
Screening for Resistant Bacteria, Antimicrobial Resistance Genes, Sexually Transmitted Infections and Schistosoma spp. in Tissue Samples from Predominantly Vaginally Delivered Placentae in Ivory Coast and Ghana

 , , , , , , , , , and
, , , , , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Placenta Tissue Sample Collection and Storage
2.3. Culture-Based Assessments
2.4. Nucleic Acid Extraction and Amplification from Primary Sample Materials
2.5. Statistical Assessment
3. Results
3.1. Growth of Resistant Bacteria on Selective Agars after Broth Enrichment
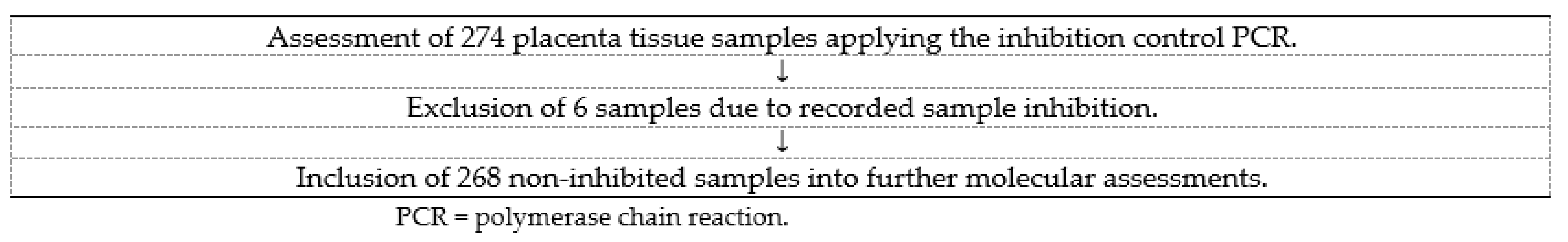
3.2. Samples Included in the Molecular Assessments
3.3. Molecular Detection of Genetic Resistance Determinants
3.4. Molecular Detection of Sexually Transmitted Infections
3.5. Molecular Detection of Schistosoma Mansoni Complex and Schistosoma Haematobium Complex
3.6. Associations with Epidemiological Data as Observed in the Epidemiological Assessment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
| PCR Target | CTX-M-Type Beta-Lactamase (Groups I–V) |
|---|---|
| Target gene | blaCTX-M |
| Detection limit | <102 copies/µL |
| Forward primer I | 5′-GCT-GGA-CTG-CCT-GCT-TCC-T-3′ |
| Forward primer II | 5′-TGC-CGA-AAT-CAT-GGG-TAG-TG-3′ |
| Forward primer III | 5′-CTA-CCC-ACA-TCG-TGG-GTT-GTC-3′ |
| Forward primer IV/V | 5′-ATT-CGG-GCC-GGC-TTA-CC-3′ |
| Reverse primer I | 5′-CGT-TGG-TGG-TGC-CAT-AGY-CA-3′ |
| Reverse primer II | 5′-TCG-TTG-GTG-GTG-CCA-TAA-TCT-3′ |
| Reverse primer III | 5′-GAT-GTC-ATT-CGT-CGT-ACC-ATA-ATC-A-3′ |
| Reverse primer IV | 5′-ATC-ATT-GGT-GGT-GCC-GTA-GYC-3′ |
| Reverse primer V | 5′-GCG-ATA-TCA-TTC-GTC-GTA-CCA-TAA-3′ |
| Probe and modifications | 5‘-VIC-CCG-CTG-CCG-GTC-TTA-TC-MGB-NFQ-3‘ |
| Positive control plasmid insert | 5′-CGC-AGC-CAG-CAT-TCG-GGC-CGG-CTT-ACC-GAC-GTC-GTG-GAC-TGT-GGG-TGA-TAA-GAC-CGG-CAG-CGG-CGA-CTA-CGG-CAC-CAC-CAA-TGA-TAT-TGC-GGT-GA-3′ |
| GenBank accession number of the insert | OM355481.1 |
| Reference | [59] |
| PCR target | VIM-type beta-lactamase |
| Target gene | blaVIM |
| Detection limit | <102 copies/µL |
| Forward primer | 5′-GAG-ATT-CCC-ACG-CAY-TCT-CTA-GA-3′ |
| Reverse primer | 5′-AAT-GCG-CAG-CAC-CAG-GAT-AG-3′ |
| Probe and modifications | 5′-JOE-ACG-CAG-TGC-GCT-TCG-GTC-CAG-T-BHQ1-3′ |
| Positive control plasmid insert | 5′-AGA-GGG-GAG-CGA-GAT-TCC-CAC-GCA-CTC-TCT-AGA-AGG-ACT-CTC-ATC-GAG-CGG-GGA-CGC-AGT-GCG-CTT-CGG-TCC-AGT-AGA-ACT-CTT-CTA-TCC-TGG-TGC-TGC-GCA-TTC-GAC-CGA-CAA-3′ |
| GenBank accession number of the insert | NG_050338.1 |
| Reference | [59] |
| PCR target | IMP-type beta-lactamase |
| Target gene | blaIMP |
| Detection limit | <102 copies/µL |
| Forward primer | 5′-GGC-GGA-ATA-GAG-TGG-CTT-AAT-TCT-C-3′ |
| Reverse primer I | 5′-GAA-TTT-TTA-GCT-TGT-ACT-TTA-CCG-TCT-TT-3′ |
| Reverse primer II | 5′-ATT-TTT-AGC-TTG-TAC-CTT-ACC-GTA-TT-3′ |
| Reverse primer III | 5′-TTT-GTA-GCT-TGC-ACC-TTA-TTG-TCT-TT-3′ |
| Probe I and modifications | 5′-FAM-ATG-CAT-CTG-AAT-TAA-C-MGB-TAMRA-3′ |
| Probe II and modifications | 5‘-FAM-TAT-*GCA-TCT-*GAA-T*TA-A*CA-AAT-*GA-TAMRA-3‘ |
| Positive control plasmid insert | 5′-CGA-CAG-CAC-GGG-CGG-AAT-AGA-GTG-GCT-TAA-TTC-TCA-ATC-TAT-CCC-CAC-GTA-TGC-ATC-TGA-ATT-AAC-AAA-TGA-ACT-TCT-TAA-AAA-AGA-CGG-TAA-AGT-ACA-AGC-TAA-AAA-TTC-ATT-TAG-CGG-AG-3′ |
| GenBank accession number of the insert | NG_049212.1 |
| Reference | [59] |
| PCR target | NDM-type beta-lactamase |
| Target gene | blaNDM |
| Detection limit | <102 copies/µL |
| Forward primer | 5′-CAT-TAG-CCG-CTG-CAT-TGA-TG-3′ |
| Reverse primer | 5′-GTC-GCC-AGT-TTC-CAT-TTG-CT-3′ |
| Probe and modifications | 5′-ROX-CAT-GCC-CGG-TGA-AAT-CCG-CC-BHQ2-3′ |
| Positive control plasmid insert | 5′-CTG-AGC-ACC-GCA-TTA-GCC-GCT-GCA-TTG-ATG-CTG-AGC-GGG-TGC-ATG-CCC-GGT-GAA-ATC-CGC-CCG-ACG-ATT-GGC-CAG-CAA-ATG-GAA-ACT-GGC-GAC-CAA-CGG-TTT-GGC-3′ |
| GenBank accession number of the insert | NG_088409.1 |
| Reference | [59] |
| PCR target | KPC-type beta-lactamase |
| Target gene | blaKPC |
| Detection limit | <102 copies/µL |
| Forward primer | 5′-TGC-AGA-GCC-CAG-TGT-CAG-TTT-3′ |
| Reverse primer | 5′-CGC-TCT-ATC-GGC-GAT-ACC-A-3′ |
| Probe and modifications | 5′-Cy5-TTC-CGT-CAC-GGC-GCG-CG-BHQ2-3′ |
| Positive control plasmid insert | 5′-GGC-CTT-CAT-GCG-CTC-TAT-CGG-CGA-TAC-CAC-GTT-CCG-TCT-GGA-CCG-CTG-GGA-GCT-GGA-GCT-GAA-CTC-CGC-CAT-CCC-AGG-CGA-TGC-GCG-CTA-TAC-CTC-ATC-GCC-GCG-CGC-CGT-GAC-GGA-AAG-CTT-ACA-AAA-ACT-GAC-ACT-GGG-CTC-TGC-ACT-GGC-TGC-GC-3′ |
| GenBank accession number of the insert | NG_067225.1 |
| Reference | [59] |
| PCR target | OXA-23-like-type beta-lactamase |
| Target gene | blaOXA-23-like |
| Detection limit | <102 copies/µL |
| Forward primer | 5′-TAA-ATG-GAA-GGG-CGA-GAA-3′ |
| Reverse primer | 5′-ACC-TGC-TGT-CCA-ATT-TCA-G-3′ |
| Probe and modifications | 5′-FAM-CCA-TGA-AGC-TTT-CTG-CAG-TCC-CAG-TC-TAMRA-3′ |
| Positive control plasmid insert | 5′-ATG-AAA-TAT-TTA-AAT-GGA-AGG-GCG-AGA-AAA-GGT-CAT-TTA-CCG-CTT-GGG-AAA-AAG-ACA-TGA-CAC-TAG-GAG-AAG-CCA-TGA-AGC-TTT-CTG-CAG-TCC-CAG-TCT-ATC-AGG-AAC-TTG-CGC-GAC-GTA-TCG-GTC-TTG-ATC-TCA-TGC-AAA-AAG-AAG-TAA-AAC-GTAT-TGG-TTT-CGG-TAA-TGC-TGA-AAT-TGG-ACA-GCA-GGT-TGA-TAA-TTT-C-3′ |
| GenBank accession number of the insert | OM310935.1 |
| Reference | [60] |
| PCR target | OXA-40/24-like-type beta-lactamase |
| Target gene | blaOXA-40/24-like |
| Detection limit | <102 copies/µL |
| Forward primer | 5′-TGA-CTT-TAG-GTG-AGG-CAA-TG-3′ |
| Reverse primer | 5′-GTT-ATG-TGC-AAG-GTC-ATC-GG-3′ |
| Probe and modifications | 5′-Cy5-TGC-AAG-ACG-GAC-TGG-CCT-AGA-GCT-AAT-BHQ2-3′ |
| Positive control plasmid insert | 5′-GAG-AAA-GAT-ATG-ACT-TTA-GGT-GAG-GCA-ATG-GCA-TTG-TCA-GCA-GTT-CCA-GTA-TAT-CAA-GAG-CTT-GCA-AGA-CGG-ACT-GGC-CTA-GAG-CTA-ATG-CAG-AAA-GAA-GTA-AAG-CGG-GTT-AAT-TTT-GGA-AAT-ACA-AAT-ATT-GGA-ACA-CAG-GTC-GAT-AAT-TTT-TGG-TTA-GTT-GGC-CCC-CTT-AAA-ATT-ACA-CCA-GTA-CAA-GAA-GTT-AAT-TTT-GCC-GAT-GAC-CTT-GCA-CAT-AAC-CGA-TTA-CCT-T-3′ |
| GenBank accession number of the insert | NG_078047.1 |
| Reference | [60] |
| PCR target | OXA-48-like-type beta-lactamase |
| Target gene | blaOXA-48-like |
| Detection limit | <102 copies/µL |
| Forward primer | 5′-AGG-GCG-TAG-TTG-TGC-TC-3′ |
| Reverse primer | 5′-GTG-TTC-ATC-CTT-AAC-CAC-GC-3′ |
| Probe and modifications | 5′-ROX-TCT-TAA-ACG-GGC-GAA-CCA-AGC-AT-BHQ2-3′ |
| Positive control plasmid insert | 5′-CAT-AAA-TCA-CAG-GGC-GTA-GTT-GTG-CTC-TGG-AAT-GAG-AAT-AAG-CAG-CAA-GGA-TTT-ACC-AAT-AAT-CTT-AAA-CGG-GCG-AAC-CAA-GCA-TTT-TTA-CCC-GCA-TCT-ACC-TTT-AAA-ATT-CCC-AAT-AGC-TTG-ATC-GCC-CTC-GAT-TTG-GGC-GTG-GTT-AAG-GAT-GAA-CAC-CAA-GTC-TTT-A-3′ |
| GenBank accession number of the insert | ON651448.1 |
| Reference | [60] |
| PCR target | OXA-58-like-type beta-lactamase |
| Target gene | blaOXA-58-like |
| Detection limit | <102 copies/µL |
| Forward primer | 5′-ATT-GGC-ACG-TCG-TAT-TGG-3′ |
| Reverse primer | 5′-CCC-CTC-TGC-GCT-CTA-CAT-A-3′ |
| Probe and modifications | 5′-JOE-AGT-GAA-TTG-CAA-CGT-ATT-GGT-TAT-GGC-A-BHQ1-3′ |
| Positive control plasmid insert | 5′-TAT-ATC-AAG-AAT-TGG-CAC-GTC-GTA-TTG-GTC-CAA-GCT-TAA-TGC-AAA-GTG-AAT-TGC-AAC-GTA-TTG-GTT-ATG-GCA-ATA-TGC-AAA-TAG-GCA-CGG-AAG-TTG-ATC-AAT-TTT-GGT-TGA-AAG-GGC-CTT-TGA-CAA-TTA-CAC-CTA-TAC-AAG-AAG-TAA-AGT-TTG-TGT-ATG-ATT-TAG-CCC-AAG-GGC-AAT-TGC-CTT-TTA-AAC-CTG-AAG-TTC-AGC-AAC-AAG-TGA-AAG-AGA-TGT-TGT-ATG-TAG-AGC-GCA-GAG-GGG-AGA-ATC-GTC-T-3′ |
| GenBank accession number of the insert | KY660721.1 |
| Reference | [60] |
| PCR target | GES-type beta-lactamase |
| Target gene | blaGES |
| Detection limit | <102 copies/µL |
| Forward primer | 5′-TGG-CTA-AAG-TCC-TCT-ATG-3′ |
| Reverse primer | 5′-CAA-CCC-AAT-CTT-TAG-GAA-A-3′ |
| Probe and modifications | 5′-FAM-CGT-CTC-CCG-TTT-GGT-TTC-CG-TAMRA-3′ |
| Positive control plasmid insert | 5′-GCA-CGT-ACT-GTG-GCT-AAA-GTC-CTC-TAT-GGC-GGC-GCA-CTG-ACG-TCC-ACC-TCG-ACC-CAC-ACC-ATT-GAG-AGG-TGG-CTG-ATC-GGA-AAC-CAA-ACG-GGA-GAC-GCG-ACA-CTA-CGA-GCG-GGT-TTT-CCT-AAA-GAT-TGG-GTT-GTT-GGA-GAG-AA-3′ |
| GenBank accession number of the insert | NG_080773.1 |
| Reference | [61] |
| PCR target | NMC-A/IMI-type beta-lactamase |
| Target gene | blaNMC-A/IMI |
| Detection limit | <102 copies/µL |
| Forward primer | 5′-GTC-ACT-TAA-TGT-AAA-ACC-AA-3′ |
| Reverse primer | 5′-CTA-CCA-TTG-AAA-TCT-GTT-TC-3′ |
| Probe and modifications | 5′-Cy5-AGC-CAT-CTT-GTT-TAG-CTC-TTG-TTT-AGT-BHQ2-3′ |
| Positive control plasmid insert | 5′-ATG-TCA-CTT-AAT-GTA-AAA-CCA-AGC-AGA-ATA-GCC-ATC-TTG-TTT-AGC-TCT-TGT-TTA-GTT-TCA-ATA-TCA-TTT-TTC-TCA-CAG-GCC-AAT-ACA-AAG-GGC-ATC-GAT-GAT-ATT-AAA-AAC-CTT-GAA-ACA-GAT-TTC-AAT-GGT-AGA-ATT-GGT-GTC-3′ |
| GenBank accession number of the insert | NG_065426.1 |
| Reference | [61] |
| PCR target | BIC-type beta-lactamase |
| Target gene | blaBIC |
| Detection limit | <102 copies/µL |
| Forward primer | 5′-GGA-GAA-ACG-TAT-CGA-CTA-TA-3′ |
| Reverse primer | 5′-TCC-AGA-AGC-AAA-TTT-GTC-3′ |
| Probe and modifications | 5′-JOE-CAC-CGT-TGT-CGC-TGT-ACT-GC-BHQ1-3′ |
| Positive control plasmid insert | 5′-AAG-GCT-TAC-TGG-AGA-AAC-GTA-TCG-ACT-ATA-AGA-ATC-GGG-TGA-TGG-AAC-CTC-ACT-CTC-CCA-TCA-GCG-CAC-AAC-ATA-GTT-CGA-CGG-GTA-TGA-CCG-TGG-CGC-AAT-TAG-CGG-CAG-CGG-CGC-TGC-AGT-ACA-GCG-ACA-ACG-GTG-CGA-CAA-ATT-TGC-TTC-TGG-AAA-ACG-TTC-TG-3′ |
| GenBank accession number of the insert | NG_048708.1 |
| Reference | [61] |
| PCR target | SME-type beta-lactamase |
| Target gene | blaSME |
| Detection limit | <102 copies/µL |
| Forward primer | 5′-GGC-TCA-GGT-ATG-ACA-TTA-3′ |
| Reverse primer | 5′-TCT-CCA-ATA-GAA-CGC-ATA-A-3′ |
| Probe and modifications | 5′-ROX-CTC-AGG-ACC-GCC-AAG-AAA-TCG-BHQ2-3′ |
| Positive control plasmid insert | 5′-AAA-ATA-TAA-AGG-CTC-AGG-TAT-GAC-ATT-AGG-TGA-TAT-GGC-TTC-TGC-TGC-ATT-GCA-ATA-TAG-CGA-CAA-TGG-GGC-AAC-AAA-TAT-AAT-TAT-GGA-ACG-ATT-TCT-TGG-CGG-TCC-TGA-GGG-GAT-GAC-TAA-ATT-TAT-GCG-TTC-TAT-TGG-AGA-TAA-TGA-GTT-T-3′ |
| GenBank accession number of the insert | MN182491.1 |
| Reference | [61] |
| PCR target | GIM-type beta-lactamase |
| Target gene | blaGIM |
| Detection limit | <102 copies/µL |
| Forward primer | 5′-TCG-ACA-CAC-CTT-GGT-CTG-AA-3′ |
| Reverse primer | 5′-AAC-TTC-CAA-CTT-TGC-CAT-GC-3′ |
| Probe and modifications | 5′-FAM-CAC-GAA-GTT-GTT-ATT-ATC-CTG-GGC-GAC-T-TAMRA-3′ |
| Positive control plasmid insert | 5′-GCC-TAT-ATT-ATC-GAC-ACA-CCT-TGG-TCT-GAA-GAA-GAC-ACG-AAG-TTG-TTA-TTA-TCC-TGG-GCG-ACT-GAC-AGG-GGA-TAC-CAG-GTT-ATG-GCT-AGC-ATC-TCA-ACT-CAT-TCT-CAT-GGA-GAT-CGC-ACT-GCT-GGT-ATC-AAG-TTG-CTA-AAT-TCA-AAG-TCA-ATT-CCT-ACA-TAC-ACA-TCA-GAG-TTA-ACT-AAA-AAG-CTT-CTT-GCC-CGT-GAA-GGA-AAG-CCG-GTT-CCT-ACC-CAC-TAC-TTT-AAA-GAC-GAC-GAA-TTC-ACA-CTG-GGA-AAT-GGG-CTT-ATA-GAG-CTC-TAC-TAT-CCA-GGT-GCT-GGG-CAT-ACA-GAG-GAT-AAT-ATT-GTT-GCT-TGG-TTA-CCC-AAA-AGC-AAA-ATA-CTA-TTT-GGT-GGC-TGC-CTC-GTG-AGG-AGT-CAT-GAG-TGG-GAA-GGC-TTA-GGT-TAC-GTA-GGC-GAC-GCC-TCA-ATT-AGC-TCT-TGG-GCT-GAC-TCA-ATT-AAA-AAT-ATT-GTA-TCG-AAA-AAA-TAT-CCC-ATT-CAA-ATG-GTC-GTT-CCG-GGG-CAT-GGC-AAA-GTT-GGA-AGT-TCA-GAT-ATA-TT-3′ |
| GenBank accession number of the insert | MK847892.1 |
| Reference | [62], probe from this study |
| PCR target | DIM-type beta-lactamase |
| Target gene | blaDIM |
| Detection limit | <102 copies/µL |
| Forward primer | 5′-GCT-TGT-CTT-CGC-TTG-CTA-ACG-3′ |
| Reverse primer | 5′-CGT-TCG-GCT-GGA-TTG-ATT-TG-3′ |
| Probe and modifications | 5′-Cy5-ACA-CAT-CAT-ACA-GTC-GTG-TGA-ATG-GGT-TTG-BHQ2-3′ |
| Positive control plasmid insert | 5′-CTT-CTA-TTC-AGC-TTG-TCT-TCG-CTT-GCT-AAC-GAC-GAG-GTA-CCT-GAG-CTA-AGA-ATC-GAG-AAA-GTA-AAA-GAG-AAC-ATC-TTT-TTG-CAC-ACA-TCA-TAC-AGT-CGT-GTG-AAT-GGG-TTT-GGT-TTG-GTC-AGT-TCA-AAC-GGC-CTT-GTT-GTC-ATA-GAT-AAG-GGT-AAT-GCT-TTC-ATT-GTT-GAT-ACA-CCT-TGG-TCA-GAC-CGA-GAT-ACA-GAA-ACG-CTC-GTA-CAT-TGG-ATT-CGT-AAA-AAT-GGT-TAT-GAG-CTA-CTG-GGG-AGT-GTT-TCT-ACT-CAT-TGG-CAT-GAG-GAT-AGA-ACC-GCA-GGA-ATT-AAA-TGG-CTT-AAT-GAC-CAA-TCA-ATT-TCT-ACG-TAT-GCC-ACG-ACT-TCA-ACC-AAC-CAT-CTC-TTG-AAA-GAA-AAT-AAA-AAA-GAG-CCA-GCG-AAA-TAC-ACC-TTG-AAA-GGA-AAT-GAG-TCC-ACA-TTG-GTT-GAC-GGC-CTT-ATC-GAA-GTA-TTT-TAT-CCA-GGA-GGT-GGT-CAT-ACA-ATA-GAC-AAC-GTA-GTG-GTG-TGG-TTG-CCA-AAG-TCG-AAA-ATC-TTA-TTT-GGC-GGC-TGT-TTT-GTG-CGT-AGC-CTT-GAT-TCC-GAG-GGG-TTA-GGC-TAC-ACT-GGT-GAA-GCC-CAT-ATT-GAT-CAA-TGG-TCC-CGA-TCA-GCT-CAG-AAT-GCT-CTG-TCT-AGG-TAC-TCA-GAA-GCC-CAG-ATA-GTA-ATT-CCT-GGC-CAT-GGG-AAA-ATC-GGG-GAT-ATA-GCG-CTG-TTA-AAA-CAC-ACC-AAA-AGT-CTG-GCT-GAG-ACA-GCC-TCT-AAC-AAA-TCA-ATC-CAG-CCG-AAC-GCT-AAC-GCG-TC-3′ |
| GenBank accession number of the insert | NG_049077.1 |
| Reference | [62], probe from this study |
| PCR target | Treponema pallidum |
| Target gene | polA |
| Detection limit | <102 copies/µL |
| Forward primer | 5′-AGG-ATC-CGG-CAT-ATG-TCC-AA-3′ |
| Reverse primer | 5′-GTG-AGC-GTC-TCA-TCA-TTC-CAA-A-3′ |
| Probe and modifications | 5′-FAM-ATG-CAC-CAG-CTT-CGA-MGB-NFQ-3′ |
| Positive control plasmid insert | 5′-TCT-GCT-GTG-CAG-GAT-CCG-GCA-TAT-GTC-CAA-GCT-GTC-ATG-CAC-CAG-CTT-CGA-CGT-CTT-TGG-AAT-GAT-GAG-ACG-CTC-ACA-CTT-GTT-ATG-3′ |
| GenBank accession number of the insert | U57757.1 |
| Reference | [63] |
| PCR target | Chlamydia trachomatis (screening) |
| Target gene | Chlamydia trachomatis cryptic plasmid sequence |
| Detection limit | <102 copies/µL |
| Forward primer | 5′-GGA-TTG-ACT-CCG-ACA-ACG-TAT-TC-3′ |
| Reverse primer | 5′-ATC-ATT-GCC-ATT-AGA-AAG-GGC-ATT-3′ |
| Probe and modifications | 5′-Cy5-TTA-CGT-GTA-GGC-GGT-TTA-GAA-AGC-GG-BHQ2-3′ |
| Positive control plasmid insert | 5′-TAC-TAA-TAC-AGG-ATT-GAC-TCC-GAC-AAC-GTA-TTC-ATT-ACG-TGT-AGG-CGG-TTT-AGA-AAG-CGG-TGT-GGT-ATG-GGT-TAA-TGC-CCT-TTC-TAA-TGG-CAA-TGA-TAT-TTT-AGG-AA-3′ |
| GenBank accession number of the insert | CP010570.1 |
| Reference | [64] |
| PCR target | Chlamydia trachomatis (differentiation) |
| Target gene | pmpH |
| Detection limit | <102 copies/µL |
| Forward primer | 5′-GGA-TAA-CTC-TGT-GGG-GTA-TTC-TCC-T-3′ |
| Reverse primer | 5′-AGA-CCC-TTT-CCG-AGC-ATC-ACT-3′ |
| Probe and modifications (pan-serovar) | 5′-FAM-CCT-GCT-CCA-ACA-GT-MGB-NFQ-3′ |
| Probe and modifications (A-K-serovars only) | 5′-ROX-GCT-TGA-AGC-AGC-AGG-AGC-TGG-TG-BHQ2-3′ |
| Positive control plasmid insert (pan-serovar) | 5′-TTG-ATT-TTC-TGG-GAT-AAC-TCC-GTG-GGG-TAT-TCT-CCT-TTA-TCT-ACT-GTG-CCA-ACC-TCA-TCA-TCA-ACT-CCG-CCT-GCT-CCA-ACA-GTT-AGT-GAT-GCT-CGG-AAA-GGG-TCT-ATT-TTT-TCT-G-3′ |
| GenBank accession number of the insert | AY184168.1 |
| Positive control plasmid insert (A-K-serovars only) | 5′-GTG-ATT-TTT-TGG-GAT-AAC-TCT-GTG-GGG-TAT-TCT-CCT-TTG-TCT-ATT-GTG-CCA-GCA-TCG-ACT-CCA-ACT-CCT-CCA-GCA-CCA-GCA-CCA-GCT-CCT-GCT-GCT-TCA-AGC-TCT-TTA-TCT-CCA-ACA-GTT-AGT-GAT-GCT-CGG-AAA-GGG-TCT-ATT-TTT-TCT-G-3′ |
| GenBank accession number of the insert | AY184158.1 |
| Reference | [64] |
| PCR target | Mycoplasma genitalium |
| Target gene | sequence of the MgPA operon |
| Detection limit | <102 copies/µL |
| Forward primer | 5′-GAG-AAA-TAC-CTT-GAT-GGT-CAG-CAA-3′ |
| Reverse primer | 5′-GTT-AAT-ATC-ATA-TAA-AGC-TCT-ACC-GTT-GTT-ATC-3′ |
| Probe and modifications | 5‘-ROX-AC*T-TT*G-CAA-*TC*A-*GAA-*GGT-BHQ2-3‘ |
| Positive control plasmid insert | 5′-CAA-TGC-TGT-TGA-GAA-ATA-CCT-TGA-TGG-TCA-GCA-AAA-CTT-TGC-AAT-CAG-AAG-GTA-TGA-TAA-CAA-CGG-TAG-AGC-TTT-ATA-TGA-TAT-TAA-CTT-AGC-AAA-AA-3′ |
| GenBank accession number of the insert | M31431.1 |
| Reference | [65,66] |
| PCR target | Neisseria gonorrhoeae (PCR 1 out of 2) |
| Target gene | multi-copy opa genes |
| Detection limit | <102 copies/µL |
| Forward primer | 5′-TTG-AAA-CAC-CGC-CCG-GAA-3′ |
| Reverse primer | 5′-TTT-CGG-CTC-CTT-ATT-CGG-TTT-AA-3′ |
| Probe and modifications | 5′-JOE-CCG-ATA-TAA-TC*C-GTC-*CTT-CAA-*CAT-CAG-BHQ1-3′ |
| Positive control plasmid insert | 5′-CCA-TAT-TGT-GTT-GAA-ACA-CCG-CCC-GGA-ACC-CGA-TAT-AAT-CCG-TCC-TTC-AAC-ATC-AGT-GAA-AAT-CTT-TTT-TTA-ACC-GGT-TAA-ACC-GAA-TAA-GGA-GCC-GAA-AAT-GAA-TCC-AG-3′ |
| GenBank accession number of the insert | X52372.1 |
| Reference | [67] |
| PCR target | Neisseria gonorrhoeae (PCR 2 out of 2) |
| Target gene | porA pseudogene |
| Detection limit | <102 copies/µL |
| Forward primer | 5′-CAG-CAT-TCA-ATT-TGT-TCC-GAG-TC-3′ |
| Reverse primer | 5′-GAA-CTG-GTT-TCA-TCT-GAT-TAC-TTT-CCA-3′ |
| Probe and modifications | 5′-Cy5-CGC-CTA-TAC-GCC-TGC-TAC-TTT-CAC-GC-BHQ2-3′ |
| Positive control plasmid insert | 5′-GTT-TCA-GCG-GCA-GCA-TTC-AAT-TTG-TTC-CGA-GTC-AAA-ACA-GCA-AGT-CCG-CCT-ATA-CGC-CTG-CTA-CTT-TCA-CGC-TGG-AAA-GTA-ATC-AGA-TGA-AAC-CAG-TTC-CGG-CTG-TTG-T-3′ |
| GenBank accession number of the insert | AJ010732.1 |
| Reference | [67] |
| PCR target | Schistosoma haematobium complex |
| Target gene | Dra1 multi-copy sequence |
| Detection limit | 5 × 102 copies/µL |
| Forward primer | 5′-GAT-CTC-ACC-TAT-CAG-ACG-AAA-C-3′ |
| Reverse primer | 5′-TCA-CAA-CGA-TAC-GAC-CAA-C-3′ |
| Probe and modifications | 5′-JOE-TGT-TGG-TGG-AAG-GCC-TGT-TTG-CAA-BHQ1-3′ |
| Positive control plasmid insert | 5‘-AAA-TTG-GAT-CTC-ACC-TAT-CAG-ACG-AAA-CAA-AGA-AAA-TTT-TAA-AAT-TGT-TGG-TGG-AAG-TGC-CTG-TTT-CGC-AAT-ATC-TCC-GGA-ATG-GTT-GGT-CGT-ATC-GTT-GTG-AAA-ATT-G-3‘ |
| GenBank accession number of the insert | DQ157698.1 |
| Reference | [68] |
| PCR target | Schistosoma mansoni complex |
| Target gene | Sm1-7 multi-copy sequence |
| Detection limit | 5 × 102 copies/µL |
| Forward primer | 5′-CCA-CGC-TCT-CGC-AAA-TAA-TCT-3′ |
| Reverse primer | 5′-CAA-CCG-TTC-TAT-GAA-AAT-CGT-TGT-3′ |
| Probe and modifications | 5′-FAM-TCC-GAA-ACC-ACT-GGA-CGG-ATT-TTT-ATG-AT-BHQ1-3′ |
| Positive control plasmid insert | 5‘-TCC-GAC-CAA-CCG-TTC-TAT-GAA-AAT-CGT-TGT-ATC-TCC-GAA-ACC-ACT-GGA-CGG-ATT-TTT-ATG-ATG-TTT-GTT-TTA-GAT-TAT-TTG-CGA-GAG-CGT-GGG-CGT-TA-3‘ |
| GenBank accession number of the insert | M61098.1 |
| Reference | [68] |
| blaCTX-M Gene-Specific Assay | blaVIM, blaIMP, blaNDM and blaKPC Gene-Specific Assay | blaOXA-23-like, blaOXA-40/24-like, blaOXA-48-like and blaOXA-58-like Gene-Specific Assay | |
|---|---|---|---|
| Reaction chemistry | |||
| Master Mix | HotStarTaq (Qiagen) | HotStarTaq (Qiagen) | HotStarTaq (Qiagen) |
| Reaction volume (µL) | 20.0 | 20.0 | 20.0 |
| Forward primer concentration (nM) | 320.0 (each) | 750.0 (each) | 800.0 (OXA-48-like & OXA-40/24-like), 600.0 (OXA-58-like), 400.0 (OXA-48-like) |
| Reverse primer concentration (nM) | 320.0 (each) | 375 (IMP reverse primers I & II), 750 (all others) | 800.0 (OXA-48-like & OXA-40/24-like), 600.0 (OXA-58-like), 400.0 (OXA-48-like) |
| Probe concentration (nM) | 160.0 | 188.0 (VIM), 200.0 (both IMP probes), 250 (NDM & KPC) | 400.0 (OXA-48-like & OXA-40/24-like), 250.0 (OXA-58-like), 150.0 (OXA-48-like) |
| Final Mg2+ concentration (nM) | 3.0 | 3.0 | 6.0 |
| Bovine serum albumin (ng/µL) | 2.0 | 2.0 | 2.0 |
| Run conditions | |||
| Initial denaturation | 95 °C, 15 min | 95 °C, 15 min | 95 °C, 15 min |
| Cycle numbers | 40 | 40 | 40 |
| Denaturation | 95 °C, 15 s | 95 °C, 15 s | 95 °C, 15 s |
| Annealing | Combined with amplification | Combined with amplification | Combined with amplification |
| Amplification | 60 °C, 60 s | 60 °C, 60 s | 60 °C, 60 s |
| Hold | 40 °C, 20 s | 40 °C, 20 s | 40 °C, 20 s |
| blaGES, blaNMC_A/IMI, blaBIC and blaSME Gene-Specific Assay | blaGIM Gene-Specific Assay | blaDIM Gene-Specific Assay | |
|---|---|---|---|
| Reaction chemistry | |||
| Master Mix | HotStarTaq (Qiagen) | HotStarTaq (Qiagen) | HotStarTaq (Qiagen) |
| Reaction volume (µL) | 20.0 | 20.0 | 20.0 |
| Forward primer concentration (nM) | 750.0 (each) | 400 | 400 |
| Reverse primer concentration (nM) | 750.0 (each) | 400 | 400 |
| Probe concentration (nM) | 375 (NMC_A/IMI), 188 (all others) | 200 | 250 |
| Final Mg2+ concentration (nM) | 3.0 | 6.0 | 6.0 |
| Bovine serum albumin (ng/µL) | 2.0 | 2.0 | 2.0 |
| Run conditions | |||
| Initial denaturation | 95 °C, 15 min | 95 °C, 15 min | 95 °C, 15 min |
| Cycle numbers | 40 | 40 | 40 |
| Denaturation | 95 °C, 15 s | 95 °C, 30 s | 95 °C, 30 s |
| Annealing | Combined with amplification | 46 °C, 40 s | 46 °C, 40 s |
| Amplification | 60 °C, 60 s | 72 °C, 50 s | 72 °C, 50 s |
| Hold | 40 °C, 20 s | 40 °C, 20 s | 40 °C, 20 s |
| Chlamydia trachomatis Screening and Differentiation Assay | Neisseria gonorrhoeae, Mycoplasma genitalium and Treponema pallidum Assay | Schistosoma haematobium Complex and Schistosoma mansoni Complex Assay | |
|---|---|---|---|
| Reaction chemistry | |||
| Master Mix | HotStarTaq (Qiagen) | HotStarTaq (Qiagen) | HotStarTaq (Qiagen) |
| Reaction volume (µL) | 20.0 | 20.0 | 20.0 |
| Forward primer concentration (nM) | 50.0 (screening), 600.0 (differentiation) | 900.0 (T. pallidum), 1000.0 (M. genitalium), 400.0 (gonococci, both assays) | 500.0 (each) |
| Reverse primer concentration (nM) | 100.0 (screening), 600.0 (differentiation) | 900.0 (T. pallidum), 1000.0 (M. genitalium), 400.0 (gonococci, both assays) | 500.0 (each) |
| Probe concentration (nM) | 100.0 (screening), 200.0 (differentiation) | 250.0 (T. pallidum), 225.0 (M. genitalium), 160.0 (gonococci, both assays) | 300.0 (each) |
| Final Mg2+ concentration (nM) | 4.0 | 5.0 | 6.0 |
| Bovine serum albumin (ng/µL) | 2.0 | 2.0 | 2.0 |
| Run conditions | |||
| Initial denaturation | 95 °C, 15 min. | 95 °C, 15 min. | 95 °C, 15 min. |
| Cycle numbers | 45 | 50 | 40 |
| Denaturation | 95 °C, 15 sec. | 95 °C, 15 sec. | 95 °C, 15 sec. |
| Annealing | Combined with amplification | Combined with amplification | Combined with amplification |
| Amplification | 60 °C, 60 sec. | 60 °C, 60 sec. | 65 °C, 60 sec. |
| Hold | 40 °C, 20 sec. | 40 °C, 20 sec. | 40 °C, 10 sec. |
| Bacterial Resistance Determinants | Sexually Transmitted Infections | Schistosomiasis | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Mean (±SD) | Median (Min., Max.) | Significance P | Mean (±SD) | Median (Min., Max.) | Significance P | Mean (±SD) | Median (Min., Max.) | Significance P | |
| Age in years (+) | 28.0 (±5.7) | 28 (18, 46) | 0.31 | 25.8 (±5.3) | 25 (18, 33) | 0.18 | 29.1 (±5.6) | 29 (18, 46) | 0.37 |
| Age in years (-) | 28.6 (±5.8) | 28.5 (18, 43) | 28.5 (±5.8) | 28 (18, 46) | 28.3 (±5.8) | 28 (18, 44) | |||
| Number of pregnancies (+) | 3.2 (±1.9) | 3 (1, 9) | 0.70 | 3.5 (±3.0) | 2 (1, 9) | 1.00 | 3.2 (±1.9) | 3 (1, 8) | 0.57 |
| Number of pregnancies (-) | 3.1 (±2.0) | 3 (1, 9) | 3.1 (±1.7) | 3 (1, 9) | 3.1 (±2.0) | 3 (1, 9) | |||
| APGAR 1 (+) | 8.0 (±0.9) | 8 (6, 10) | 0.17 | 8.1 (±1.1) | 8 (6, 9) | 0.46 | 7.5 (±1.1) | 8 (5, 10) | 0.01 |
| APGAR 1 (-) | 7.8 (±1.1) | 8 (2, 10) | 7.9 (±1.1) | 8 (2, 9) | 7.9 (±1.1) | 8 (2, 10) | |||
| APGAR 2 (+) | 8.8 (±0.5) | 9 (7, 10) | 0.37 | 9.1 (±0.3) | 9 (9, 10) | 0.12 | 8.6 (±0.7) | 9 (7, 10) | 0.08 |
| APGAR 2 (-) | 8.7 (±0.7) | 9 (4, 10) | 8.7 (±0.7) | 9 (4, 10) | 8.8 (±0.6) | 9 (4, 10) | |||
| Bacterial Resistance Determinants | Sexually Transmitted Infections | Schistosomiasis | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Proportion of Samples Positive for Bacterial Resistance Determinants in % (n/n) | Odds Ratio (95%-CI) | Significance P | Proportion of Samples Positive for Sexually Transmitted Infections in % (n/n) | Odds Ratio (95%-CI) | Significance P | Proportion of Samples Positive for Schistosomiasis in % (n/n) | Odds Ratio (95%-CI) | Significance P | |
| Breeding of chicken | 18.4% (7/38) | 0.45 (0.19, 1.07) | 0.09 | 2.6% (1/38) | 0.68 (0.08, 5.54) | 1.00 | 15.8% (6/38) | 0.80 (0.31, 2.02) | 0.82 |
| No breeding of chicken | 33.4% (79/236) | 3.8% (9/236) | 19.7% (45/236) | ||||||
| Possession of a freezer | 25.3% (25/99) | 0.63 (0.36, 1.09) | 0.11 | 2.0% (2/99) | 0.43 (0.09, 2.07) | 0.34 | 33.3% (25/99) | 4.36 (2.29, 8.29) | <0.0001 |
| No possession of a freezer | 34.9% (61/175) | 4.6% (8/175) | 10.3% (18/175) | ||||||
| Electricity at home | 32.9% (86/269) | 5.19 (0.28, 94.90) | 0.33 | 3.7% (10/269) | 0.45 (0.02, 8.60) | 1.00 | 19.0% (51/269) | 2.59 (0.14, 47.67) | 0.59 |
| No electricity at home (-) | 0.0% (0/5) | 0.0% (0/5) | 0.0% (0/5) | ||||||
| Delivery via Caesarian section | 19.0% (4/21) | 0.50 (0.16, 1.54) | 0.33 | 0.0% (0/21) | 0.58 (0.03, 10.26) | 1.00 | 61.9% (13/21) | 9.39 (3.63, 24.26) | <0.0001 |
| Vaginal delivery | 32.0% (78/244) | 3.7% (9/244) | 14.8% (36/244) | ||||||
| Country: Ivory Coast | 33.6% (75/223) | 1.84 (0.89, 3.80) | 0.13 | 4.0% (9/223) | 2.10 (0.26, 16.99) | 0.69 | 10.3% (23/223) | 0.094 (0.047, 0.190) | <0.0001 |
| Country: Ghana | 21.6% (11/51) | 2.0% (1/51) | 54.9% (28/51) | ||||||
| Bacterial Resistance Determinants | Sexually Transmitted Infections | Schistosomiasis | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Proportion of Samples Positive for Bacterial Resistance Determinants in % (n/n) | Chi-Square Value | Significance P | Proportion of Samples Positive for Sexually Transmitted Infections in % (n/n) | Chi-Square Value | Significance P | Proportion of Samples Positive for Schistosomiasis in % (n/n) | Chi-Square Value | Significance P | |
| Floor at home: earth/sand | 26.7% (4/15) | 0.24 | 0.89 | 0.0% (0/15) | 1.21 | 0.55 | 2.0% (3/15) | 4.56 | 0.10 |
| Floor at home: vinyl/tiles | 30.0% (15/50) | 2.0% (1/50) | 8.0% (4/50) | ||||||
| Floor at home: cement | 32.1% (67/209) | 4.3%% (9/209) | 21.1% (44/209) | ||||||
| Toilet: pit latrine | 36.5% (54/148) | 4.21 | 0.12 | 5.4% (8/148) | 3.48 | 0.18 | 12.8% (19/148) | 7.15 | 0.03 |
| Toilet: improved pit latrine | 27.4% (20/73) | 2.7% (2/73) | 24.7% (18/73) | ||||||
| Toilet: flush toilet | 22.6% (12/53) | 0% (0/53) | 26.4% (12/53) | ||||||
| Water source: tanker | 0.0% (0/1) | 2.74 | 0.43 | 0.0% (0/1) | 0.93 | 0.82 | 0.0% (0/1) | 7.09 | 0.07 |
| Water source: well | 22.2% (2/9) | 0.0% (0/9) | 44.4% (4/9) | ||||||
| Water source: piped water | 32.2% (84/260) | 3.8% (10/260) | 17.3% (45/260) | ||||||
| Water source: other (also not surface water or bore hole) | 0.0% (0/4) | 0.0% (0/4) | 50.0% (2/4) | ||||||
| Occupation: housewife | 31.0% (18/58) | 4.24 | 0.38 | 3.4% (2/58) | 0.10 | 1.00 | 13.8% (8/58) | 3.64 | 0.46 |
| Occupation: farmer | 100% (1/1) | 0.0% (0/1) | 0.0% (0/1) | ||||||
| Occupation: trader | 32.2% (19/59) | 3.4% (2/59 | 13.6% (8/59) | ||||||
| Occupation: salary worker | 20.0% (6/30) | 3.3% (1/30) | 23.3% (7/30) | ||||||
| Occupation: other | 33.3% (42/126) | 4.0% (5/126) | 22.2% (28/126) | ||||||
| Education: none | 36.7% (40/109) | 3.60 | 0.31 | 2.8% (3/109) | 2.59 | 0.46 | 15.6% (17/109) | 2.43 | 0.49 |
| Education: primary | 25.9% (21/81) | 3.7% (3/81) | 23.5% (19/81) | ||||||
| Education: secondary | 32.8% (20/61) | 6.6% (4/61) | 19.7% (12/61) | ||||||
| Education: tertiary | 21.7% (5/23) | 0.0% (0/23) | 13.0% (3/23) | ||||||
References
- Ganyaglo, G.Y.; Hill, W.C. A 6-year (2004–2009) review of maternal mortality at the Eastern Regional Hospital, Koforidua, Ghana. Semin. Perinatol. 2012, 36, 79–83. [Google Scholar] [CrossRef]
- Lee, Q.Y.; Odoi, A.T.; Opare-Addo, H.; Dassah, E.T. Maternal mortality in Ghana: A hospital-based review. Acta Obstet. Gynecol. Scand. 2012, 91, 87–92. [Google Scholar] [CrossRef]
- Wiredu, E.K.; Tettey, Y. Autopsy studies on still births in Korle Bu Teaching Hospital. II: Causes of death in 93 still births. West Afr. J. Med. 1998, 17, 148–152. [Google Scholar]
- Tettey, Y.; Wiredu, E.K. Autopsy studies on still births in Korle Bu Teaching Hospital: Pathological findings in still births and their placentae. West Afr. J. Med. 1997, 16, 12–19. [Google Scholar]
- Ampofo, G.D.; Osarfo, J.; Aberese-Ako, M.; Asem, L.; Komey, M.N.; Mohammed, W.; Ofosu, A.A.; Tagbor, H. Malaria in pregnancy control and pregnancy outcomes: A decade’s overview using Ghana’s DHIMS II data. Malar. J. 2022, 21, 303. [Google Scholar] [CrossRef]
- Hommerich, L.; von Oertzen, C.; Bedu-Addo, G.; Holmberg, V.; Acquah, P.A.; Eggelte, T.A.; Bienzle, U.; Mockenhaupt, F.P. Decline of placental malaria in southern Ghana after the implementation of intermittent preventive treatment in pregnancy. Malar. J. 2007, 6, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osarfo, J.; Ampofo, G.D.; Tagbor, H. Trends of malaria infection in pregnancy in Ghana over the past two decades: A review. Malar. J. 2022, 21, 3. [Google Scholar] [CrossRef] [PubMed]
- Mwin, P.K.; Kuffuor, A.; Nuhu, K.; Okine, R.; Kubio, C.; Wurapa, F.; Osei, F.A.; Afari, E. Predictors of placental malaria in Upper West Regional Hospital-Ghana. BMC Pregnancy Childbirth 2021, 21, 403. [Google Scholar] [CrossRef]
- Akinnawo, A.; Seyram, K.; Kaali, E.B.; Harrison, S.; Dosoo, D.; Cairns, M.; Asante, K.P. Assessing the relationship between gravidity and placental malaria among pregnant women in a high transmission area in Ghana. Malar. J. 2022, 21, 240. [Google Scholar] [CrossRef] [PubMed]
- Tadesse Boltena, M.; El-Khatib, Z.; Kebede, A.S.; Asamoah, B.O.; Yaw, A.S.C.; Kamara, K.; Constant Assogba, P.; Tadesse Boltena, A.; Adane, H.T.; Hailemeskel, E.; et al. Malaria and Helminthic Co-Infection during Pregnancy in Sub-Saharan Africa: A Systematic Review and Meta-Analysis. Int. J. Environ. Res. Public. Health 2022, 19, 5444. [Google Scholar] [CrossRef]
- Brinkmann, U.K.; Krämer, P.; Presthus, G.T.; Sawadogo, B. Transmission in utero of microfilariae of Onchocerca volvulus. Bull. World Health Organ. 1976, 54, 708–709. [Google Scholar] [PubMed]
- Kwofie, K.D.; Ghansah, A.; Osei, J.H.; Frempong, K.K.; Obed, S.; Frimpong, E.H.; Boakye, D.A.; Suzuki, T.; Ohta, N.; Ayi, I. Indication of Risk of Mother-to-Child Toxoplasma gondii Transmission in the Greater Accra Region of Ghana. Matern. Child. Health J. 2016, 20, 2581–2588. [Google Scholar]
- Asmah, R.H.; Blankson, H.N.A.; Seanefu, K.A.; Obeng-Nkrumah, N.; Awuah-Mensah, G.; Cham, M.; Ayeh-Kumi, P.F. Trichomoniasis and associated co-infections of the genital tract among pregnant women presenting at two hospitals in Ghana. BMC Womens Health 2017, 17, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dako-Gyeke, P.; Dornoo, B.; Ayisi Addo, S.; Atuahene, M.; Addo, N.A.; Yawson, A.E. Towards elimination of mother-to-child transmission of HIV in Ghana: An analysis of national programme data. Int. J. Equity Health 2016, 15, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanDeusen, A.; Paintsil, E.; Agyarko-Poku, T.; Long, E.F. Cost effectiveness of option B plus for prevention of mother-to-child transmission of HIV in resource-limited countries: Evidence from Kumasi, Ghana. BMC Infect. Dis. 2015, 15, 130. [Google Scholar] [CrossRef] [Green Version]
- Jacquemyn, Y.; Van Casteren, C.; Luijks, M.; Colpaert, C. Disseminated tuberculosis in pregnancy unknown to doctors in Western Europe case presentation: ‘part of the routine study in infertility’. BMJ Case Rep. 2012, 2012, bcr2012006227. [Google Scholar] [CrossRef] [Green Version]
- Koné, S.; Hürlimann, E.; Baikoro, N.; Dao, D.; Bonfoh, B.; N’Goran, E.K.; Utzinger, J.; Jaeger, F.N. Pregnancy-related morbidity and risk factors for fatal foetal outcomes in the Taabo health and demographic surveillance system, Côte d’Ivoire. BMC Pregnancy Childbirth 2018, 18, 216. [Google Scholar] [CrossRef]
- Mbouamboua, Y.; Koukouikila-Koussounda, F.; Ntoumi, F.; Adukpo, S.; Kombo, M.; Vouvoungui, C.; van Helden, J.; Kobawila, S.C. Sub-microscopic Plasmodium falciparum infections in matched peripheral, placental and umbilical cord blood samples from asymptomatic Congolese women at delivery. Acta Trop. 2019, 193, 142–147. [Google Scholar] [CrossRef]
- Doritchamou, J.Y.A.; Akuffo, R.A.; Moussiliou, A.; Luty, A.J.F.; Massougbodji, A.; Deloron, P.; Tuikue Ndam, N.G. Submicroscopic placental infection by non-falciparum Plasmodium spp. PLoS Negl. Trop. Dis. 2018, 12, e0006279. [Google Scholar] [CrossRef] [Green Version]
- Fried, M.; Muehlenbachs, A.; Duffy, P.E. Diagnosing malaria in pregnancy: An update. Expert. Rev. Anti Infect. Ther. 2012, 10, 1177–1187. [Google Scholar] [CrossRef]
- Kattenberg, J.H.; Ochodo, E.A.; Boer, K.R.; Schallig, H.D.; Mens, P.F.; Leeflang, M.M. Systematic review and meta-analysis: Rapid diagnostic tests versus placental histology, microscopy and PCR for malaria in pregnant women. Malar. J. 2011, 10, 321. [Google Scholar] [CrossRef] [PubMed]
- Sprong, K.E.; Mabenge, M.; Wright, C.A.; Govender, S. Ureaplasma species and preterm birth: Current perspectives. Crit. Rev. Microbiol. 2020, 46, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Reagan-Steiner, S.; Bhatnagar, J.; Martines, R.B.; Milligan, N.S.; Gisondo, C.; Williams, F.B.; Lee, E.; Estetter, L.; Bullock, H.; Goldsmith, C.S.; et al. Detection of SARS-CoV-2 in Neonatal Autopsy Tissues and Placenta. Emerg. Infect. Dis. 2022, 28, 510–517. [Google Scholar] [CrossRef]
- Orish, V.N.; Morhe, E.K.S.; Azanu, W.; Alhassan, R.K.; Gyapong, M. The parasitology of female genital schistosomiasis. Curr. Res. Parasitol. Vector Borne Dis. 2022, 2, 100093. [Google Scholar] [CrossRef]
- Bustinduy, A.L.; Randriansolo, B.; Sturt, A.S.; Kayuni, S.A.; Leustcher, P.D.C.; Webster, B.L.; Van Lieshout, L.; Stothard, J.R.; Feldmeier, H.; Gyapong, M. An update on female and male genital schistosomiasis and a call to integrate efforts to escalate diagnosis, treatment and awareness in endemic and non-endemic settings: The time is now. Adv. Parasitol. 2022, 115, 1–44. [Google Scholar]
- Nyarko, R.; Torpey, K.; Ankomah, A. Schistosoma haematobium, Plasmodium falciparum infection and anaemia in children in Accra, Ghana. Trop. Dis. Travel. Med. Vaccines 2018, 4, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, T.; Gravett, C.A.; Velu, P.P.; Theodoratou, E.; Wagner, T.A.; Zhang, J.S.; Campbell, H.; Rubens, C.E.; Gravett, M.G.; Rudan, I. Epidemiology and aetiology of maternal parasitic infections in low- and middle-income countries. J. Glob. Health 2011, 1, 189–200. [Google Scholar] [PubMed]
- Angora, E.K.; Vangraefschepe, A.; Allienne, J.F.; Menan, H.; Coulibaly, J.T.; Meïté, A.; Raso, G.; Winkler, M.S.; Yavo, W.; Touré, A.O.; et al. Population genetic structure of Schistosoma haematobium and Schistosoma haematobium × Schistosoma bovis hybrids among school-aged children in Côte d’Ivoire. Parasite 2022, 29, 23. [Google Scholar] [CrossRef]
- Bassa, F.K.; Eze, I.C.; Assaré, R.K.; Essé, C.; Koné, S.; Acka, F.; Laubhouet-Koffi, V.; Kouassi, D.; Bonfoh, B.; Utzinger, J.; et al. Prevalence of Schistosoma mono- and co-infections with multiple common parasites and associated risk factors and morbidity profile among adults in the Taabo health and demographic surveillance system, South-Central Côte d’Ivoire. Infect. Dis. Poverty 2022, 11, 3. [Google Scholar] [CrossRef]
- Assaré, R.K.; N’Tamon, R.N.; Bellai, L.G.; Koffi, J.A.; Mathieu, T.I.; Ouattara, M.; Hürlimann, E.; Coulibaly, J.T.; Diabaté, S.; N’Goran, E.K.; et al. Characteristics of persistent hotspots of Schistosoma mansoni in western Côte d’Ivoire. Parasit. Vectors 2020, 13, 337. [Google Scholar] [CrossRef]
- Angora, E.K.; Allienne, J.F.; Rey, O.; Menan, H.; Touré, A.O.; Coulibaly, J.T.; Raso, G.; Yavo, W.; N’Goran, E.K.; Utzinger, J.; et al. High prevalence of Schistosoma haematobium × Schistosoma bovis hybrids in schoolchildren in Côte d’Ivoire. Parasitology 2020, 147, 287–294. [Google Scholar] [CrossRef] [PubMed]
- M’Bra, R.K.; Kone, B.; Yapi, Y.G.; Silué, K.D.; Sy, I.; Vienneau, D.; Soro, N.; Cissé, G.; Utzinger, J. Risk factors for schistosomiasis in an urban area in northern Côte d’Ivoire. Infect. Dis. Poverty 2018, 7, 47. [Google Scholar] [CrossRef] [Green Version]
- Assaré, R.K.; Lai, Y.S.; Yapi, A.; Tian-Bi, Y.N.; Ouattara, M.; Yao, P.K.; Knopp, S.; Vounatsou, P.; Utzinger, J.; N’Goran, E.K. The spatial distribution of Schistosoma mansoni infection in four regions of western Côte d’Ivoire. Geospat. Health 2015, 10, 345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coulibaly, J.T.; N’Gbesso, Y.K.; N’Guessan, N.A.; Winkler, M.S.; Utzinger, J.; N’Goran, E.K. Epidemiology of schistosomiasis in two high-risk communities of south Cote d’Ivoire with particular emphasis on pre-school-aged children. Am. J. Trop. Med. Hyg. 2013, 89, 32–41. [Google Scholar] [CrossRef]
- Matthys, B.; Tschannen, A.B.; Tian-Bi, N.T.; Comoé, H.; Diabaté, S.; Traoré, M.; Vounatsou, P.; Raso, G.; Gosoniu, L.; Tanner, M.; et al. Risk factors for Schistosoma mansoni and hookworm in urban farming communities in western Côte d’Ivoire. Trop. Med. Int. Health 2007, 12, 709–723. [Google Scholar] [CrossRef]
- Utzinger, J.; Müller, I.; Vounatsou, P.; Singer, B.H.; N’Goran, E.K.; Tanner, M. Random spatial distribution of Schistosoma mansoni and hookworm infections among school children within a single village. J. Parasitol. 2003, 89, 686–692. [Google Scholar] [CrossRef]
- Ahenkorah, B.; Nsiah, K.; Baffoe, P.; Anto, E.O. Biochemical and hematological changes among anemic and non-anemic pregnant women attending antenatal clinic at the Bolgatanga regional hospital, Ghana. BMC Hematol. 2018, 18, 27. [Google Scholar] [CrossRef] [Green Version]
- Donkor, E.S.; Dayie, N.T.K.D.; Tette, E.M.A. Methicillin-Resistant Staphylococcus aureus in Ghana: Past, Present, and Future. Microb. Drug Resist. 2019, 25, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Asamoah, B.; Labi, A.K.; Gupte, H.A.; Davtyan, H.; Peprah, G.M.; Adu-Gyan, F.; Nair, D.; Muradyan, K.; Jessani, N.S.; Sekyere-Nyantakyi, P. High Resistance to Antibiotics Recommended in Standard Treatment Guidelines in Ghana: A Cross-Sectional Study of Antimicrobial Resistance Patterns in Patients with Urinary Tract Infections between 2017–2021. Int. J. Environ. Res. Public Health 2022, 19, 16556. [Google Scholar] [CrossRef]
- Dekker, D.; Krumkamp, R.; Eibach, D.; Sarpong, N.; Boahen, K.G.; Frimpong, M.; Fechtner, E.; Poppert, S.; Hagen, R.M.; Schwarz, N.G.; et al. Characterization of Salmonella enterica from invasive bloodstream infections and water sources in rural Ghana. BMC Infect. Dis. 2018, 18, 47. [Google Scholar] [CrossRef]
- Müller-Schulte, E.; Tuo, M.N.; Akoua-Koffi, C.; Schaumburg, F.; Becker, S.L. High prevalence of ESBL-producing Klebsiella pneumoniae in clinical samples from central Côte d’Ivoire. Int. J. Infect. Dis. 2020, 91, 207–209. [Google Scholar] [CrossRef] [Green Version]
- Jeannot, K.; Guessennd, N.; Fournier, D.; Müller, E.; Gbonon, V.; Plésiat, P. Outbreak of metallo-β-lactamase VIM-2-positive strains of Pseudomonas aeruginosa in the Ivory Coast. J. Antimicrob. Chemother. 2013, 68, 2952–2954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labi, A.K.; Obeng-Nkrumah, N.; Bjerrum, S.; Enweronu-Laryea, C.; Newman, M.J. Neonatal bloodstream infections in a Ghanaian Tertiary Hospital: Are the current antibiotic recommendations adequate? BMC Infect. Dis. 2016, 16, 598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aku, F.Y.; Akweongo, P.; Nyarko, K.; Sackey, S.; Wurapa, F.; Afari, E.A.; Ameme, D.K.; Kenu, E. Bacteriological profile and antibiotic susceptibility pattern of common isolates of neonatal sepsis, Ho Municipality, Ghana-2016. Matern. Health Neonatol. Perinatol. 2018, 4, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addy, P.A. Susceptibility pattern of Neisseria gonorrhoeae isolated at the Komfo, Anokye Teaching Hospital, Ghana to commonly prescribed antimicrobial agents. East. Afr. Med. J. 1994, 71, 368–372. [Google Scholar] [PubMed]
- Wondergem, P.; Green, K.; Wambugu, S.; Asamoah-Adu, C.; Clement, N.F.; Amenyah, R.; Atuahene, K.; Szpir, M. A short history of HIV prevention programs for female sex workers in Ghana: Lessons learned over 3 decades. J. Acquir. Immune Defic. Syndr. 2015, 68 (Suppl. 2), S138–S145. [Google Scholar] [CrossRef]
- Cassels, S.; Jenness, S.M.; Biney, A.A.E. Coital Frequency and Male Concurrent Partnerships During Pregnancy and Postpartum in Agbogbloshie, Ghana. AIDS Behav. 2019, 23, 1508–1517. [Google Scholar] [CrossRef]
- Gakoué, D.Z.; Tiembré, I. Epidemiological aspects of genital ulcers at the STI center of the National Institute of Public Hygiene (Abidjan, Côte d’Ivoire) 2008 to 2010. Med. Sante Trop. 2017, 27, 90–94. [Google Scholar] [CrossRef]
- Vuylsteke, B.; Semdé, G.; Sika, L.; Crucitti, T.; Ettiègne Traoré, V.; Buvé, A.; Laga, M. HIV and STI prevalence among female sex workers in Côte d’Ivoire: Why targeted prevention programs should be continued and strengthened. PLoS ONE 2012, 7, e32627. [Google Scholar] [CrossRef]
- Vuylsteke, B.; Semde, G.; Sika, L.; Crucitti, T.; Ettiegne Traore, V.; Buve, A.; Laga, M. High prevalence of HIV and sexually transmitted infections among male sex workers in Abidjan, Cote d’Ivoire: Need for services tailored to their needs. Sex. Transm. Infect. 2012, 88, 288–293. [Google Scholar] [CrossRef] [Green Version]
- Vuylsteke, B.; Traore, M.; Mah-Bi, G.; Konan, Y.; Ghys, P.; Diarra, J.; Laga, M. Quality of sexually transmitted infections services for female sex workers in Abidjan, Côte d’Ivoire. Trop. Med. Int. Health 2004, 9, 638–643. [Google Scholar] [CrossRef] [PubMed]
- Vuylsteke, B.L.; Ettiègne-Traore, V.; Anoma, C.K.; Bandama, C.; Ghys, P.D.; Maurice, C.E.; Van Dyck, E.; Wiktor, S.Z.; Laga, M. Assessment of the validity of and adherence to sexually transmitted infection algorithms at a female sex worker clinic in Abidjan, Côte d’Ivoire. Sex. Transm. Dis. 2003, 30, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Vuylsteke, B.; Ghys, P.D.; Mah-bi, G.; Konan, Y.; Traoré, M.; Wiktor, S.Z.; Laga, M. Where do sex workers go for health care? A community based study in Abidjan, Côte d’Ivoire. Sex. Transm. Infect. 2001, 77, 351–352. [Google Scholar] [CrossRef] [Green Version]
- Lafort, Y.; Sawadogo, Y.; Delvaux, T.; Vuylsteke, B.; Laga, M. Should family planning clinics provide clinical services for sexually transmitted infections? A case study from Côte d’Ivoire. Trop. Med. Int. Health 2003, 8, 552–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bindt, C.; Appiah-Poku, J.; Te Bonle, M.; Schoppen, S.; Feldt, T.; Barkmann, C.; Koffi, M.; Baum, J.; Nguah, S.B.; Tagbor, H.; et al. Antepartum depression and anxiety associated with disability in African women: Cross-sectional results from the CDS study in Ghana and Côte d’Ivoire. PLoS ONE 2012, 7, e48396. [Google Scholar] [CrossRef] [Green Version]
- Bindt, C.; Guo, N.; Bonle, M.T.; Appiah-Poku, J.; Hinz, R.; Barthel, D.; Schoppen, S.; Feldt, T.; Barkmann, C.; Koffi, M.; et al. No association between antenatal common mental disorders in low-obstetric risk women and adverse birth outcomes in their offspring: Results from the CDS study in Ghana and Côte D’Ivoire. PLoS ONE 2013, 8, e80711. [Google Scholar] [CrossRef]
- Lina, G.; Piémont, Y.; Godail-Gamot, F.; Bes, M.; Peter, M.O.; Gauduchon, V.; Vandenesch, F.; Etienne, J. Involvement of Panton-Valentine leukocidin-producing Staphylococcus aureus in primary skin infections and pneumonia. Clin. Infect. Dis. 1999, 29, 1128–1132. [Google Scholar] [CrossRef] [Green Version]
- Harmsen, D.; Claus, H.; Witte, W.; Rothgänger, J.; Claus, H.; Turnwald, D.; Vogel, U. Typing of methicillin-resistant Staphylococcus aureus in a university hospital setting by using novel software for spa repeat determination and database management. J. Clin. Microbiol. 2003, 41, 5442–5448. [Google Scholar] [CrossRef] [Green Version]
- van der Zee, A.; Roorda, L.; Bosman, G.; Fluit, A.C.; Hermans, M.; Smits, P.H.; van der Zanden, A.G.; Te Witt, R.; Bruijnesteijn van Coppenraet, L.E.; Cohen Stuart, J.; et al. Multi-centre evaluation of real-time multiplex PCR for detection of carbapenemase genes OXA-48, VIM, IMP, NDM and KPC. BMC Infect. Dis. 2014, 14, 27. [Google Scholar] [CrossRef] [Green Version]
- Probst, K.; Boutin, S.; Bandilla, M.; Heeg, K.; Dalpke, A.H. Fast and automated detection of common carbapenemase genes using multiplex real-time PCR on the BD MAX™ system. J. Microbiol. Methods 2021, 185, 106224. [Google Scholar] [CrossRef]
- Berneking, L.; Both, A.; Berinson, B.; Hoffmann, A.; Lütgehetmann, M.; Aepfelbacher, M.; Rohde, H. Performance of the BD Phoenix CPO detect assay for detection and classification of carbapenemase-producing organisms. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 979–985. [Google Scholar] [CrossRef]
- Poirel, L.; Walsh, T.R.; Cuvillier, V.; Nordmann, P. Multiplex PCR for detection of acquired carbapenemase genes. Diagn. Microbiol. Infect. Dis. 2011, 70, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Leslie, D.E.; Azzato, F.; Karapanagiotidis, T.; Leydon, J.; Fyfe, J. Development of a real-time PCR assay to detect Treponema pallidum in clinical specimens and assessment of the assay’s performance by comparison with serological testing. J. Clin. Microbiol. 2007, 45, 93–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.Y.; Chi, K.H.; Alexander, S.; Ison, C.A.; Ballard, R.C. A real-time quadriplex PCR assay for the diagnosis of rectal lymphogranuloma venereum and non-lymphogranuloma venereum Chlamydia trachomatis infections. Sex. Transm. Infect. 2008, 84, 273–276. [Google Scholar] [CrossRef]
- Jensen, J.S.; Björnelius, E.; Dohn, B.; Lidbrink, P. Use of TaqMan 5’ nuclease real-time PCR for quantitative detection of Mycoplasma genitalium DNA in males with and without urethritis who were attendees at a sexually transmitted disease clinic. J. Clin. Microbiol. 2004, 42, 683–692. [Google Scholar] [CrossRef] [Green Version]
- Bayette, J.; Jreige, R.; Marchandin, H.; Laurens, C.; Joullié, F.; Clarivet, B.; Sebbane, M.; Jean-Pierre, H. Prévalence des infections à Chlamydia trachomatis, Neisseria gonorrhoeae et Mycoplasma genitalium chez des patients admis aux urgences [Prevalence of Chlamydia trachomatis, Neisseria gonorrhoeae and Mycoplasma genitalium infections in the emergency department]. Pathol. Biol. 2013, 61, 245–249. [Google Scholar] [PubMed]
- Goire, N.; Nissen, M.D.; LeCornec, G.M.; Sloots, T.P.; Whiley, D.M. A duplex Neisseria gonorrhoeae real-time polymerase chain reaction assay targeting the gonococcal porA pseudogene and multicopy opa genes. Diagn. Microbiol. Infect. Dis. 2008, 61, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Frickmann, H.; Lunardon, L.M.; Hahn, A.; Loderstädt, U.; Lindner, A.K.; Becker, S.L.; Mockenhaupt, F.P.; Weber, C.; Tannich, E. Evaluation of a duplex real-time PCR in human serum for simultaneous detection and differentiation of Schistosoma mansoni and Schistosoma haematobium infections—Cross-sectional study. Travel Med. Infect. Dis. 2021, 41, 102035. [Google Scholar] [CrossRef]
- Niesters, H.G. Quantitation of viral load using real-time amplification techniques. Methods 2001, 25, 419–429. [Google Scholar] [CrossRef]
- Baker, G.C.; Smith, J.J.; Cowan, D.A. Review and re-analysis of domain-specific 16S primers. J. Microbiol. Methods 2003, 55, 541–555. [Google Scholar] [CrossRef] [Green Version]
- Apisarnthanarak, A.; Kiratisin, P.; Mundy, L.M. Evaluation of Ochrobactrum intermedium bacteremia in a patient with bladder cancer. Diagn. Microbiol. Infect. Dis. 2005, 53, 153–155. [Google Scholar] [CrossRef] [PubMed]
- Bharucha, T.; Sharma, D.; Sharma, H.; Kandil, H.; Collier, S. Ochromobactrum intermedium: An emerging opportunistic pathogen-case of recurrent bacteraemia associated with infective endocarditis in a haemodialysis patient. New Microbes New Infect. 2016, 15, 14–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shittu, A.O.; Okon, K.; Adesida, S.; Oyedara, O.; Witte, W.; Strommenger, B.; Layer, F.; Nübel, U. Antibiotic resistance and molecular epidemiology of Staphylococcus aureus in Nigeria. BMC Microbiol. 2011, 11, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayepola, O.O.; Olasupo, N.A.; Egwari, L.O.; Becker, K.; Schaumburg, F. Molecular Characterization and Antimicrobial Susceptibility of Staphylococcus aureus Isolates from Clinical Infection and Asymptomatic Carriers in Southwest Nigeria. PLoS ONE 2015, 10, e0137531. [Google Scholar] [CrossRef] [Green Version]
- Titouche, Y.; Houali, K.; Ruiz-Ripa, L.; Vingadassalon, N.; Nia, Y.; Fatihi, A.; Cauquil, A.; Bouchez, P.; Bouhier, L.; Torres, C.; et al. Enterotoxin genes and antimicrobial resistance in Staphylococcus aureus isolated from food products in Algeria. J. Appl. Microbiol. 2020, 29, 1043–1052. [Google Scholar] [CrossRef]
- El-Ashker, M.; Gwida, M.; Monecke, S.; El-Gohary, F.; Ehricht, R.; Elsayed, M.; Akinduti, P.; El-Fateh, M.; Maurischat, S. Antimicrobial resistance pattern and virulence profile of S. aureus isolated from household cattle and buffalo with mastitis in Egypt. Vet. Microbiol. 2020, 240, 108535. [Google Scholar] [CrossRef]
- Alseqely, M.; Newton-Foot, M.; Khalil, A.; El-Nakeeb, M.; Whitelaw, A.; Abouelfetouh, A. Association between fluoroquinolone resistance and MRSA genotype in Alexandria, Egypt. Sci. Rep. 2021, 11, 4253. [Google Scholar] [CrossRef]
- Essien, U.C.; Boswihi, S.S.; Agbakoba, N.R.; Udo, E.E. Description of Methicillin-Susceptible Staphylococcus aureus Clonal Complex 30 Related to the Pandemic Phage Type 80/81 Isolated from Patients in Three Tertiary Hospitals in Jos, North Central Nigeria. Med. Princ. Pract. 2022, 31, 269–275. [Google Scholar] [CrossRef]
- Agyekum, A.; Fajardo-Lubián, A.; Ansong, D.; Partridge, S.R.; Agbenyega, T.; Iredell, J.R. blaCTX-M-15 carried by IncF-type plasmids is the dominant ESBL gene in Escherichia coli and Klebsiella pneumoniae at a hospital in Ghana. Diagn. Microbiol. Infect. Dis. 2016, 84, 328–333. [Google Scholar] [CrossRef]
- Falgenhauer, L.; Imirzalioglu, C.; Oppong, K.; Akenten, C.W.; Hogan, B.; Krumkamp, R.; Poppert, S.; Levermann, V.; Schwengers, O.; Sarpong, N.; et al. Detection and Characterization of ESBL-Producing Escherichia coli From Humans and Poultry in Ghana. Front. Microbiol. 2019, 9, 3358. [Google Scholar] [CrossRef] [Green Version]
- Obeng-Nkrumah, N.; Labi, A.K.; Blankson, H.; Awuah-Mensah, G.; Oduro-Mensah, D.; Anum, J.; Teye, J.; Kwashie, S.D.; Bako, E.; Ayeh-Kumi, P.F.; et al. Household cockroaches carry CTX-M-15-, OXA-48- and NDM-1-producing enterobacteria, and share beta-lactam resistance determinants with humans. BMC Microbiol. 2019, 19, 272. [Google Scholar] [CrossRef] [PubMed]
- Obeng-Nkrumah, N.; Hansen, D.S.; Awuah-Mensah, G.; Blankson, N.K.; Frimodt-Møller, N.; Newman, M.J.; Opintan, J.A.; Krogfelt, K.A. High level of colonization with third-generation cephalosporin-resistant Enterobacterales in African community settings, Ghana. Diagn. Microbiol. Infect. Dis. 2023, 106, 115918. [Google Scholar] [CrossRef] [PubMed]
- Mahazu, S.; Sato, W.; Ayibieke, A.; Prah, I.; Hayashi, T.; Suzuki, T.; Iwanaga, S.; Ablordey, A.; Saito, R. Insights and genetic features of extended-spectrum beta-lactamase producing Escherichia coli isolates from two hospitals in Ghana. Sci. Rep. 2022, 12, 1843. [Google Scholar] [CrossRef] [PubMed]
- Pankok, F.; Taudien, S.; Dekker, D.; Thye, T.; Oppong, K.; Wiafe Akenten, C.; Lamshöft, M.; Jaeger, A.; Kaase, M.; Scheithauer, S.; et al. Epidemiology of Plasmids in Escherichia coli and Klebsiella pneumoniae with Acquired Extended Spectrum Beta-Lactamase Genes Isolated from Chronic Wounds in Ghana. Antibiotics 2022, 11, 689. [Google Scholar] [CrossRef] [PubMed]
- Eibach, D.; Dekker, D.; Gyau Boahen, K.; Wiafe Akenten, C.; Sarpong, N.; Belmar Campos, C.; Berneking, L.; Aepfelbacher, M.; Krumkamp, R.; Owusu-Dabo, E.; et al. Extended-spectrum beta-lactamase-producing Escherichia coli and Klebsiella pneumoniae in local and imported poultry meat in Ghana. Vet. Microbiol. 2018, 217, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Adedze-Kpodo, R.K.; Feglo, P.K.; Agboli, E.; Asmah, R.H.; Kwadzokpui, P.K. Genotypic characterization of extended-spectrum β-lactamase producing urinary isolates among pregnant women in Ho municipality, Ghana. Heliyon 2022, 8, e12513. [Google Scholar] [CrossRef]
- Moirongo, R.M.; Lorenz, E.; Ntinginya, N.E.; Dekker, D.; Fernandes, J.; Held, J.; Lamshöft, M.; Schaumburg, F.; Mangu, C.; Sudi, L.; et al. Regional Variation of Extended-Spectrum Beta-Lactamase (ESBL)-Producing Enterobacterales, Fluoroquinolone-Resistant Salmonella enterica and Methicillin-Resistant Staphylococcus aureus Among Febrile Patients in Sub-Saharan Africa. Front. Microbiol. 2020, 11, 567235. [Google Scholar] [CrossRef]
- Eibach, D.; Belmar Campos, C.; Krumkamp, R.; Al-Emran, H.M.; Dekker, D.; Boahen, K.G.; Kreuels, B.; Adu-Sarkodie, Y.; Aepfelbacher, M.; Park, S.E.; et al. Extended spectrum beta-lactamase producing Enterobacteriaceae causing bloodstream infections in rural Ghana, 2007-2012. Int. J. Med. Microbiol. 2016, 306, 249–254. [Google Scholar] [CrossRef] [Green Version]
- Akenten, C.W.; Ofori, L.A.; Khan, N.A.; Mbwana, J.; Sarpong, N.; May, J.; Thye, T.; Obiri-Danso, K.; Paintsil, E.K.; Fosu, D.; et al. Prevalence, Characterization, and Antimicrobial Resistance of Extended-Spectrum Beta-Lactamase-Producing Escherichia coli from Domestic Free-Range Poultry in Agogo, Ghana. Foodborne Pathog. Dis. 2023, 20, 59–66. [Google Scholar] [CrossRef]
- Agyepong, N.; Govinden, U.; Owusu-Ofori, A.; Amoako, D.G.; Allam, M.; Janice, J.; Pedersen, T.; Sundsfjord, A.; Essack, S. Genomic characterization of multidrug-resistant ESBL-producing Klebsiella pneumoniae isolated from a Ghanaian teaching hospital. Int. J. Infect. Dis. 2019, 85, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Donkor, E.S.; Horlortu, P.Z.; Dayie, N.T.; Obeng-Nkrumah, N.; Labi, A.K. Community acquired urinary tract infections among adults in Accra, Ghana. Infect. Drug. Resist. 2019, 12, 2059–2067. [Google Scholar] [CrossRef] [Green Version]
- Quansah, E.; Amoah Barnie, P.; Omane Acheampong, D.; Obiri-Yeboah, D.; Odarkor Mills, R.; Asmah, E.; Cudjoe, O.; Dadzie, I. Geographical Distribution of β-Lactam Resistance among Klebsiella spp. from Selected Health Facilities in Ghana. Trop. Med. Infect. Dis. 2019, 4, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohene Larbi, R.; Ofori, L.A.; Sylverken, A.A.; Ayim-Akonor, M.; Obiri-Danso, K. Antimicrobial Resistance of Escherichia coli from Broilers, Pigs, and Cattle in the Greater Kumasi Metropolis, Ghana. Int. J. Microbiol. 2021, 2021, 5158185. [Google Scholar] [CrossRef] [PubMed]
- Janice, J.; Agyepong, N.; Owusu-Ofori, A.; Govinden, U.; Essack, S.Y.; Samuelsen, Ø.; Sundsfjord, A.; Pedersen, T. Carbapenem Resistance Determinants Acquired through Novel Chromosomal Integrations in Extensively Drug-Resistant Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2021, 65, e0028921. [Google Scholar] [CrossRef] [PubMed]
- Olu-Taiwo, M.A.; Opintan, J.A.; Codjoe, F.S.; Obeng Forson, A. Metallo-Beta-Lactamase-Producing Acinetobacter spp. from Clinical Isolates at a Tertiary Care Hospital in Ghana. Biomed Res. Int. 2020, 2020, 3852419. [Google Scholar] [CrossRef]
- Labi, A.K.; Bjerrum, S.; Enweronu-Laryea, C.C.; Ayibor, P.K.; Nielsen, K.L.; Marvig, R.L.; Newman, M.J.; Andersen, L.P.; Kurtzhals, J.A.L. High Carriage Rates of Multidrug-Resistant Gram-Negative Bacteria in Neonatal Intensive Care Units From Ghana. Open Forum Infect. Dis. 2020, 7, ofaa109. [Google Scholar] [CrossRef]
- Ayibieke, A.; Sato, W.; Mahazu, S.; Prah, I.; Addow-Thompson, J.; Ohashi, M.; Suzuki, T.; Iwanaga, S.; Ablordey, A.; Saito, R. Molecular characterisation of the NDM-1-encoding plasmid p2189-NDM in an Escherichia coli ST410 clinical isolate from Ghana. PLoS ONE 2018, 13, e0209623. [Google Scholar] [CrossRef] [Green Version]
- Codjoe, F.S.; Donkor, E.S.; Smith, T.J.; Miller, K. Phenotypic and Genotypic Characterization of Carbapenem-Resistant Gram-Negative Bacilli Pathogens from Hospitals in Ghana. Microb. Drug. Resist. 2019, 25, 1449–1457. [Google Scholar] [CrossRef]
- Codjoe, F.S.; Brown, C.A.; Smith, T.J.; Miller, K.; Donkor, E.S. Genetic relatedness in carbapenem-resistant isolates from clinical specimens in Ghana using ERIC-PCR technique. PLoS ONE 2019, 14, e0222168. [Google Scholar] [CrossRef] [Green Version]
- Ayibieke, A.; Kobayashi, A.; Suzuki, M.; Sato, W.; Mahazu, S.; Prah, I.; Mizoguchi, M.; Moriya, K.; Hayashi, T.; Suzuki, T.; et al. Prevalence and Characterization of Carbapenem-Hydrolyzing Class D β-Lactamase-Producing Acinetobacter Isolates From Ghana. Front. Microbiol. 2020, 11, 587398. [Google Scholar] [CrossRef]
- Acolatse, J.E.E.; Portal, E.A.R.; Boostrom, I.; Akafity, G.; Dakroah, M.P.; Chalker, V.J.; Sands, K.; Spiller, O.B. Environmental surveillance of ESBL and carbapenemase-producing gram-negative bacteria in a Ghanaian Tertiary Hospital. Antimicrob. Resist. Infect. Control. 2022, 11, 49. [Google Scholar] [CrossRef] [PubMed]
- Dwomoh, F.P.; Kotey, F.C.N.; Dayie, N.T.K.D.; Osei, M.M.; Amoa-Owusu, F.; Bannah, V.; Alzahrani, F.M.; Halawani, I.F.; Alzahrani, K.J.; Egyir, B.; et al. Phenotypic and genotypic detection of carbapenemase-producing Escherichia coli and Klebsiella pneumoniae in Accra, Ghana. PLoS ONE 2022, 17, e0279715. [Google Scholar] [CrossRef] [PubMed]
- Monnheimer, M.; Cooper, P.; Amegbletor, H.K.; Pellio, T.; Groß, U.; Pfeifer, Y.; Schulze, M.H. High Prevalence of Carbapenemase-Producing Acinetobacter baumannii in Wound Infections, Ghana, 2017/2018. Microorganisms 2021, 9, 537. [Google Scholar] [CrossRef]
- Labi, A.K.; Nielsen, K.L.; Marvig, R.L.; Bjerrum, S.; Enweronu-Laryea, C.; Bennedbæk, M.; Newman, M.J.; Ayibor, P.K.; Andersen, L.P.; Kurtzhals, J.A.L. Oxacillinase-181 Carbapenemase-Producing Klebsiella pneumoniae in Neonatal Intensive Care Unit, Ghana, 2017–2019. Emerg. Infect. Dis. 2020, 26, 2235–2238. [Google Scholar] [CrossRef]
- Prah, I.; Ayibieke, A.; Mahazu, S.; Sassa, C.T.; Hayashi, T.; Yamaoka, S.; Suzuki, T.; Iwanaga, S.; Ablordey, A.; Saito, R. Emergence of oxacillinase-181 carbapenemase-producing diarrheagenic Escherichia coli in Ghana. Emerg. Microbes Infect. 2021, 10, 865–873. [Google Scholar] [CrossRef]
- Chihi, H.; Bonnin, R.A.; Bourouis, A.; Mahrouki, S.; Besbes, S.; Moussa, M.B.; Belhadj, O.; Naas, T. GES-11-producing Acinetobacter baumannii clinical isolates from Tunisian hospitals: Long-term dissemination of GES-type carbapenemases in North Africa. J. Glob. Antimicrob. Resist. 2016, 5, 47–50. [Google Scholar] [CrossRef]
- Pedersen, T.; Sekyere, J.O.; Govinden, U.; Moodley, K.; Sivertsen, A.; Samuelsen, Ø.; Essack, S.Y.; Sundsfjord, A. Spread of Plasmid-Encoded NDM-1 and GES-5 Carbapenemases among Extensively Drug-Resistant and Pandrug-Resistant Clinical Enterobacteriaceae in Durban, South Africa. Antimicrob. Agents Chemother. 2018, 62, e02178-17. [Google Scholar] [CrossRef] [Green Version]
- Karlowsky, J.A.; Bouchillon, S.K.; El Mahdy Kotb, R.; Mohamed, N.; Stone, G.G.; Sahm, D.F. Carbapenem-resistant Enterobacterales and Pseudomonas aeruginosa causing infection in Africa and the Middle East: A surveillance study from the ATLAS programme (2018-20). JAC Antimicrob. Resist. 2022, 4, dlac060. [Google Scholar] [CrossRef]
- Bentsi, C.; Klufio, C.A.; Perine, P.L.; Bell, T.A.; Cles, L.D.; Koester, C.M.; Wang, S.P. Genital infections with Chlamydia trachomatis and Neisseria gonorrhoeae in Ghanaian women. Genitourin. Med. 1985, 61, 48–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apea-Kubi, K.A.; Yamaguchi, S.; Sakyi, B.; Kishimoto, T.; Ofori-Adjei, D.; Hagiwara, T. Neisseria gonorrhoea, Chlamydia trachomatis, and Treponema pallidum infection in antenatal and gynecological patients at Korle-Bu Teaching Hospital, Ghana. Jpn. J. Infect. Dis. 2004, 57, 253–256. [Google Scholar] [PubMed]
- Yirenya-Tawiah, D.; Annang, T.N.; Apea-Kubi, K.A.; Lomo, G.; Mensah, D.; Akyeh, L.; Bosompem, K.M. Chlamydia trachomatis and Neisseria gonorrhoeae prevalence among women of reproductive age living in urogenital schistosomiasis endemic area in Ghana. BMC Res. Notes 2014, 7, 349. [Google Scholar] [CrossRef] [PubMed]
- Duplessis, C.; Puplampu, N.; Nyarko, E.; Carroll, J.; Dela, H.; Mensah, A.; Amponsah, A.; Sanchez, J. Gonorrhea surveillance in Ghana, Africa. Mil. Med. 2015, 180, 17–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lokpo, S.Y.; Owusu-Dabo, E.; Deku, J.G.; Orish, V.N.; Kye-Duodu, G.; Ussher, F.A.; Boakye, T.; Adigbli, D.; Ameke, L.S.; Fianko, W.K.; et al. A Comparative Study of the Epidemiology of Treponemal Infection in the Volta and Oti Regions of Ghana: A Five-Year Multisite Parallel Population-Based Analysis vis-à-vis the Sentinel Survey. Biomed Res. Int. 2021, 2021, 4462389. [Google Scholar] [CrossRef] [PubMed]
- Völker, F.; Cooper, P.; Bader, O.; Uy, A.; Zimmermann, O.; Lugert, R.; Groß, U. Prevalence of pregnancy-relevant infections in a rural setting of Ghana. BMC Pregnancy Childbirth 2017, 17, 172. [Google Scholar] [CrossRef] [Green Version]
- Sylverken, A.A.; Owusu-Dabo, E.; Yar, D.D.; Salifu, S.P.; Awua-Boateng, N.Y.; Amuasi, J.H.; Okyere, P.B.; Agyarko-Poku, T. Bacterial etiology of sexually transmitted infections at a STI clinic in Ghana; use of multiplex real time PCR. Ghana. Med. J. 2016, 50, 142–148. [Google Scholar] [CrossRef]
- Weinreich, F.; Weinreich, F.; Hahn, A.; Hagen, R.M.; Rohde, H.; Sarfo, F.S.; Feldt, T.; Dompreh, A.; Asibey, S.O.; Boateng, R.; et al. Screening for Schistosoma spp. and Leishmania spp. DNA in Serum of Ghanaian Patients with Acquired Immunodeficiency. Pathogens 2022, 11, 760. [Google Scholar] [CrossRef] [PubMed]
- Lyons, G.R. Schistosomiasis in north-western Ghana. Bull. World Health Organ. 1974, 51, 621–632. [Google Scholar] [PubMed]
- Szela, E.; Bachicha, J.; Miller, D.; Till, M.; Wilson, J.B. Schistosomiasis and cervical cancer in Ghana. Int. J. Gynaecol. Obstet. 1993, 42, 127–130. [Google Scholar] [CrossRef]
- Ntajal, J.; Evers, M.; Kistemann, T.; Falkenberg, T. Influence of human-surface water interactions on the transmission of urinary schistosomiasis in the Lower Densu River basin, Ghana. Soc. Sci. Med. 2021, 288, 113546. [Google Scholar] [CrossRef]
- Yirenya-Tawiah, D.; Amoah, C.; Apea-Kubi, K.A.; Dade, M.; Ackumey, M.; Annang, T.; Mensah, D.Y.; Bosompem, K.M. A survey of female genital schistosomiasis of the lower reproductive tract in the volta basin of Ghana. Ghana Med. J. 2011, 45, 16–21. [Google Scholar] [CrossRef] [Green Version]
- Aryeetey, M.E.; Wagatsuma, Y.; Yeboah, G.; Asante, M.; Mensah, G.; Nkrumah, F.K.; Kojima, S. Urinary schistosomiasis in southern Ghana: 1. Prevalence and morbidity assessment in three (defined) rural areas drained by the Densu river. Parasitol. Int. 2000, 49, 155–163. [Google Scholar] [CrossRef]
- Scott, D.; Senker, K.; England, E.C. Epidemiology of human Schistosoma haematobium infection around Volta Lake, Ghana, 1973–1975. Bull. World Health Organ. 1982, 60, 89–100. [Google Scholar] [PubMed]
- Bozdĕch, V. Incidence of schistosomiasis in the urban population of Accra, Ghana. Folia Parasitol. 1972, 19, 171–174. [Google Scholar]
- Wolfe, M.S. Urinary schistosomiasis in Ghana: A report of 53 cases, with special reference to pyelographic and cystoscopic abnormalities. Trans. R. Soc. Trop. Med. Hyg. 1967, 61, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Wagatsuma, Y.; Aryeetey, M.E.; Nkrumah, F.K.; Sack, D.A.; Kojima, S. Highly symptom-aware children were heavily infected with urinary schistosomiasis in southern Ghana. Cent. Afr. J. Med. 2003, 49, 16–19. [Google Scholar]
- McCullough, F.S. The distribution of human schistosomiasis and the potential snail hosts in Ghana. West. Afr. Med. J. 1957, 6, 87–97. [Google Scholar]
- Bogoch, I.I.; Koydemir, H.C.; Tseng, D.; Ephraim, R.K.D.; Duah, E.; Tee, J.; Andrews, J.R.; Ozcan, A. Evaluation of a Mobile Phone-Based Microscope for Screening of Schistosoma haematobium Infection in Rural Ghana. Am. J. Trop. Med. Hyg. 2017, 96, 1468–1471. [Google Scholar] [CrossRef]
- Klumpp, R.K.; Chu, K.Y. Importance of the aquatic weed Ceratophyllum to transmission of Schistosoma haematobium in the Volta Lake, Ghana. Bull. World Health Organ. 1980, 58, 791–798. [Google Scholar]
- Rosei, L.; McCullough, F.S.; Odei, M.A. Bilharziasis in the Brong Ahafo Region of Ghana. West. Afr. Med. J. 1966, 15, 75–79. [Google Scholar]
- Amankwa, J.A.; Bloch, P.; Meyer-Lassen, J.; Olsen, A.; Christensen, N.O. Urinary and intestinal schistosomiasis in the Tono Irrigation Scheme, Kassena/Nankana District, upper east region, Ghana. Trop. Med. Parasitol. 1994, 45, 319–323. [Google Scholar]
- Wen, S.T.; Chu, K.Y. Preliminary schistosomiasis survey in the lower Volta River below Akosombo Dam, Ghana. Ann. Trop. Med. Parasitol. 1984, 78, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Anyan, W.K.; Abonie, S.D.; Aboagye-Antwi, F.; Tettey, M.D.; Nartey, L.K.; Hanington, P.C.; Anang, A.K.; Muench, S.B. Concurrent Schistosoma mansoni and Schistosoma haematobium infections in a peri-urban community along the Weija dam in Ghana: A wake up call for effective National Control Programme. Acta Trop. 2019, 199, 105116. [Google Scholar] [CrossRef] [PubMed]
- Feldmeier, H.; Krantz, I.; Poggensee, G. Female genital schistosomiasis as a risk-factor for the transmission of HIV. Int. J. STD AIDS 1994, 5, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Sturt, A.S.; Webb, E.L.; Himschoot, L.; Phiri, C.R.; Mapani, J.; Mudenda, M.; Kjetland, E.F.; Mweene, T.; Levecke, B.; van Dam, G.J.; et al. Association of Female Genital Schistosomiasis With the Cervicovaginal Microbiota and Sexually Transmitted Infections in Zambian Women. Open Forum Infect. Dis. 2021, 8, ofab438. [Google Scholar] [CrossRef]
- Shukla, J.D.; Kleppa, E.; Holmen, S.; Ndhlovu, P.D.; Mtshali, A.; Sebitloane, M.; Vennervald, B.J.; Gundersen, S.G.; Taylor, M.; Kjetland, E.F. The Association Between Female Genital Schistosomiasis and Other Infections of the Lower Genital Tract in Adolescent Girls and Young Women: A Cross-Sectional Study in South Africa. J. Low. Genit. Tract. Dis. 2023, 27, 291–296. [Google Scholar] [CrossRef]

| Country of Origin (n = 274) | |
| Ivory Coast (n, %) | 223, 81.4% |
| Ghana (n, %) | 51, 18.6% |
| Age (n = 274) | |
| Mean (±SD) | 28.4 (±5.8) |
| Median (Min., Max.) | 28 (18, 46) |
| Number of pregnancies (n = 252) | |
| Mean (±SD) | 3.1 (±2.0) |
| Median (Min., Max.) | 3 (1, 9) |
| APGAR 1 score value (n = 268) | |
| Mean (±SD) | 7.9 (±1.1) |
| Median (Min., Max.) | 8 (2, 10) |
| APGAR 2 score value (n = 268) | |
| Mean (±SD) | 8.7 (±0.7) |
| Median (Min., Max.) | 9 (4, 10) |
| Breeding of chicken (n = 274) | |
| Yes (n, %) | 13.9% (38/274) |
| No (n, %) | 86.1% (236/274) |
| Possession of a freezer (n = 274) | |
| Yes (n, %) | 36.1% (99/274) |
| No (n, %) | 63.9% (175/274) |
| Electricity at home (n = 274) | |
| Yes (n, %) | 98.2% (269/274) |
| No (n, %) | 1.8% (5/274) |
| Type of birth (n = 265) | |
| Caesarian section (n, %) | 7.9% (21/265) |
| Vaginal delivery (n, %) | 92.1% (244/265) |
| Floor quality (n = 274) | |
| Earth/sand | 5.5% (15/274) |
| Wooden/bamboo | 0.0% (0/274)) |
| Vinyl/tiles | 18.2% (50/274) |
| Cement | 76.3% (209/274) |
| Other | 0.0% (0/274) |
| Education level (n = 274) | |
| None | 39.8% (109/274) |
| Basic | 29.6% (81/274) |
| Secondary | 22.3% (61/274) |
| Tertiary | 8.4% (23/274) |
| Occupation (n = 274) | |
| Housewife | 21.2% (58/274) |
| Farmer | 0.4% (1/274) |
| Trader | 21.5% (59/274) |
| Salary worker | 10.9% (30/274) |
| Other | 46.0% (126/274) |
| Drinking water source (n = 274) | |
| Surface water | 0.0% (0/274) |
| Tanker | 0.4% (1/274) |
| Well | 3.3% (9/274) |
| Bore hole | 0.0% (0/274) |
| Piped water | 94.9% (260/274) |
| Other | 1.5% (4/274) |
| Toilet (n = 274) | |
| No facility | 0.0% (0/274) |
| Pit latrine | 54.0% (148/274) |
| Improved pit latrine | 26.6% (73/274) |
| Flush toilet | 19.3% (53/274) |
| Other | 0.0% (0/274) |
| PCR Target | Numbers and Proportions of Detections n/n (%) | Mean (SD) of the Measured CT Values | Median (Min., Max.) of the Measured CT Values |
|---|---|---|---|
| blaCTX-M | 42/268 (15.7%) | 30.9 (2.8) | 32 (24, 37) |
| blaNDM | 6/268 (2.2%) | 29.5 (1.9) | 30 (27, 33) |
| blaKPC | 2/268 (0.7%) | 31.0 (1.0) | 31 (30, 32) |
| blaVIM | 12/268 (4.5%) | 30.0 (3.1) | 31 (21, 33) |
| blaIMP | 25/268 (9.3%) | 30.4 (2.0) | 30 (26, 34) |
| blaOXA-23-like | 5/268 (1.9%) | 30.8 (4.5) | 32 (24, 37) |
| blaOXA-40/24-like | 0/268 (0%) | n.e. | n.e. |
| blaOXA-48-like | 5/268 (1.9%) | 31.8 (2.8) | 31 (29, 37) |
| blaOXA-58-like | 10/268 (3.7%) | 34.2 (3.3) | 35.5 (26, 37) |
| blaGES | 20/268 (7.5%) | 35.2 (3.2) | 35 (29, 40) |
| blaNMC_A/IMI | 0/268 (0%) | n.e. | n.e. |
| blaBIC | 0/268 (0%) | n.e. | n.e. |
| blaSME | 0/268 (0%) | n.e. | n.e. |
| blaGIM | 0/268 (0%) | n.e. | n.e. |
| blaDIM | 0/268 (0%) | n.e. | n.e. |
| PCR Target | Numbers and Proportions of Detections n/n (%) | Mean (SD) of the Measured CT Values | Median (Min., Max.) of the Measured CT Values |
|---|---|---|---|
| Treponema pallidum | 0/268 (0%) | n.e. | n.e. |
| Chlamydia trachomatis screening | 3/268 (1.1%) | 31.3 (2.6) | 30 (29, 35) |
| 0/268 (0%) | n.e. | n.e. |
| 3/268 (1.1%) | 34.3 (1.7) | 35 (32, 36) |
| Mycoplasma genitalium | 0/268 (0%) | n.e. | n.e. |
| Neisseria gonorrhoeae * | 5/268 (1.9%) | Please see below! | Please see below! |
| 5/268 (1.9%) | 31.8 (2.2) | 33 (29, 34) |
| 5/268 (1.9%) | 34.0 (2.6) | 33 (31, 38) |
| PCR Target | Numbers and Proportions of Detections n/n (%) | Mean (SD) of the Measured CT Values | Median (Min., Max.) of the Measured CT Values |
|---|---|---|---|
| Schistosoma mansoni complex | 1/268 (0.4%) | 37 (n.a.) | 37 (n.a.) |
| Schistosoma haematobium complex | 50/269 * (18.6%) | 31.0 (1.6) | 32 (27, 34) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franz, R.; Hahn, A.; Hagen, R.M.; Rohde, H.; Eberhardt, K.A.; Ehrhardt, S.; Baum, J.; Claussen, L.; Feldt, T.; Hinz, R.; et al. Screening for Resistant Bacteria, Antimicrobial Resistance Genes, Sexually Transmitted Infections and Schistosoma spp. in Tissue Samples from Predominantly Vaginally Delivered Placentae in Ivory Coast and Ghana. Pathogens 2023, 12, 999. https://doi.org/10.3390/pathogens12080999
Franz R, Hahn A, Hagen RM, Rohde H, Eberhardt KA, Ehrhardt S, Baum J, Claussen L, Feldt T, Hinz R, et al. Screening for Resistant Bacteria, Antimicrobial Resistance Genes, Sexually Transmitted Infections and Schistosoma spp. in Tissue Samples from Predominantly Vaginally Delivered Placentae in Ivory Coast and Ghana. Pathogens. 2023; 12(8):999. https://doi.org/10.3390/pathogens12080999
Chicago/Turabian StyleFranz, Roman, Andreas Hahn, Ralf Matthias Hagen, Holger Rohde, Kirsten Alexandra Eberhardt, Stephan Ehrhardt, Jana Baum, Lisa Claussen, Torsten Feldt, Rebecca Hinz, and et al. 2023. "Screening for Resistant Bacteria, Antimicrobial Resistance Genes, Sexually Transmitted Infections and Schistosoma spp. in Tissue Samples from Predominantly Vaginally Delivered Placentae in Ivory Coast and Ghana" Pathogens 12, no. 8: 999. https://doi.org/10.3390/pathogens12080999
APA StyleFranz, R., Hahn, A., Hagen, R. M., Rohde, H., Eberhardt, K. A., Ehrhardt, S., Baum, J., Claussen, L., Feldt, T., Hinz, R., Barthel, D., Bindt, C., Tagbor, H., Nguah, S. B., Koffi, M., Köller, T., Warnke, P., Pankok, F., Taudien, S., ... Schoppen, S. (2023). Screening for Resistant Bacteria, Antimicrobial Resistance Genes, Sexually Transmitted Infections and Schistosoma spp. in Tissue Samples from Predominantly Vaginally Delivered Placentae in Ivory Coast and Ghana. Pathogens, 12(8), 999. https://doi.org/10.3390/pathogens12080999