
Lactate Dehydrogenase Inhibitors Suppress Borrelia burgdorferi Growth In Vitro



Abstract
:1. Introduction
2. Materials and Methods
2.1. Acquisition of LDH Inhibitors
2.2. Growth Inhibition Assays
2.3. Minimum Bactericidal Concentration (MBC) Assays
2.4. Data Analysis
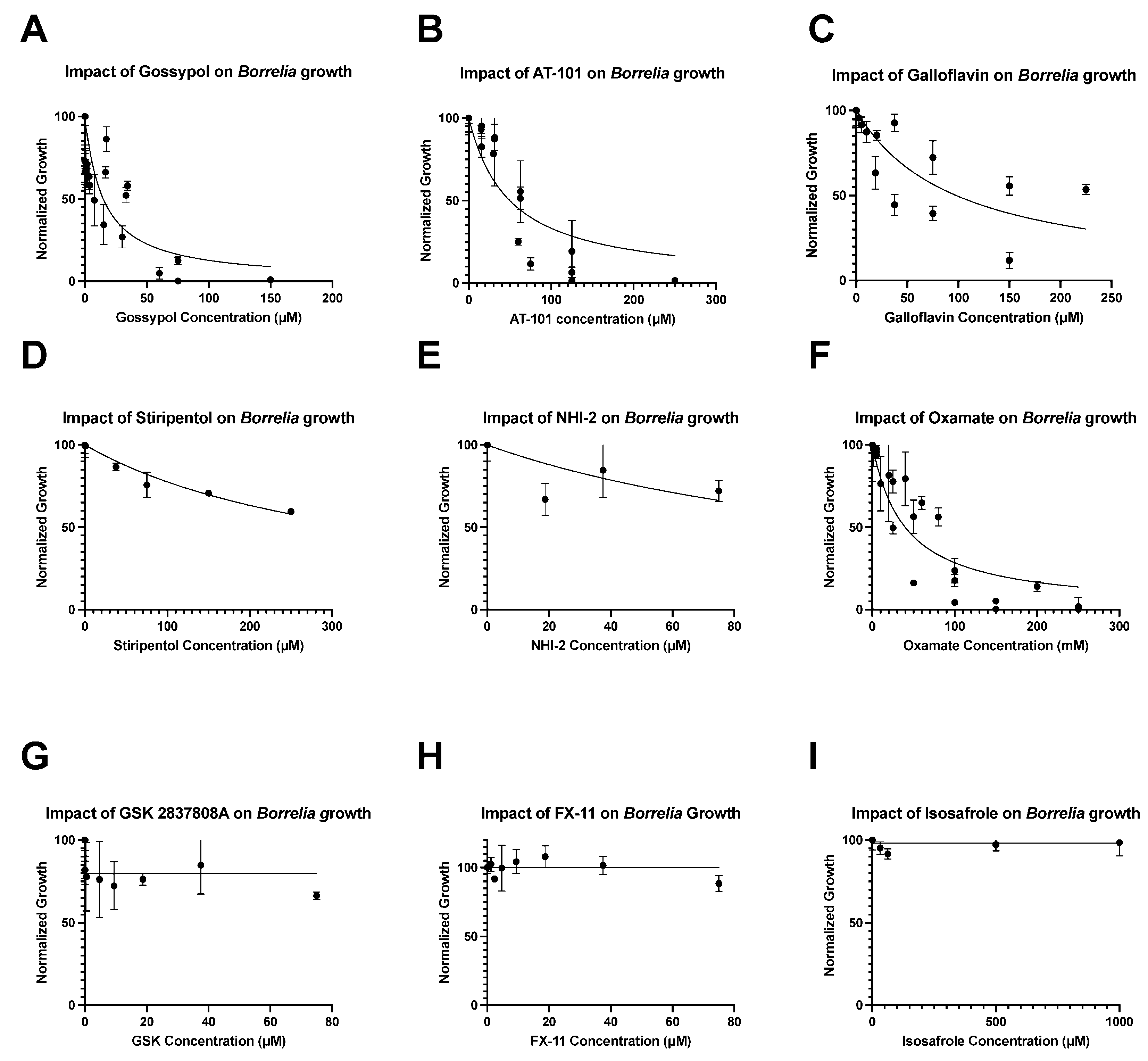
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eddens, T.; Kaplan, D.J.; Anderson, A.J.M.; Nowalk, A.J.; Campfield, B.T.; Eddens, T. Insights from the Geographic Spread of the Lyme Disease Epidemic. Clin. Infect. Dis. 2018, 68, 426–434. [Google Scholar] [CrossRef]
- Wormser, G.P.; Ramanathan, R.; Nowakowski, J.; McKenna, D.; Holmgren, D.; Visintainer, P.; Dornbush, R.; Singh, B.; Nadelman, R.B. Duration of Antibiotic Therapy for Early Lyme Disease: A Randomized, Double-Blind, Placebo-Controlled Trial. Ann. Intern. Med. 2003, 138, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Nadelman, R.B.; Nowakowski, J.; Fish, D.; Falco, R.C.; Freeman, K.; McKenna, D.; Welch, P.; Marcus, R.; Agüero-Rosenfeld, M.E.; Dennis, D.T.; et al. Prophylaxis with Single-Dose Doxycycline for the Prevention of Lyme Disease after an Ixodes scapularis Tick Bite. N. Engl. J. Med. 2001, 345, 79–84. [Google Scholar] [CrossRef]
- Stanek, G.; Strle, F. Lyme borreliosis–from tick bite to diagnosis and treatment. FEMS Microbiol. Rev. 2018, 42, 233–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warshafsky, S.; Lee, D.H.; Francois, L.K.; Nowakowski, J.; Nadelman, R.B.; Wormser, G.P. Efficacy of antibiotic prophylaxis for the prevention of Lyme disease: An updated systematic review and meta-analysis. J. Antimicrob. Chemother. 2010, 65, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Bian, J.; Deng, Y.; Smith, A.; Nunez, R.E.; Li, M.B.; Pal, U.; Yu, A.-M.; Qiu, W.; Ealick, S.E.; et al. Lyme disease spirochaete Borrelia burgdorferi does not require thiamin. Nat. Microbiol. 2016, 2, 16213. [Google Scholar] [CrossRef] [Green Version]
- Corona, A.; Schwartz, I. Borrelia burgdorferi: Carbon Metabolism and the Tick-Mammal Enzootic Cycle. Microbiol. Spectr. 2015, 3, 167–184. [Google Scholar] [CrossRef] [Green Version]
- Xing, B.-C.; Wang, C.; Ji, F.-J.; Zhang, X.-B. Synergistically suppressive effects on colorectal cancer cells by combination of mTOR inhibitor and glycolysis inhibitor, Oxamate. Int. J. Clin. Exp. Pathol. 2018, 11, 4439–4445. [Google Scholar]
- Zhang, H.; Guo, F.; Zhu, G. Cryptosporidium Lactate Dehydrogenase Is Associated with the Parasitophorous Vacuole Membrane and Is a Potential Target for Developing Therapeutics. PLoS Pathog. 2015, 11, e1005250. [Google Scholar] [CrossRef]
- Le, A.; Cooper, C.R.; Gouw, A.M.; Dinavahi, R.; Maitra, A.; Deck, L.M.; Royer, R.E.; Vander Jagt, D.L.; Semenza, G.L.; Dang, C.V. Inhibition of lactate dehydrogenase A induces oxidative stress and inhibits tumor progression. Proc. Natl. Acad. Sci. USA 2010, 107, 2037–2042. [Google Scholar] [CrossRef]
- Wu, Y.W.; Chik, C.L.; Knazek, R.A. An In Vitro and In Vivo Study of Antitumor Effects of Gossypol on Human SW-13 Adrenocortical Carcinoma. Cancer Res. 1989, 49, 3754–3758. [Google Scholar] [PubMed]
- Urbańska, K.; Orzechowski, A. Unappreciated Role of LDHA and LDHB to Control Apoptosis and Autophagy in Tumor Cells. Int. J. Mol. Sci. 2019, 20, 2085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, B.; Schafer, X.L.; Ambeskovic, A.; Spencer, C.M.; Land, H.; Munger, J. Addiction to Coupling of the Warburg Effect with Glutamine Catabolism in Cancer Cells. Cell Rep. 2016, 17, 821–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg Effect: The Metabolic Requirements of Cell Proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Pinto, B.M. Human Lactate Dehydrogenase A Inhibitors: A Molecular Dynamics Investigation. PLoS ONE 2014, 9, e86365. [Google Scholar] [CrossRef] [Green Version]
- Vudriko, P.; Masatani, T.; Cao, S.; Terkawi, M.A.; Kamyingkird, K.; Mousa, A.A.; Moumouni, P.F.A.; Nishikawa, Y.; Xuan, X. Molecular and Kinetic Characterization of Babesia microti Gray Strain Lactate Dehydrogenase as a Potential Drug Target. Drug Target Insights 2014, 8, 31–38. [Google Scholar] [CrossRef]
- He, L.; Bastos, R.G.; Yu, L.; Laughery, J.M.; Suarez, C.E. Lactate Dehydrogenase as a Potential Therapeutic Drug Target to Control Babesia bigemina. Front. Cell. Infect. Microbiol. 2022, 12, 870852. [Google Scholar] [CrossRef]
- Granchi, C.; Paterni, I.; Rani, R.; Minutolo, F.; Kumar, V.; A Christopher, J.; Jeong, Y.; Kwon, D.; Hong, S.; Roychowdhury, A.; et al. Small-molecule inhibitors of human LDH5. Futur. Med. Chem. 2013, 5, 1967–1991. [Google Scholar] [CrossRef] [Green Version]
- Manerba, M.; Vettraino, M.; Fiume, L.; Di Stefano, G.; Sartini, A.; Giacomini, E.; Buonfiglio, R.; Roberti, M.; Recanatini, M. Galloflavin (CAS 568-80-9): A Novel Inhibitor of Lactate Dehydrogenase. Chemmedchem 2011, 7, 311–317. [Google Scholar] [CrossRef]
- Sada, N.; Lee, S.; Katsu, T.; Otsuki, T.; Inoue, T. Targeting LDH enzymes with a stiripentol analog to treat epilepsy. Science 2015, 347, 1362–1367. [Google Scholar] [CrossRef]
- Doherty, J.R.; Cleveland, J.L. Targeting lactate metabolism for cancer therapeutics. J. Clin. Investig. 2013, 123, 3685–3692. [Google Scholar] [CrossRef]
- Lea, M.A.; Guzman, Y.; Desbordes, C. Inhibition of Growth by Combined Treatment with Inhibitors of Lactate Dehydrogenase and either Phenformin or Inhibitors of 6-Phosphofructo-2-kinase/Fructose-2,6-bisphosphatase 3. Anticancer Res. 2016, 36, 1479–1488. [Google Scholar] [PubMed]
- Li, X.; Lu, W.; Hu, Y.; Wen, S.; Qian, C.; Wu, W.; Huang, P. Effective inhibition of nasopharyngeal carcinoma in vitro and in vivo by targeting glycolysis with oxamate. Int. J. Oncol. 2013, 43, 1710–1718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.-F.; Yu, Y.-W.; Tang, Z.-M.; Wang, M.-Z. Pharmacokinetics of (±)-, (+)-, and (−)-gossypol in humans and dogs. Clin. Pharmacol. Ther. 1986, 39, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Billiard, J.; Dennison, J.B.; Briand, J.; Annan, R.S.; Chai, D.; Colón, M.; Dodson, C.S.; Gilbert, S.A.; Greshock, J.; Jing, J.; et al. Quinoline 3-sulfonamides inhibit lactate dehydrogenase A and reverse aerobic glycolysis in cancer cells. Cancer Metab. 2013, 1, 19. [Google Scholar] [CrossRef] [Green Version]
- Olgiati, K.L.; Toscano, W.A. Kinetics of gossypol inhibition of bovine lactate dehydrogenase X. Biochem. Biophys. Res. Commun. 1983, 115, 180–185. [Google Scholar] [CrossRef]
- Caol, S.; Divers, T.; Crisman, M.; Chang, Y.-F. In vitro susceptibility of Borrelia burgdorferi isolates to three antibiotics commonly used for treating equine Lyme disease. BMC Veter Res. 2017, 13, 293. [Google Scholar] [CrossRef] [Green Version]
- Sicklinger, M.; Wienecke, R.; Neubert, U. In Vitro Susceptibility Testing of Four Antibiotics against Borrelia burgdorferi: A Comparison of Results for the Three Genospecies Borrelia afzelii, Borrelia garinii, and Borrelia burgdorferi Sensu Stricto. J. Clin. Microbiol. 2003, 41, 1791–1793. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M.; Marayati, R.; Moffitt, R.; Yeh, J.J. Hexokinase 2 promotes tumor growth and metastasis by regulating lactate production in pancreatic cancer. Oncotarget 2016, 8, 56081–56094. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, E.; Fennell, M.; Stafford, D.M. Modification of Tumour Glucose Metabolism for Therapeutic Benefit. Acta Oncol. 1995, 34, 429–433. [Google Scholar] [CrossRef]
- Liu, S.; Kulp, S.K.; Sugimoto, Y.; Jiang, J.; Chang, H.-L.; Dowd, M.K.; Wan, P.; Lin, Y.C. The (-)-enantiomer of gossypol possesses higher anticancer potency than racemic gossypol in human breast cancer. Anticancer Res. 2002, 22, 33–38. [Google Scholar] [PubMed]
- Dong, Y.; Mao, B.; Li, L.; Guan, H.; Su, Y.; Li, X.; Lian, Q.; Huang, P.; Ge, R.-S. Gossypol enantiomers potently inhibit human placental 3β-hydroxysteroid dehydrogenase 1 and aromatase activities. Fitoterapia 2016, 109, 132–137. [Google Scholar] [CrossRef]
- Lordelo, M.M.; Davis, A.J.; Calhoun, M.C.; Dowd, M.K.; Dale, N.M. Relative toxicity of gossypol enantiomers in broilers. Poult. Sci. 2005, 84, 1376–1382. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, M.C.; Naqvi, R.H.; Matlin, S.A.; Zhou, R.H.; Bialy, G.; Blye, R.P. Comparative anti-fertility effects of gossypol enantiomers in male hamsters. Int. J. Androl. 1987, 10, 619–623. [Google Scholar] [CrossRef]
- PubChem. 3-[[3-(Cyclopropylsulfamoyl)-7-(2,4-dimethoxypyrimidin-5-yl)quinolin-4-yl]amino]-5-(3,5-difluorophenoxy)benzoic Acid. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/71533725 (accessed on 21 January 2020).
- Suzuki, K.; Maeda, S.; Morokuma, K. Roles of Closed- and Open-Loop Conformations in Large-Scale Structural Transitions of l-Lactate Dehydrogenase. ACS Omega 2019, 4, 1178–1184. [Google Scholar] [CrossRef] [PubMed]
- Granchi, C.; Calvaresi, E.C.; Tuccinardi, T.; Paterni, I.; Macchia, M.; Martinelli, A.; Hergenrother, P.J.; Minutolo, F. Assessing the differential action on cancer cells of LDH-A inhibitors based on the N-hydroxyindole-2-carboxylate (NHI) and malonic (Mal) scaffolds. Org. Biomol. Chem. 2013, 11, 6588–6596. [Google Scholar] [CrossRef] [Green Version]
- Marlier, J.F.; Cleland, W.W.; Zeczycki, T.N. Oxamate Is an Alternative Substrate for Pyruvate Carboxylase from Rhizobium etli. Biochemistry 2013, 52, 2888–2894. [Google Scholar] [CrossRef]
- Thornburg, J.M.; Nelson, K.K.; Clem, B.F.; Lane, A.N.; Arumugam, S.; Simmons, A.; Eaton, J.W.; Telang, S.; Chesney, J. Targeting aspartate aminotransferase in breast cancer. Breast Cancer Res. 2008, 10, R84. [Google Scholar] [CrossRef]
- Saleh, S.R.; Attia, R.; Ghareeb, D.A. The Ameliorating Effect of Berberine-Rich Fraction against Gossypol-Induced Testicular Inflammation and Oxidative Stress. Oxidative Med. Cell. Longev. 2018, 2018, 1056173. [Google Scholar] [CrossRef]
- Santana, A.T.; Guelfi, M.; Medeiros, H.C.D.; A Tavares, M.; Bizerra, P.F.V.; Mingatto, F.E. Mechanisms involved in reproductive damage caused by gossypol in rats and protective effects of vitamin E. Biol. Res. 2015, 48, 43. [Google Scholar] [CrossRef] [Green Version]
- Ramsey, M.E.; Hyde, J.A.; Medina, D.N.; Lin, T.; Gao, L.; Lundt, M.E.; Li, X.; Norris, S.J.; Skare, J.T.; Hu, L.T. A high-throughput genetic screen identifies previously uncharacterized Borrelia burgdorferi genes important for resistance against reactive oxygen and nitrogen species. PLoS Pathog. 2017, 13, e1006225. [Google Scholar] [CrossRef] [PubMed]
- Bourret, T.J.; Boyle, W.K.; Zalud, A.K.; Valenzuela, J.G.; Oliveira, F.; Lopez, J.E. The relapsing fever spirochete Borrelia turicatae persists in the highly oxidative environment of its soft-bodied tick vector. Cell. Microbiol. 2018, 21, e12987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radolf, J.D.; Samuels, D.S. Borrelia: Molecular Biology, Host Interaction and Pathogenesis; Horizon Scientific Press: Norfolk, UK, 2010. [Google Scholar]
- Di Tommaso, P.; Moretti, S.; Xenarios, I.; Orobitg, M.; Montanyola, A.; Chang, J.-M.; Taly, J.-F.; Notredame, C. T-Coffee: A web server for the multiple sequence alignment of protein and RNA sequences using structural information and homology extension. Nucleic Acids Res. 2011, 39, W13–W17. [Google Scholar] [CrossRef] [PubMed]
- Notredame, C.; Higgins, D.G.; Heringa, J. T-coffee: A novel method for fast and accurate multiple sequence alignment. J. Mol. Biol. 2000, 302, 205–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Compound | IC50 | MIC | MBC |
|---|---|---|---|
| Gossypol (+/−) | 14 µM | 70.5 µM | 150 µM |
| AT-101 | 50 µM | 187.5 µM | 250 µM |
| Galloflavin | 98 µM | - | - |
| NHI-2 | 150 µM | - | - |
| Stiripentol | 350 µM | - | - |
| Oxamate | 38 mM | 125 mM | 200 mM |
| FX-11 | - | - | - |
| Isosafrole | - | - | - |
| GSK2837808A | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lynch, A.; Pearson, P.; Savinov, S.N.; Li, A.Y.; Rich, S.M. Lactate Dehydrogenase Inhibitors Suppress Borrelia burgdorferi Growth In Vitro. Pathogens 2023, 12, 962. https://doi.org/10.3390/pathogens12070962
Lynch A, Pearson P, Savinov SN, Li AY, Rich SM. Lactate Dehydrogenase Inhibitors Suppress Borrelia burgdorferi Growth In Vitro. Pathogens. 2023; 12(7):962. https://doi.org/10.3390/pathogens12070962
Chicago/Turabian StyleLynch, Adam, Patrick Pearson, Sergey N. Savinov, Andrew Y. Li, and Stephen M. Rich. 2023. "Lactate Dehydrogenase Inhibitors Suppress Borrelia burgdorferi Growth In Vitro" Pathogens 12, no. 7: 962. https://doi.org/10.3390/pathogens12070962
APA StyleLynch, A., Pearson, P., Savinov, S. N., Li, A. Y., & Rich, S. M. (2023). Lactate Dehydrogenase Inhibitors Suppress Borrelia burgdorferi Growth In Vitro. Pathogens, 12(7), 962. https://doi.org/10.3390/pathogens12070962