.R.; Minemba, D.; Areke, M.W.; Jones, R.A.C. Genetic connectivity between papaya ringspot virus genomes from Papua New Guinea and Northern Australia, and new recombination insights. Plant Dis. 2019, 103, 737–747. [Google Scholar] [CrossRef] [Green Version]
- Kalleshwaraswamy, C.M.; Kumar, N.K.K. Transmission Efficiency of papaya ringspot virus by three aphid species. Phytopathology 2008, 98, 541–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamim, I.; Al Rwahnih, M.; Borth, W.B.; Suzuki, J.Y.; Melzer, M.J.; Wall, M.M.; Green, J.C.; Hu, J.S. Papaya ringspot virus isolates from papaya in Bangladesh: Detection, characterization, and distribution. Plant Dis. 2019, 103, 2920–2924. [Google Scholar] [CrossRef] [PubMed]
- Taylor, L.R. Distribution of virus disease and the migrant vector aphid. In Plant Virus Epidemics: Monitoring, Modelling and Predicting Outbreaks; McLean, G.D., Garrett, R.G., Ruesink, W.G., Eds.; Academic Press: Sydney, Australia, 1986; pp. 35–57. [Google Scholar]
- Silva-Rosales, L.; Becerra-Leor, N.; Ruiz-Castro, S.; Téliz-Ortiz, D.; Noa-Carrazana, J.C. Coat protein sequence comparisons of three mexican isolates of papaya ringspot virus with other geographical isolates reveal a close relationship to American and Australian isolates. Arch. Virol. 2000, 145, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.J.; Antoniw, J.F.; Beaudoin, F. Overview and analysis of the polyprotein cleavage sites in the family Potyviridae. Mol. Plant Pathol. 2005, 6, 471–487. [Google Scholar] [CrossRef]
- Parameswari, B.; Mangrauthia, S.K.; Praveen, S.; Jain, R.K. Complete genome sequence of an isolate of papaya ringspot virus from India. Arch. Virol. 2007, 152, 843–845. [Google Scholar] [CrossRef]
- Pasin, F.; Simón-Mateo, C.; García, J.A. The hypervariable amino-terminus of P1 Protease modulates potyviral replication and host defense responses. PLoS Pathog. 2014, 10, e1003985. [Google Scholar] [CrossRef]
- Gorane, A.; Verma, R.; Naik, A.; Nikam, T.; Ade, A.; Mahapatro, G.; Tripathi, S. Characterization of a severe isolate of papaya ringspot virus from papaya in Western India. J. Plant Pathol. 2019, 101, 1203–1209. [Google Scholar] [CrossRef]
- Mangrauthia, S.K.; Parameswari, B.; Jain, R.K.; Praveen, S. Role of genetic recombination in the molecular architecture of papaya ringspot virus. Biochem. Genet. 2008, 46, 835–846. [Google Scholar] [CrossRef]
- Maina, S.; Coutts, B.A.; Edwards, O.R.; de Almeida, L.; Ximenes, A.; Jones, R.A. Papaya ringspot virus populations from East Timorese and Northern Australian Cucurbit crops: Biological and molecular properties, and absence of genetic connectivity. Plant Dis. 2017, 101, 985–993. [Google Scholar] [CrossRef] [Green Version]
- Kehoe, M.A.; Coutts, B.A.; Buirchell, B.J.; Jones, R.A. Split Personality of a Potyvirus: To specialize or not to specialize? PLoS ONE 2014, 9, e105770. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.-C.; Wu, X.-Y.; Zhang, Y.-M.; Wu, P.; Wu, X.-Z.; Liu, L.-W.; Wang, Q.; Hang, Y.-Y.; Yang, J.-Y.; Shao, Z.-Q. A genomic survey of thirty soybean-infecting bean common mosaic virus (BCMV) isolates from China pointed BCMV as a potential threat to soybean production. Virus Res. 2014, 191, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Mangrauthia, S.K.; Parameswari, B.; Praveen, S.; Jain, R.K. Comparative genomics of papaya ringspot virus pathotypes P and W from India. Arch. Virol. 2009, 154, 727–730. [Google Scholar] [CrossRef]
- Sultana, S.; Roy, B.; Sherpa, A.R. The complete sequence of a papaya ringspot virus (PRSV) isolate from West Bengal, India infecting papaya and study of genetic variation. Eur. J. Plant Pathol. 2021, 159, 203–210. [Google Scholar] [CrossRef]
- Dietrich, C.; Miller, J.; Mckenzie, G.; Palkovics, L.; Balázs, E.; Palukaitis, P.; Maiss, E. No recombination detected in artificial potyvirus mixed infections and between potyvirus derived transgenes and heterologous challenging potyviruses. Environ. Biosaf. Res. 2007, 6, 207–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escriu, F.; Fraile, A.; García-Arenal, F. Constraints to genetic exchange support gene coadaptation in a tripartite RNA virus. PLoS Pathog. 2007, 3, e8. [Google Scholar] [CrossRef] [Green Version]
- Saleem, A.; Ali, Z.; Yeh, S.-D.; Saeed, W.; Binat Imdad, A.; Akbar, M.F.; Goodman, R.E.; Naseem, S. Genetic variability and evolutionary dynamics of a typical papaya ringspot virus infecting papaya. PLoS ONE 2021, 16, e0258298. [Google Scholar] [CrossRef]
- Chin, M.; Rojas, Y.; Moret, J.; Fermin, G.; Tennant, P.; Gonsalves, D. Varying genetic diversity of papaya ringspot virus isolates from two time-separated outbreaks in Jamaica and Venezuela. Arch. Virol. 2007, 152, 2101–2106. [Google Scholar] [CrossRef]
- Taillebois, E.; Cartereau, A.; Jones, A.K.; Thany, S.H. Neonicotinoid insecticides mode of action on insect nicotinic acetylcholine receptors using binding studies. Pestic. Biochem. Physiol. 2018, 151, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.; Preston, S.; Song, H.; Jabbar, A.; Liu, Y.; Baell, J.; Hofmann, A.; Hutchinson, D.; Wang, T.; Koehler, A.V. Assessing the anthelmintic activity of pyrazole-5-carboxamide derivatives against Haemonchus Contortus. Parasit. Vectors 2017, 10, 272. [Google Scholar] [CrossRef] [Green Version]
- Okada, I.; Okui, S.; Fukuchi, T.; Yoshiya, K. Synthesis and insecticidal activity of n-(tolyloxybenzyl)-pyrazolecarboxamide derivatives. J. Pestic. Sci. 1999, 24, 393–396. [Google Scholar] [CrossRef] [Green Version]
- Kalyan, R.K.; Saini, D.P.; Babu, S.R. Evaluation of different doses of tolfenpyrad against aphids and thrips in cotton. J. Cotton Res. Dev. 2014, 28, 293–296. [Google Scholar]
- Priyanka, N.; Shaila, O.; Anurdha, M.; Rani, V.D.; Rajashekhar, M. Efficacy of newer insecticides against the major sucking pests of groundnut (Arachis hypogea L.). Int. J. Environ. Clim. Chang. 2022, 12, 557–564. [Google Scholar] [CrossRef]
- Bajpai, N.K.; Swami, H.; Ameta, K.D. Bio-efficacy of tolfenpyrad against diamond back moth (Plutella xylostella Linn.) infesting cabbage. Curr. Adv. Agric. Sci. 2014, 6, 79–81. [Google Scholar]
- Bajpai, N.K.; Swami, H.; Tripathi, N.N. Bioefficacy of tolfenpyrad (15% EC) against aphid Brevicoryne brassicae L. infesting cabbage in Rajasthan. Indian J. Entomol. 2013, 75, 208–211. [Google Scholar]
- Patra, B. Bioefficacy and phytotoxicity study of tolfenpyrad 15% EC, a novel pyrazole insecticide for management of Plutella Xylostella (L.) and Brevicoryne Brassicae L. Infesting cabbage and its effect on coccinellid predator. J. Entomol. Res. 2017, 41, 387–394. [Google Scholar] [CrossRef]
- Chavada, K.M.; Godhani, P.H.; Bhatt, N.A.; Patel, H.B.; Sapteshwriya, S.V. Bioefficacy of insecticides against aphid, Lipaphis Erysimi infesting cabbage. Int. J. Plant Prot. 2021, 48, 4. [Google Scholar]
- Bajpai, N.K.; Swami, H.; Jeengar, K.L. Bio-efficacy of tolfenpyrad against aphid (Aphisgossypii Glover) infesting okra in Rajasthan. Curr. Adv. Agric. Sci. 2013, 5, 81–83. [Google Scholar]
- Mallick, S.; Chand, P.; Mandal, S.K. Tolfenpyrad 15% EC, a new pyrazole insecticide, against sucking pests of okra and its impact on natural enemies. Indian J. Entomol. 2016, 78, 51–55. [Google Scholar] [CrossRef]
- Shivaleela, G.; Chowdary, R. Efficacy of new insecticide chemistry tolfenpyrad 15% EC against insect pests of cucumber (Cucumis Sativus L.). J. Entomol. Zool. Stu. 2020, 8, 879–884. [Google Scholar]
- Babu, S.R.; Singh, V. Bioefficacy of tolfenpyrad 15 EC against hopper complex in mango. Pest. Manag. Eortic. Ecsyst. 2014, 20, 22–25. [Google Scholar]
- Shewale, C.P.; Borad, P.K. Efficacy of chemical insecticides against fennel aphid, Hyadaphis coriandri Das infesting fennel (Foeniculum vulgare L.). J. Entomol. Zool. Stu. 2020, 8, 2362–2666. [Google Scholar]
- Raju, K.A.; Patil, C.S.; Kulkarni, S.R.; Kulkarni, S.S.; Walunj, A.R.; Deore, B.V. Bio-efficacy of newer insecticides against pomegranate aphid, Aphis Punicae. J. Pharmacogn. Phytochem. 2021, 10, 2685–2688. [Google Scholar]
- Samara, R.; Lowery, T.D.; Stobbs, L.W.; Vickers, P.M.; Bittner, L.A. Assessment of the effects of novel insecticides on green peach aphid (Myzus Persicae) feeding and transmission of turnip mosaic virus (TuMV). Pest. Manag. Sci. 2021, 77, 1482–1491. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, P.; Nauen, R.; Schindler, M.; Elbert, A. Overview of the status and global strategy for neonicotinoids. J. Agric. Food Chem. 2011, 59, 2897–2908. [Google Scholar] [CrossRef] [PubMed]
- Castle, S.; Palumbo, J.; Merten, P.; Cowden, C.; Prabhaker, N. Effects of foliar and systemic insecticides on whitefly transmission and incidence of cucurbit yellow stunting disorder virus. Pest. Manag. Sci. 2017, 73, 1462–1472. [Google Scholar] [CrossRef]
- Patel, V.; Saxena, S.; Lund, C.; Thornicroft, G.; Baingana, F.; Bolton, P.; Chisholm, D.; Collins, P.Y.; Cooper, J.L.; Eaton, J.; et al. The lancet commission on global mental health and sustainable development. Lancet 2018, 392, 1553–1598. [Google Scholar] [CrossRef] [Green Version]
- Amine, H.M.; Anber, H.A.; Abu-Shaishaa, A.R.; El-Rahman, A. efficacy of certain insecticides and mineral oil in controlling aphid, Aphis Gossypii Glov. and papaya ringspot virus in squash at Kafr El-Sheikh Governorate. J. Plant Prot. Pathol. 2022, 13, 141–146. [Google Scholar] [CrossRef]
- Sousa, T.; Cotas, J.; Pacheco, D.; Bahcevandziev, K.; Gonçalves, A.M.M.; Pereira, L. Can Seaweeds be used as immunity boosters? In Sustainable Global Resources of Seaweeds; Ranga Rao, A., Ravishankar, G.A., Eds.; Springer International Publishing: Cham, Switzerland, 2022; Volume 1, pp. 421–431. [Google Scholar] [CrossRef]
- Tang, F.; Li, L.; Chen, D. Mesoporous silica nanoparticles: Synthesis, biocompatibility and drug delivery. Adv. Mater. 2012, 24, 1504–1534. [Google Scholar] [CrossRef]
- Choi, H.W.; Klessig, D.F. DAMPs, MAMPs, and NAMPs in plant innate immunity. BMC Plant Biol. 2016, 16, 232. [Google Scholar] [CrossRef] [Green Version]
- Ali, O.; Ramsubhag, A.; Jayaraman, J. Application of extracts from caribbean seaweeds improves plant growth and yields and increases disease resistance in tomato and sweet pepper plants. Phytoparasitica 2022, 50, 1–19. [Google Scholar] [CrossRef]
- Pardee, K.I.; Ellis, P.; Bouthillier, M.; Towers, G.H.; French, C.J. Plant virus inhibitors from marine algae. Canad. J. Bot. 2004, 82, 304–309. [Google Scholar] [CrossRef] [Green Version]
- Ghannam, A.; Abbas, A.; Alek, H.; Al-Waari, Z.; Al-Ktaifani, M. Enhancement of local plant immunity against tobacco mosaic virus infection after treatment with sulphated-carrageenan from red alga (Hypnea musciformis). Physiol. Mol. Plant Pathol. 2013, 84, 19–27. [Google Scholar] [CrossRef]
- Alburquerque, N.; Faize, L.; Faize, M.; Nortes, M.D.; Bernardeau, J.; Fernandez, J.M.R.; Burgos, L. Towards the valorization of the invasive seaweeds Caulerpa Cylindracea and Asparagopsis taxiformis in the Mediterranean sea: Applications for in vitro plant regeneration and crop protection. J. Appl. Phycol. 2019, 31, 1403–1413. [Google Scholar] [CrossRef]
- El-Sawy, M.M.; Elsharkawy, M.M.; Abass, J.M.; Kasem, M.H. Antiviral Activity of 2-nitromethyl phenol, zinc nanoparticles and seaweed extract against cucumber mosaic virus (CMV) in eggplant. J. Virol. Antivir. Res. 2017, 6, 2. [Google Scholar] [CrossRef]
- Madhusudhan, K.N.; Vinayarani, G.; Deepak, S.A.; Niranjana, S.R.; Prakash, H.S.; Singh, G.P.; Sinha, A.K.; Prasad, B.C. Antiviral activity of plant extracts and other inducers against tobamoviruses infection in bell pepper and tomato plants. Int. J. Plant Pathol. 2011, 2, 35–42. [Google Scholar] [CrossRef]
- Abu-Ghannam, N.; Rajauria, G. Antimicrobial activity of compounds isolated from algae. In Functional Ingredients from Algae for Foods and Nutraceuticals; Elsevier: Amsterdam, The Netherlands, 2013; pp. 287–306. ISBN 978-0-85709-512-1. [Google Scholar]
- Arioli, T.; Mattner, S.W.; Winberg, P.C. Applications of seaweed extracts in australian agriculture: Past, present and future. J. Appl. Phycol. 2015, 27, 2007–2015. [Google Scholar] [CrossRef] [Green Version]
- Gonsalves, C.; Cai, W.; Tennant, P.; Gonsalves, D. Effective development of papaya ringspot virus resistant papaya with untranslatable coat protein gene using a modified microprojectile transformation method. Acta Hortic. 1997, 461, 311–314. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
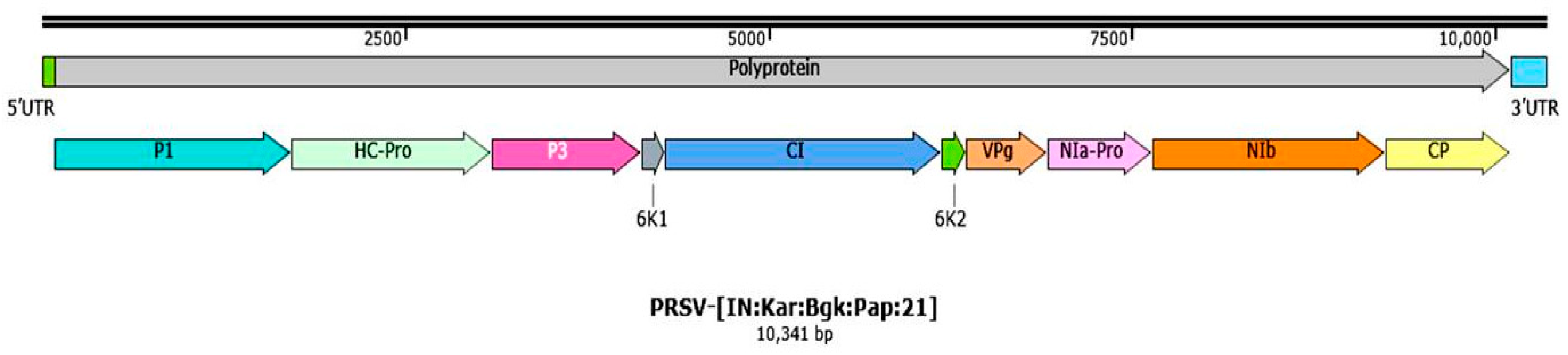
| Sl. No. | Genomic Regions | Position in Genome (Start-Stop Codon) | Predicted ORF’s Size (nt) | Predicted Size of Protein (No. of Amino Acids) | Predicted Protein Weight (kDa) | Predicted Cleavage Site | Strand Orientation | Direction with Start Codon |
|---|---|---|---|---|---|---|---|---|
| 1 | Genome length (nt) | 1–10,341 | - | - | - | - | - | - |
| 2 | Polyprotein | 86–10,114 | 10,029 | 3342 | 380.24 | - | +ve | Clockwise |
| 3 | 5′UTR a | 1–85 | 85 | - | - | - | - | - |
| 4 | P1 a | 86–1723 | 1638 | 546 | 62.3 | MEQY/N | +ve | Clockwise |
| 5 | HC-Pro a | 1724–3094 | 1371 | 457 | 52.12 | HYIVG/G | +ve | Clockwise |
| 6 | P3 | 3095–4129 | 1035 | 345 | 39.9 | VIHQ/A | +ve | Clockwise |
| 7 | 6K1 | 4130–4285 | 156 | 52 | 6.01 | VYHQ/S | +ve | Clockwise |
| 8 | CI a | 4286–6190 | 1905 | 635 | 71.35 | VYHQ/G | +ve | Clockwise |
| 9 | 6K2 | 6191–6361 | 171 | 57 | 6.36 | VFHQ/G | +ve | Clockwise |
| 10 | VPg a | 6362–6928 | 567 | 189 | 21.33 | VHHE/G | +ve | Clockwise |
| 11 | NIa-Pro a | 6929–7642 | 714 | 238 | 26.51 | VFEQ/S | +ve | Clockwise |
| 12 | Nib a | 7643–9253 | 1611 | 537 | 61.82 | VYHQ/S | +ve | Clockwise |
| 13 | CP a | 9254–10,114 | 861 | 286 | 32.72 | - | +ve | Clockwise |
| 14 | 3′UTR | 10,115–10,341 | 227 | - | - | - | - | - |
| 15 | Name assigned based on NSI | Papaya ringspot virus-[India:Karnataka:Bagalkote:Papaya:2021] | ||||||
| 16 | Virus acronyms | PRSV-[IN:Kar:Bgk:Pap:21] | ||||||
| 17 | Accession no. | OL677454 | ||||||
| Virus | Events | Breakpoint (nt) | Region/s | Recombination Parents | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Begin | End | Major Parent | Minor Parent | RDP | Geneconv | BootScan | MaxChi | Chimaera | SiScan | 3Seq | |||
| PRSV-[IN:KAR:BGK:Pap:21] OL677454 | 1 | 1 | 1031 | 5′ UTR to P1 | PRSV-[EC:Mild-Los-Rios:Pap:18] MT747167 | PRSV-[IN:Meg:Umi:Pap:15] MF356497 | 3.251 × 10−6 | - * | - | 1.140 × 10−7 | 5.526 × 10−9 | 1.026 × 10−6 | 6.155 × 10−12 |
| 10,328 | 10,341 | 3′ UTR | |||||||||||
| 2 | 1373 | 1710 | P1 | PRSV-[IN:TS:Hyd:HYD:Pap:15] KP743981 | PRSV-[IN:MH:PM-I:Pap:15]MF405296 | 3.407 × 10−2 | - | - | 6.918 × 10−4 | - | - | 7.044 × 10−3 | |
| 3 | 7327 | 9061 | NIa-Pro to NIb | PRSV-[IN:TS:Hyd:HYD:Pap:15] KP743981 | PRSV-[IN:MH:PM-I:Pap:15] MF405296 | 2.677 × 10−47 | 2.990 × 10−41 | - | 8.308 × 10−16 | 2.461 × 10−16 | 2.373 × 10−21 | 6.155 × 10−12 | |
| 4 | 9205 | 10,075 | NIb to CP | PRSV-[TW:TXG:Pap:18] NC001785 | PRSV-[IN:PRSVR3:Pap:13] KJ755852 | 3.041 × 10−5 | - | - | 5.194 × 10−5 | 1.131 × 10−5 | 5.576 × 10−3 | 4.050 × 10−8 | |
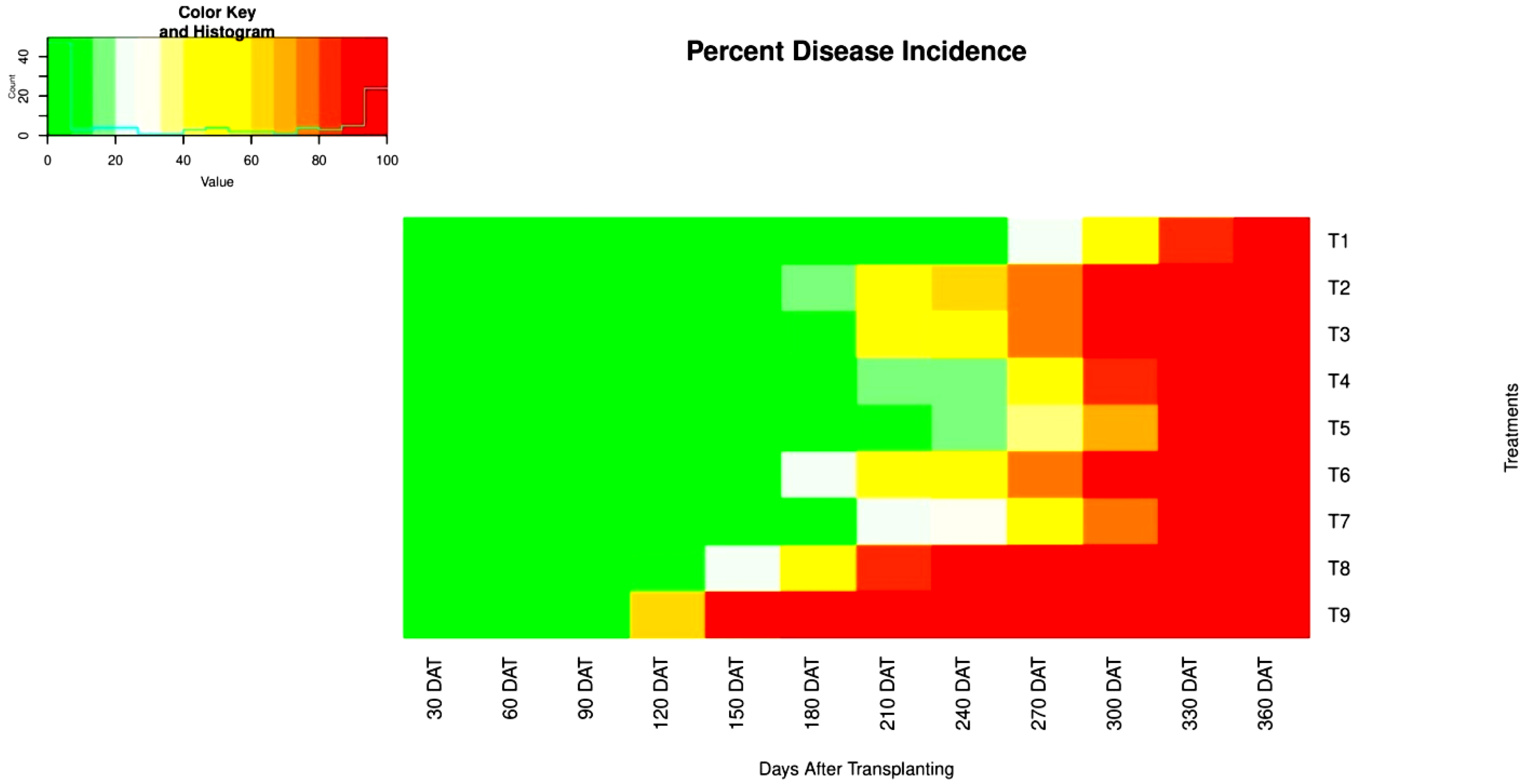
| Disease Incidence (%) at | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatments | 30 DAT * | 60 DAT | 90 DAT | 120 DAT | 150 DAT | 180 DAT | 210 DAT | 240 DAT | 270 DAT | 300 DAT | 330 DAT | 360 DAT |
| T1 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.49 | 5.19 | 24.07 | 50.90 | 84.25 | 100 |
| (0.00) ** | (0.00) | (0.00) | (0.00) | (0.00) | (0.00) | (4.75) | (10.79) | (29.31) | (45.59) | (70.89) | (90.00) | |
| T2 | 0.00 | 0.00 | 0.00 | 0.00 | 4.00 | 19.65 | 54.15 | 61.56 | 79.39 | 92.82 | 100 | 100 |
| (0.00) | (0.00) | (0.00) | (0.00) | (8.21) | (25.99) | (47.44) | (51.93) | (67.49) | (78.99) | (90.00) | (90.00) | |
| T3 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 9.58 | 43.04 | 47.79 | 74.07 | 89.33 | 100 | 100 |
| (0.00) | (0.00) | (0.00) | (0.00) | (0.00) | (16.23) | (40.61) | (43.63) | (64.50) | (74.28) | (90.00) | (90.00) | |
| T4 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2.61 | 15.38 | 19.54 | 46.28 | 81.57 | 100 | 100 |
| (0.00) | (0.00) | (0.00) | (0.00) | (0.00) | (5.42) | (20.92) | (25.90) | (42.83) | (65.18) | (90.00) | (90.00) | |
| T5 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 10.97 | 13.75 | 39.81 | 71.88 | 100 | 100 |
| (0.00) | (0.00) | (0.00) | (0.00) | (0.00) | (0.00) | (17.56) | (21.55) | (38.37) | (58.34) | (90.00) | (90.00) | |
| T6 | 0.00 | 0.00 | 0.00 | 0.00 | 3.65 | 25.10 | 46.04 | 48.94 | 74.29 | 90.50 | 100 | 100 |
| (0.00) | (0.00) | (0.00) | (0.00) | (7.83) | (29.99) | (42.72) | (44.39) | (59.79) | (77.35) | (90.00) | (90.00) | |
| T7 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2.19 | 23.60 | 28.22 | 52.31 | 75.56 | 100 | 100 |
| (0.00) | (0.00) | (0.00) | (0.00) | (0.00) | (5.54) | (28.09) | (31.71) | (46.81) | (60.65) | (90.00) | (90.00) | |
| T8 | 0.00 | 0.00 | 0.00 | 10.63 | 25.56 | 55.56 | 81.47 | 87.03 | 100 | 100 | 100 | 100 |
| (0.00) | (0.00) | (0.00) | (17.19) | (30.18) | (48.38) | (68.84) | (74.80) | (90.00) | (90.00) | (90.00) | (90.00) | |
| T9 | 0.00 | 0.00 | 0.57 | 65.61 | 90.38 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| (0.00) | (0.00) | (2.49) | (54.19) | (73.76) | (90.00) | (90.00) | (90.00) | (90.00) | (90.00) | (90.00) | (90.00) | |
| S Em ± | NS | NS | 0.53 | 1.43 | 2.22 | 2.35 | 3.85 | 3.04 | 5.13 | 3.48 | 2.17 | NS |
| C D at 5% | NS | NS | 1.51 | 4.09 | 6.35 | 6.72 | 11.02 | 8.68 | 14.66 | 9.94 | 6.21 | NS |
| Treatments | Plant Height (cm) at 270 DAT * | Internodal Length (cm) at 270 DAT | Plant Girth (cm) at 270 DAT | No. of Leaves per Plants at 270 DAT | No. of Days Taken for First Flowering | No. of Flowers at 270 DAT | No. of Days Taken from Flowering to First Fruit Set | No. of Days Taken to First Fruit Set to Harvest | No. of Fruits per Plant | Fruit Diameter (cm) | Fruit Length (cm) | Fruit Breadth (cm) | Fruit Cavity Diameter (cm) | Fruit Yield per Plant (Kg) | Fruit Yield per Hectare (t) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T1 | 225.93 | 4.11 | 40.45 | 29.50 | 93.01 | 73.06 | 94.01 | 125.64 | 26.09 | 13.13 | 27.78 | 14.43 | 6.81 | 57.86 | 178.56 |
| T2 | 204.98 | 3.60 | 37.44 | 23.19 | 105.33 | 32.05 | 103.90 | 110.44 | 16.22 | 9.89 | 20.25 | 9.93 | 6.07 | 26.18 | 80.79 |
| T3 | 206.43 | 3.71 | 38.26 | 27.19 | 100.83 | 58.26 | 101.07 | 118.44 | 23.03 | 11.43 | 24.75 | 11.36 | 6.53 | 45.83 | 141.44 |
| T4 | 214.18 | 4.10 | 39.48 | 28.29 | 99.31 | 56.70 | 95.93 | 123.50 | 22.43 | 12.05 | 24.69 | 12.18 | 6.62 | 42.45 | 131.01 |
| T5 | 216.71 | 4.10 | 39.68 | 28.67 | 95.94 | 59.88 | 96.38 | 123.88 | 24.21 | 12.78 | 25.42 | 12.55 | 6.73 | 52.16 | 160.97 |
| T6 | 205.79 | 3.49 | 37.75 | 26.35 | 101.93 | 54.40 | 100.85 | 114.42 | 21.69 | 10.82 | 22.93 | 11.04 | 6.40 | 40.76 | 125.80 |
| T7 | 214.49 | 3.73 | 39.12 | 27.31 | 100.19 | 56.67 | 101.43 | 115.97 | 21.75 | 11.21 | 24.39 | 11.56 | 6.45 | 41.75 | 128.83 |
| T8 | 160.95 | 3.39 | 34.05 | 20.54 | 105.47 | 20.73 | 111.15 | 98.33 | 5.66 | 8.88 | 17.78 | 9.46 | 5.94 | 6.43 | 19.84 |
| T9 | 135.53 | 3.28 | 20.95 | 19.80 | 106.81 | 15.15 | 115.18 | 98.58 | 1.19 | 8.53 | 15.06 | 8.91 | 5.63 | 0.81 | 2.51 |
| S Em ± | 1.79 | 0.04 | 0.18 | 0.21 | 0.17 | 0.70 | 0.46 | 0.46 | 0.54 | 0.13 | 0.19 | 0.10 | 0.05 | 0.79 | 2.45 |
| C D at 5% | 5.11 | 0.11 | 0.51 | 0.59 | 0.49 | 2.00 | 1.31 | 1.32 | 1.54 | 0.36 | 0.54 | 0.30 | 0.14 | 2.27 | 7.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Premchand, U.; Mesta, R.K.; Devappa, V.; Basavarajappa, M.P.; Venkataravanappa, V.; Narasimha Reddy, L.R.C.; Shankarappa, K.S. Survey, Detection, Characterization of Papaya Ringspot Virus from Southern India and Management of Papaya Ringspot Disease. Pathogens 2023, 12, 824. https://doi.org/10.3390/pathogens12060824
Premchand U, Mesta RK, Devappa V, Basavarajappa MP, Venkataravanappa V, Narasimha Reddy LRC, Shankarappa KS. Survey, Detection, Characterization of Papaya Ringspot Virus from Southern India and Management of Papaya Ringspot Disease. Pathogens. 2023; 12(6):824. https://doi.org/10.3390/pathogens12060824
Chicago/Turabian StylePremchand, Udavatha, Raghavendra K. Mesta, Venkatappa Devappa, Mantapla Puttappa Basavarajappa, Venkataravanappa Venkataravanappa, Lakshminarayana Reddy C. Narasimha Reddy, and Kodegandlu Subbanna Shankarappa. 2023. "Survey, Detection, Characterization of Papaya Ringspot Virus from Southern India and Management of Papaya Ringspot Disease" Pathogens 12, no. 6: 824. https://doi.org/10.3390/pathogens12060824
APA StylePremchand, U., Mesta, R. K., Devappa, V., Basavarajappa, M. P., Venkataravanappa, V., Narasimha Reddy, L. R. C., & Shankarappa, K. S. (2023). Survey, Detection, Characterization of Papaya Ringspot Virus from Southern India and Management of Papaya Ringspot Disease. Pathogens, 12(6), 824. https://doi.org/10.3390/pathogens12060824