
Antimicrobial Effects of Plant-Based Supplements on Gut Microbial Diversity in Small Ruminants



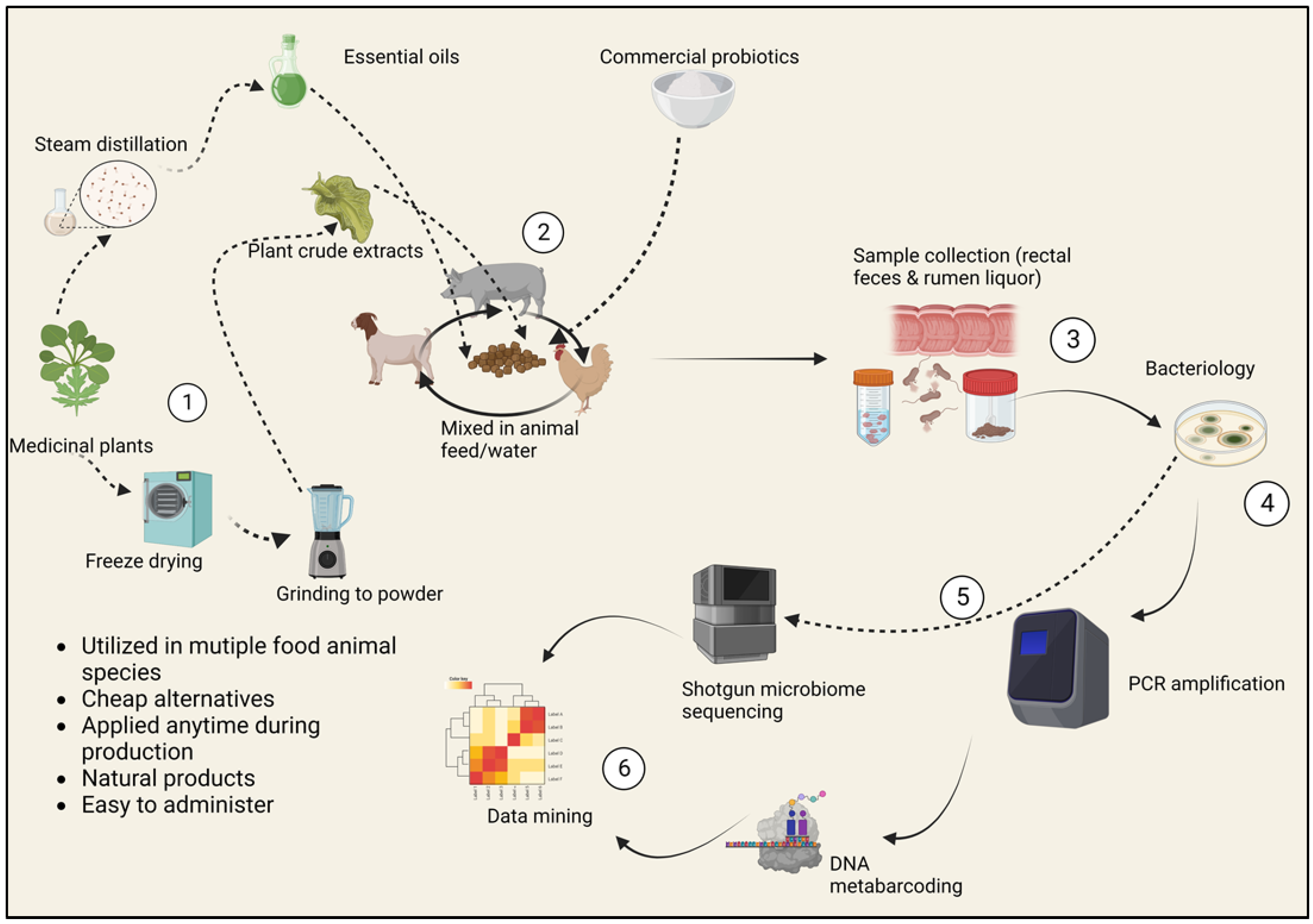{kind=link}
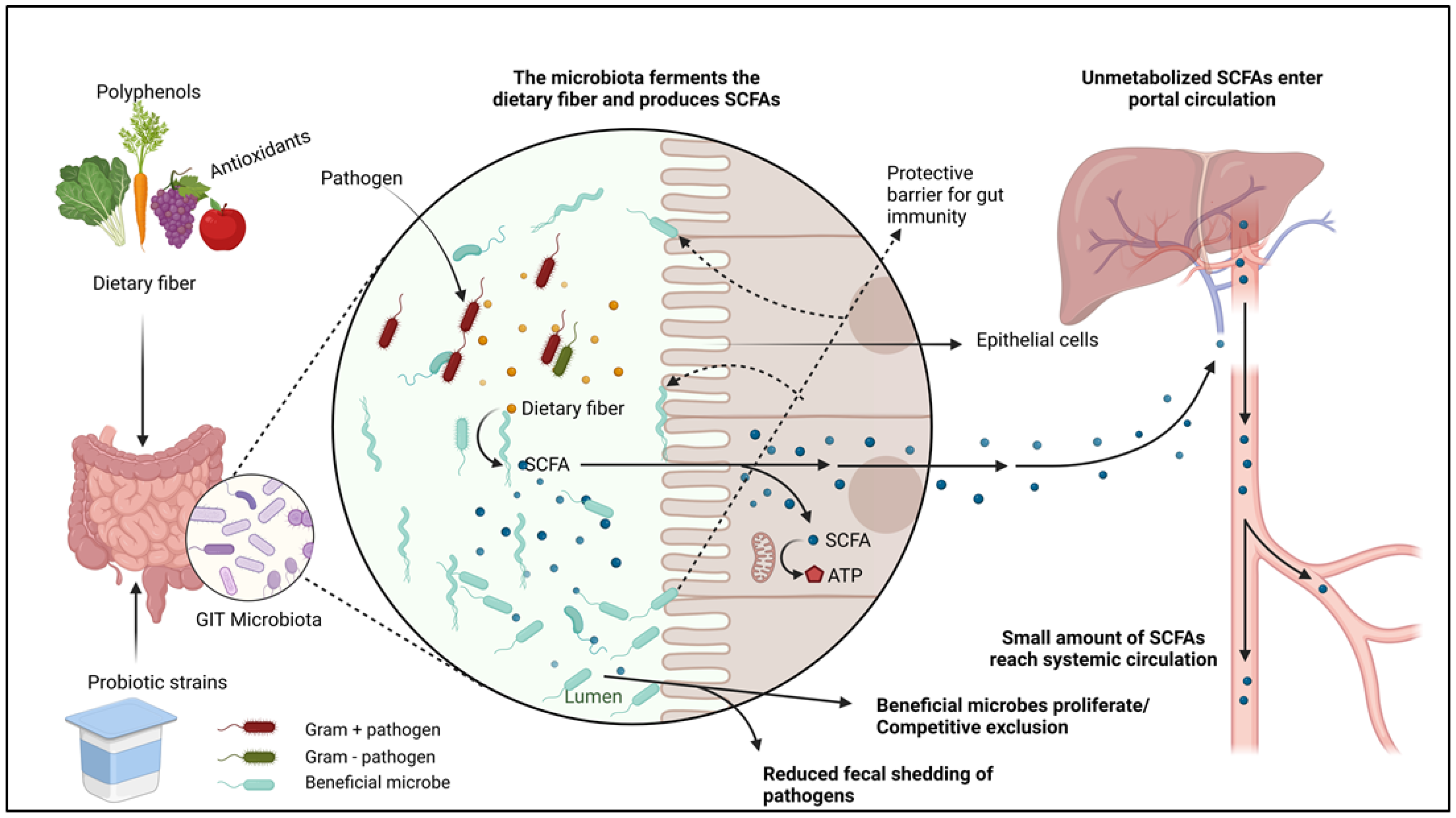{kind=link}
Abstract
:1. Introduction
2. Burden of Pathogens in Small Ruminant Source Foods
3. Global Threat of Antimicrobial Resistance
4. Growing Global Interest in Small Ruminants as a Food Source
5. Small Ruminant Production in the United States
6. The Link between Antimicrobial Use and the Emergence of Antimicrobial Resistant Bacteria in Food Animals
7. Examples of Resistant Animal-Borne Bacterial Species
7.1. Salmonella enterica
7.2. Listeria monocytogenes
7.3. Escherichia coli
7.4. Campylobacter spp.
7.5. Clostridium perfringens
8. The Ruminant Intestinal Microbiota—The Gut Microbiome
9. Impact of Dietary Supplements on the Gut Microbiome
10. Antimicrobial Traits of Plant-Based Essential Oils and Other Plant Extracts
11. Examples of Plant-Based Dietary Supplements with Antimicrobial Effects
11.1. Sweet Potato (Ipomoea batatas) Tops (Leaves and Vines)
11.2. Daikon Radish (Raphanus raphanistrum subsp. sativus)
11.3. Rice Bran (Oryza sativa L)
12. Conclusions
Funding
Conflicts of Interest
References
- Heredia, N.; García, S. Animals as sources of food-borne pathogens: A review. Anim. Nutr. 2018, 4, 250–255. [Google Scholar] [CrossRef]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.-A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States—Major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef]
- White, A.E.; Tillman, A.R.; Hedberg, C.; Bruce, B.B.; Batz, M.; Seys, S.A.; Dewey-Mattia, D.; Bazaco, M.C.; Walter, E.S. Foodborne Illness Outbreaks Reported to National Surveillance, United States, 2009–2018. Emerg Infect Dis 2022, 28, 1117–1127. [Google Scholar] [CrossRef]
- Gourama, H. Foodborne Pathogens; Springer International Publishing: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Lee, H.; Yoon, Y. Etiological Agents Implicated in Foodborne Illness World Wide. Food Sci. Anim. Resour. 2021, 41, 1–7. [Google Scholar] [CrossRef]
- Pickard, J.M.; Zeng, M.Y.; Caruso, R.; Núñez, G. Gut microbiota: Role in pathogen colonization, immune responses, and inflammatory disease. Immunol. Rev. 2017, 279, 70–89. [Google Scholar] [CrossRef]
- Callaway, T.; Edrington, T.; Anderson, R.; Harvey, R.; Genovese, K.; Kennedy, C.; Venn, D.; Nisbet, D. Probiotics, prebiotics and competitive exclusion for prophylaxis against bacterial disease. Anim. Health Res. Rev. 2008, 9, 217–225. [Google Scholar] [CrossRef]
- Doyle, M.P.; Erickson, M.C. Reducing the carriage of foodborne pathogens in livestock and poultry. Poult. Sci. 2006, 85, 960–973. [Google Scholar] [CrossRef]
- Gaggìa, F.; Mattarelli, P.; Biavati, B. Probiotics and prebiotics in animal feeding for safe food production. Int. J. Food Microbiol. 2010, 141 (Suppl. 1), S15–S28. [Google Scholar] [CrossRef]
- Jensen, A.P.; Bjørnvad, C.R. Clinical effect of probiotics in prevention or treatment of gastrointestinal disease in dogs: A systematic review. J. Vet. Intern. Med. 2019, 33, 1849–1864. [Google Scholar] [CrossRef]
- Wegener, H.C. Antibiotics in animal feed and their role in resistance development. Curr. Opin. Microbiol. 2003, 6, 439–445. [Google Scholar] [CrossRef]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef] [PubMed]
- Roca, I.; Akova, M.; Baquero, F.; Carlet, J.; Cavaleri, M.; Coenen, S.; Cohen, J.; Findlay, D.; Gyssens, I.; Heure, O. The global threat of antimicrobial resistance: Science for intervention. New Microbes New Infect. 2015, 6, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Looft, T.; Johnson, T.A.; Allen, H.K.; Bayles, D.O.; Alt, D.P.; Stedtfeld, R.D.; Sul, W.J.; Stedtfeld, T.M.; Chai, B.; Cole, J.R. In-feed antibiotic effects on the swine intestinal microbiome. Proc. Natl. Acad. Sci. USA 2012, 109, 1691–1696. [Google Scholar] [CrossRef] [PubMed]
- Maron, D.F.; Smith, T.J.; Nachman, K.E. Restrictions on antimicrobial use in food animal production: An international regulatory and economic survey. Glob. Health 2013, 9, 48. [Google Scholar] [CrossRef] [PubMed]
- Bintsis, T. Foodborne pathogens. AIMS Microbiol. 2017, 3, 529. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Havelaar, A.H.; Hoffmann, S.; Hald, T.; Kirk, M.D.; Torgerson, P.R.; Devleesschauwer, B. Global disease burden of pathogens in animal source foods, 2010. PLoS ONE 2019, 14, e0216545. [Google Scholar] [CrossRef] [PubMed]
- Dawson, L.; Allen, J.; Olcott, B. Meat goat herd health procedures and prevention. In Proceedings of the 22nd Annual Goat Field Day; Langston University: Langston, OK, USA, 2007. [Google Scholar]
- Redel, H. Foodborne infections and intoxications. Emerg. Infect. Dis. 2013, 19, 2067. [Google Scholar] [CrossRef]
- Founou, L.L.; Founou, R.C.; Essack, S.Y. Antibiotic resistance in the food chain: A developing country-perspective. Front. Microbiol. 2016, 7, 1881. [Google Scholar] [CrossRef]
- Cassini, A.; Högberg, L.D.; Plachouras, D.; Quattrocchi, A.; Hoxha, A.; Simonsen, G.S.; Colomb-Cotinat, M.; Kretzschmar, M.E.; Devleesschauwer, B.; Cecchini, M. Attributable deaths and disability-adjusted life-years caused by infections with antibiotic-resistant bacteria in the EU and the European Economic Area in 2015: A population-level modelling analysis. Lancet Infect. Dis. 2019, 19, 56–66. [Google Scholar] [CrossRef]
- Hansson, I.; Hamilton, C.; Ekman, T.; Forslund, K. Carcass quality in certified organic production compared with conventional livestock production. J. Vet. Med. Ser. B 2000, 47, 111–120. [Google Scholar] [CrossRef]
- Hanlon, K.E.; Miller, M.F.; Guillen, L.M.; Echeverry, A.; Dormedy, E.; Cemo, B.; Branham, L.A.; Sanders, S.; Brashears, M.M. Presence of Salmonella and Escherichia coli O157 on the hide, and presence of Salmonella, Escherichia coli O157 and Campylobacter in feces from small-ruminant (goat and lamb) samples collected in the United States, Bahamas and Mexico. Meat Sci. 2018, 135, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Hanlon, K.; Brashears, M.; Miller, M. Presence of Salmonella on the Carcass, Hide and Feces of Goats and Lambs from Major Livestock Shows in Texas Collected Over 4 Yr. Meat Muscle Biol. 2019, 1, 130. [Google Scholar] [CrossRef]
- Wall, B.; Mateus, A.; Marshall, L.; Pfeiffer, D.; Lubroth, J.; Ormel, H.; Otto, P.; Patriarchi, A. Drivers, Dynamics and Epidemiology of Antimicrobial Resistance in Animal Production; Food and Agriculture Organization of the United Nations: Rome, Italy, 2016. [Google Scholar]
- Otokunefor, K.; Agbagwa, O.E.; Oshifade, E.; Ibezim, C.N.E.; Azi, E. Antimicrobial Resistance: A Collective Responsibility; Springer Nature: Cham, Switzerland, 2023; pp. 89–98. [Google Scholar] [CrossRef]
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Tagliabue, A.; Rappuoli, R. Changing priorities in vaccinology: Antibiotic resistance moving to the top. Front. Immunol. 2018, 9, 1068. [Google Scholar] [CrossRef] [PubMed]
- Boucher, H.W. Bad bugs, no drugs 2002–2020: Progress, challenges, and call to action. Trans. Am. Clin. Climatol. Assoc. 2020, 131, 65. [Google Scholar] [PubMed]
- Toner, E.; Adalja, A.; Gronvall, G.K.; Cicero, A.; Inglesby, T.V. Antimicrobial Resistance Is a Global Health Emergency. Health Secur. 2015, 13, 153–155. [Google Scholar] [CrossRef] [PubMed]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: A global multifaceted phenomenon. Pathog. Glob. Health 2015, 109, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Kawecki, D.; Pacholczyk, M.; Lagiewska, B.; Sawicka-Grzelak, A.; Durlik, M.; Mlynarczyk, G.; Chmura, A. Bacterial and fungal infections in the early post-transplantation period after liver transplantation: Etiologic agents and their susceptibility. In Transplantation Proceedings; Elsevier: Amsterdam, The Netherlands, 2014; pp. 2777–2781. [Google Scholar]
- Teoh, F.; Pavelka, N. How chemotherapy increases the risk of systemic candidiasis in cancer patients: Current paradigm and future directions. Pathogens 2016, 5, 6. [Google Scholar] [CrossRef]
- Miller, B.A.; Lu, C.D. Current status of global dairy goat production: An overview. Asian-Australas. J. Anim. Sci. 2019, 32, 1219. [Google Scholar] [CrossRef]
- Monteiro, A.L.G.; Faro, A.M.C.d.F.; Peres, M.T.P.; Batista, R.; Poli, C.H.E.C.; Villalba, J.J. The role of small ruminants on global climate change. Acta Scientiarum. Anim. Sci. 2018, 40, 43124. [Google Scholar] [CrossRef]
- Panel, M.M. Meat, Milk and More: Policy Innovations to Shepherd Inclusive and Sustainable Livestock Systems in Africa; The International Food Policy Research Institute: Washington, DC, USA, 2020. [Google Scholar]
- Guerra, M.M.M.; De Almeida, A.M.; Willingham, A.L. An overview of food safety and bacterial foodborne zoonoses in food production animals in the Caribbean region. Trop. Anim. Health Prod. 2016, 48, 1095–1108. [Google Scholar] [CrossRef]
- Foote, W. History and accomplishments of the International Goat Association during the last 20 years. Small Rumin. Res. 2004, 51, 115–121. [Google Scholar] [CrossRef]
- Marino, R.; Atzori, A.S.; D’andrea, M.; Iovane, G.; Trabalza-Marinucci, M.; Rinaldi, L. Climate change: Production performance, health issues, greenhouse gas emissions and mitigation strategies in sheep and goat farming. Small Rumin. Res. 2016, 135, 50–59. [Google Scholar] [CrossRef]
- Sherman, D. The spread of pathogens through trade in small ruminants and their products. Rev. Sci. Et. Tech.-OIE 2011, 30, 207. [Google Scholar] [CrossRef]
- Okpebholo, F.; Kahan, T. Opportunities and challenges for developing small ruminant systems. Caprine Chron. Off. Newsl. Fla. Meat Goat Assoc. 2007, 22. [Google Scholar]
- Coffey, L. Meat Goats: Sustainable Production; National Sustainable Agriculture Information Service: Arkansas, CA, USA, 2006; Available online: https://attra.ncat.org/publication/meat-goats-sustainable-production/ (accessed on 1 November 2023).
- Albenzio, M.; Santillo, A.; Avondo, M.; Nudda, A.; Chessa, S.; Pirisi, A.; Banni, S. Nutritional properties of small ruminant food products and their role on human health. Small Rumin. Res. 2016, 135, 3–12. [Google Scholar] [CrossRef]
- Animal and Plant Health Inspection Service; Veterinary Services; Center for Epidemiology and Animal Health. Small-Scale U.S. Goat Operations; United States Department of Agriculture, National Animal Health Monitoring System: Fort Collins, CO, USA, 2011; p. 41. [Google Scholar]
- Milani, F.; Wendorff, W. Goat and sheep milk products in the United States (USA). Small Rumin. Res. 2011, 101, 134–139. [Google Scholar] [CrossRef]
- Economou, V.; Gousia, P. Agriculture and food animals as a source of antimicrobial-resistant bacteria. Infect. Drug Resist. 2015, 8, 49–61. [Google Scholar] [CrossRef]
- Prescott, J.F. Antimicrobial use in food and companion animals. Anim. Health Res. Rev. 2008, 9, 127–133. [Google Scholar] [CrossRef]
- Anderson, A.D.; Nelson, J.M.; Rossiter, S.; Angulo, F.J. Public health consequences of use of antimicrobial agents in food animals in the United States. Microb. Drug Resist. 2003, 9, 373–379. [Google Scholar] [CrossRef]
- Teale, C. Antimicrobial resistance and the food chain. J. Appl. Microbiol. 2002, 92, 85S–89S. [Google Scholar] [CrossRef]
- Piñeiro, S.A.; Cerniglia, C.E. Antimicrobial drug residues in animal-derived foods: Potential impact on the human intestinal microbiome. J. Vet. Pharmacol. Ther. 2021, 44, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Reig, M.; Toldrá, F. Veterinary drug residues in meat: Concerns and rapid methods for detection. Meat Sci. 2008, 78, 60–67. [Google Scholar] [CrossRef]
- Moreno, L.; Lanusse, C. Veterinary drug residues in meat-related edible tissues. In New Aspects of Meat Quality; Elsevier: Amsterdam, The Netherlands, 2017; pp. 581–603. [Google Scholar]
- Singh, S.; Shukla, S.; Tandia, N.; Kumar, N.; Paliwal, R. Antibiotic Residues: A Global Challenge. Pharma Sci. Monit. 2014, 5, 184–197. [Google Scholar]
- Beyene, T. Veterinary drug residues in food-animal products: Its risk factors and potential effects on public health. J. Vet. Sci. Technol. 2016, 7, 1–7. [Google Scholar] [CrossRef]
- Cameron, A.; McAllister, T.A. Antimicrobial usage and resistance in beef production. J. Anim. Sci. Biotechnol. 2016, 7, 68. [Google Scholar] [CrossRef] [PubMed]
- Shterzer, N.; Mizrahi, I. The animal gut as a melting pot for horizontal gene transfer. Can. J. Microbiol. 2015, 61, 603–605. [Google Scholar] [CrossRef]
- Hammerum, A.M.; Heuer, O.E. Human health hazards from antimicrobial-resistant Escherichia coli of animal origin. Clin. Infect. Dis. 2009, 48, 916–921. [Google Scholar] [CrossRef] [PubMed]
- Meek, R.W.; Vyas, H.; Piddock, L.J.V. Nonmedical uses of antibiotics: Time to restrict their use? PLoS Biol. 2015, 13, e1002266. [Google Scholar] [CrossRef]
- Auffret, M.D.; Dewhurst, R.J.; Duthie, C.-A.; Rooke, J.A.; John Wallace, R.; Freeman, T.C.; Stewart, R.; Watson, M.; Roehe, R. The rumen microbiome as a reservoir of antimicrobial resistance and pathogenicity genes is directly affected by diet in beef cattle. Microbiome 2017, 5, 159. [Google Scholar] [CrossRef]
- Ji, S.; Jiang, T.; Yan, H.; Guo, C.; Liu, J.; Su, H.; Alugongo, G.M.; Shi, H.; Wang, Y.; Cao, Z.; et al. Ecological Restoration of Antibiotic-Disturbed Gastrointestinal Microbiota in Foregut and Hindgut of Cows. Front. Cell. Infect. Microbiol. 2018, 8, 79. [Google Scholar] [CrossRef]
- Shen, H.; Lu, Z.; Xu, Z.; Shen, Z. Antibiotic pretreatment minimizes dietary effects on reconstructure of rumen fluid and mucosal microbiota in goats. MicrobiologyOpen 2018, 7, e00537. [Google Scholar] [CrossRef] [PubMed]
- Lammie, S.L.; Hughes, J.M. Antimicrobial resistance, food safety, and one health: The need for convergence. Annu. Rev. Food Sci. Technol. 2016, 7, 287–312. [Google Scholar] [CrossRef] [PubMed]
- Aljahdali, N.H.; Sanad, Y.M.; Han, J.; Foley, S.L. Current knowledge and perspectives of potential impacts of Salmonella enterica on the profile of the gut microbiota. BMC Microbiol. 2020, 20, 353. [Google Scholar] [CrossRef] [PubMed]
- Singh, V. Salmonella serovars and their host specificity. J. Vet. Sci. Anim. Husb. 2013, 1, 301. [Google Scholar] [CrossRef]
- Jajere, S.M. A review of Salmonella enterica with particular focus on the pathogenicity and virulence factors, host specificity and antimicrobial resistance including multidrug resistance. Vet. World 2019, 12, 504. [Google Scholar] [CrossRef] [PubMed]
- Bäumler, A.; Fang, F.C. Host specificity of bacterial pathogens. Cold Spring Harb. Perspect. Med. 2013, 3, a010041. [Google Scholar] [CrossRef] [PubMed]
- Hempstead, S.C.; Gensler, C.A.; Keelara, S.; Brennan, M.; Urie, N.J.; Wiedenheft, A.M.; Marshall, K.L.; Morningstar-Shaw, B.; Lantz, K.; Fedorka-Cray, P.J. Detection and molecular characterization of Salmonella species on US goat operations. Prev. Vet. Med. 2022, 208, 105766. [Google Scholar] [CrossRef] [PubMed]
- Popa, G.L.; Papa, M.I. Salmonella spp. Infection—A continuous threat worldwide. Germs 2021, 11, 88–96. [Google Scholar] [CrossRef]
- Webb, H.E.; Brichta-Harhay, D.M.; Brashears, M.M.; Nightingale, K.K.; Arthur, T.M.; Bosilevac, J.M.; Kalchayanand, N.; Schmidt, J.W.; Wang, R.; Granier, S.A. Salmonella in peripheral lymph nodes of healthy cattle at slaughter. Front. Microbiol. 2017, 8, 2214. [Google Scholar] [CrossRef]
- Haneklaus, A.N.; Harris, K.B.; Griffin, D.B.; Edrington, T.S.; Lucia, L.M.; Savell, J.W. Salmonella prevalence in bovine lymph nodes differs among feedyards. J. Food Prot. 2012, 75, 1131–1133. [Google Scholar] [CrossRef]
- Gragg, S.E.; Loneragan, G.H.; Brashears, M.M.; Arthur, T.M.; Bosilevac, J.M.; Kalchayanand, N.; Wang, R.; Schmidt, J.W.; Brooks, J.C.; Shackelford, S.D. Cross-sectional study examining Salmonella enterica carriage in subiliac lymph nodes of cull and feedlot cattle at harvest. Foodborne Pathog. Dis. 2013, 10, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Besser, J.M. Salmonella epidemiology: A whirlwind of change. Food Microbiol. 2018, 71, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Quereda, J.J.; Leclercq, A.; Moura, A.; Vales, G.; Gómez-Martín, Á.; García-Muñoz, Á.; Thouvenot, P.; Tessaud-Rita, N.; Bracq-Dieye, H.; Lecuit, M. Listeria valentina sp. nov., isolated from a water trough and the faeces of healthy sheep. Int. J. Syst. Evol. Microbiol. 2020, 70, 5868–5879. [Google Scholar] [CrossRef] [PubMed]
- Schoder, D.; Pelz, A.; Paulsen, P. Transmission Scenarios of Listeria monocytogenes on Small Ruminant On-Farm Dairies. Foods 2023, 12, 265. [Google Scholar] [CrossRef] [PubMed]
- de Noordhout, C.M.; Devleesschauwer, B.; Angulo, F.J.; Verbeke, G.; Haagsma, J.; Kirk, M.; Havelaar, A.; Speybroeck, N. The global burden of listeriosis: A systematic review and meta-analysis. Lancet Infect. Dis. 2014, 14, 1073–1082. [Google Scholar] [CrossRef] [PubMed]
- Cardenas-Alvarez, M.X.; Zeng, H.; Webb, B.T.; Mani, R.; Muñoz, M.; Bergholz, T.M. Comparative Genomics of Listeria monocytogenes Isolates from Ruminant Listeriosis Cases in the Midwest United States. Microbiol. Spectr. 2022, 10, e01579-22. [Google Scholar] [CrossRef] [PubMed]
- Condoleo, R.; Giangolini, G.; Chiaverini, A.; Patriarca, D.; Scaramozzino, P.; Mezher, Z. Occurrence of Listeria monocytogenes and Escherichia coli in raw sheep’s milk from farm bulk tanks in Central Italy. J. Food Prot. 2020, 83, 1929–1933. [Google Scholar] [CrossRef]
- Hazards, E.P.o.B.; Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Fernández Escámez, P.S.; Girones, R.; Herman, L.; Koutsoumanis, K. Listeria monocytogenes contamination of ready-to-eat foods and the risk for human health in the EU. EFSA J. 2018, 16, e05134. [Google Scholar]
- Baher, W.; Shalaby, M.; Abdelghfar, S. Prevalence of multidrug-resistant Listeria monocytogenes in retailed goat meat and offal. Damanhour J. Vet. Sci. 2021, 7, 19–22. [Google Scholar] [CrossRef]
- Smith, S.; Wang, J.; Fanning, S.; McMahon, B.J. Antimicrobial resistant bacteria in wild mammals and birds: A coincidence or cause for concern? Ir. Vet. J. 2014, 67, 8. [Google Scholar] [CrossRef]
- Bélanger, L.; Garenaux, A.; Harel, J.; Boulianne, M.; Nadeau, E.; Dozois, C.M. Escherichia coli from animal reservoirs as a potential source of human extraintestinal pathogenic E. coli. FEMS Immunol. Med. Microbiol. 2011, 62, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Havelaar, A.H.; Van Duynhoven, Y.T.H.P.; Nauta, M.J.; Bouwknegt, M.; Heuvelink, A.E.; De Wit, G.A.; Nieuwenhuizen, M.G.M.; Van De Kar, N.C.A.J. Disease burden in The Netherlands due to infections with Shiga toxin-producing Escherichia coli O157. Epidemiol. Infect. 2004, 132, 467–484. [Google Scholar] [CrossRef] [PubMed]
- Boyer, O.; Niaudet, P. Hemolytic uremic syndrome: New developments in pathogenesis and treatment. Int. J. Nephrol. 2011, 2011, 908407. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, P.; Singh, N.S.; Virdi, J.S. Escherichia coli β-Lactamases: What Really Matters. Front. Microbiol. 2016, 7, 417. [Google Scholar] [CrossRef] [PubMed]
- Awosile, B.; Fritzler, J.; Levent, G.; Rahman, M.K.; Ajulo, S.; Daniel, I.; Tasnim, Y.; Sarkar, S. Genomic Characterization of Fecal Escherichia coli Isolates with Reduced Susceptibility to Beta-Lactam Antimicrobials from Wild Hogs and Coyotes. Pathogens 2023, 12, 929. [Google Scholar] [CrossRef] [PubMed]
- Obaidat, M.M.; Tarazi, Y.H.; AlSmadi, W.M. Sheep and goats are reservoirs of colistin resistant Escherichia coli that co-resist critically important antimicrobials: First study from Jordan. J. Food Saf. 2023, 43, e13023. [Google Scholar] [CrossRef]
- Ruiz-Palacios, G.M. The Health Burden of Campylobacter Infection and the Impact of Antimicrobial Resistance: Playing Chicken. Clin. Infect. Dis. 2007, 44, 701–703. [Google Scholar] [CrossRef]
- Denis, M.; Rose, V.; Nagard, B.; Thépault, A.; Lucas, P.; Meunier, M.; Benoit, F.; Wilhem, A.; Gassilloud, B.; Cauvin, E. Comparative Analysis of Campylobacter jejuni and C. coli Isolated from Livestock Animals to C. jejuni and C. coli Isolated from Surface Water Using DNA Sequencing and MALDI-TOF. Pathogens 2023, 12, 1069. [Google Scholar] [CrossRef]
- Zhang, Q.; Beyi, A.F.; Yin, Y. Zoonotic and antibiotic-resistant Campylobacter: A view through the One Health lens. One Health Adv. 2023, 1, 4. [Google Scholar] [CrossRef]
- Kiu, R.; Hall, L.J. An update on the human and animal enteric pathogen Clostridium perfringens. Emerg. Microbes Infect. 2018, 7, 141. [Google Scholar] [CrossRef]
- Simpson, K.M.; Callan, R.J.; Van Metre, D.C. Clostridial abomasitis and enteritis in ruminants. Vet. Clin. Food Anim. Pract. 2018, 34, 155–184. [Google Scholar] [CrossRef] [PubMed]
- García, S.; Heredia, N. Clostridium perfringens: A Dynamic Foodborne Pathogen. Food Bioprocess. Technol. 2011, 4, 624–630. [Google Scholar] [CrossRef]
- Mohiuddin, M.; Iqbal, Z.; Siddique, A.; Liao, S.; Salamat, M.K.F.; Qi, N.; Din, A.M.; Sun, M. Prevalence, genotypic and phenotypic characterization and antibiotic resistance profile of Clostridium perfringens type A and D isolated from feces of sheep (Ovis aries) and goats (Capra hircus) in Punjab, Pakistan. Toxins 2020, 12, 657. [Google Scholar] [CrossRef] [PubMed]
- Hamza, D.; Dorgham, S.M.; Elhariri, M.; Elhelw, R.; Ismael, E. New insight of apparently healthy animals as a potential reservoir for Clostridium perfringens: A public health implication. J. Vet. Res. 2018, 62, 457. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, H.; Elfeky, M.M.; Al-Azeem, A.; Wasel, F.A. Molecular characterization of Clostridium perfringens in small ruminants. SVU-Int. J. Vet. Sci. 2023, 6, 104–123. [Google Scholar] [CrossRef]
- Khan, M.A.; Bahadar, S.; Ullah, N.; Ullah, S.; Shakeeb, U.; Khan, A.Z.; Khan, I.U.; Kalhoro, N.H.; Shah, M.B.; Malik, M.I.U. Distribution and antimicrobial resistance patterns of Clostridium Perfringens isolated from vaccinated and unvaccinated goats. Small Rumin. Res. 2019, 173, 70–73. [Google Scholar] [CrossRef]
- Yadav, J.P.; Kaur, S.; Dhaka, P.; Vijay, D.; Bedi, J.S. Prevalence, molecular characterization, and antimicrobial resistance profile of Clostridium perfringens from India: A scoping review. Anaerobe 2022, 77, 102639. [Google Scholar] [CrossRef]
- Hao, H.; Cheng, G.; Iqbal, Z.; Ai, X.; Hussain, H.I.; Huang, L.; Dai, M.; Wang, Y.; Liu, Z.; Yuan, Z. Benefits and risks of antimicrobial use in food-producing animals. Front. Microbiol. 2014, 5, 288. [Google Scholar] [CrossRef]
- Dudek-Wicher, R.K.; Junka, A.; Bartoszewicz, M. The influence of antibiotics and dietary components on gut microbiota. Gastroenterol. Rev./Przegląd Gastroenterol. 2018, 13, 85–92. [Google Scholar] [CrossRef]
- Huws, S.A.; Creevey, C.J.; Oyama, L.B.; Mizrahi, I.; Denman, S.E.; Popova, M.; Muñoz-Tamayo, R.; Forano, E.; Waters, S.M.; Hess, M. Addressing global ruminant agricultural challenges through understanding the rumen microbiome: Past, present, and future. Front. Microbiol. 2018, 9, 2161. [Google Scholar] [CrossRef]
- Willyard, C. Baby’s first bacteria. Nature 2018, 553, 264–266. [Google Scholar] [CrossRef]
- Li, B.; Zhang, K.; Li, C.; Wang, X.; Chen, Y.; Yang, Y. Characterization and comparison of microbiota in the gastrointestinal tracts of the goat (Capra hircus) during preweaning development. Front. Microbiol. 2019, 10, 2125. [Google Scholar] [CrossRef]
- Zhang, K.; Li, B.; Guo, M.; Liu, G.; Yang, Y.; Wang, X.; Chen, Y.; Zhang, E. Maturation of the goat rumen microbiota involves three stages of microbial colonization. Animals 2019, 9, 1028. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Nagata, R.; Ohkubo, A.; Ohtani, N.; Kushibiki, S.; Ichijo, T.; Sato, S. Changes in ruminal and reticular pH and bacterial communities in Holstein cattle fed a high-grain diet. BMC Vet. Res. 2018, 14, 310. [Google Scholar] [CrossRef]
- Bi, Y.; Cox, M.S.; Zhang, F.; Suen, G.; Zhang, N.; Tu, Y.; Diao, Q. Feeding modes shape the acquisition and structure of the initial gut microbiota in newborn lambs. Environ. Microbiol. 2019, 21, 2333–2346. [Google Scholar] [CrossRef]
- Shabana, I.I.; Albakri, N.N.; Bouqellah, N.A. Metagenomic investigation of faecal microbiota in sheep and goats of the same ages. J. Taibah Univ. Sci. 2021, 15, 1–9. [Google Scholar] [CrossRef]
- He, J.; Yi, L.; Hai, L.; Ming, L.; Gao, W.; Ji, R. Characterizing the bacterial microbiota in different gastrointestinal tract segments of the Bactrian camel. Sci. Rep. 2018, 8, 654. [Google Scholar] [CrossRef]
- Vickers, N.J. Animal communication: When i’m calling you, will you answer too? Curr. Biol. 2017, 27, R713–R715. [Google Scholar] [CrossRef]
- Yeoman, C.J.; Ishaq, S.L.; Bichi, E.; Olivo, S.K.; Lowe, J.; Aldridge, B.M. Biogeographical differences in the influence of maternal microbial sources on the early successional development of the bovine neonatal gastrointestinal tract. Sci. Rep. 2018, 8, 3197. [Google Scholar] [CrossRef]
- Hallstrom, K.; McCormick, B.A. Salmonella interaction with and passage through the intestinal mucosa: Through the lens of the organism. Front. Microbiol. 2011, 2, 88. [Google Scholar] [CrossRef]
- MacDonald, T.T.; Monteleone, G. Immunity, inflammation, and allergy in the gut. Science 2005, 307, 1920–1925. [Google Scholar] [CrossRef]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef]
- Christaki, E.; Bonos, E.; Giannenas, I.; Florou-Paneri, P. Aromatic plants as a source of bioactive compounds. Agriculture 2012, 2, 228–243. [Google Scholar] [CrossRef]
- Caroprese, M.; Ciliberti, M.G.; Albenzio, M. Application of aromatic plants and their extracts in dairy animals. In Feed Additives; Elsevier: Amsterdam, The Netherlands, 2020; pp. 261–277. [Google Scholar]
- Perumalla, A.; Hettiarachchy, N.S.; Ricke, S.C. Current perspectives on probiotics in poultry preharvest food safety. In Direct-Fed Microbials and Prebiotics for Animals: Science and Mechanisms of Action; Springer: New York, NY, USA, 2011; pp. 89–120. [Google Scholar]
- Cobellis, G.; Yu, Z.; Forte, C.; Acuti, G.; Trabalza-Marinucci, M. Dietary supplementation of Rosmarinus officinalis L. leaves in sheep affects the abundance of rumen methanogens and other microbial populations. J. Anim. Sci. Biotechnol. 2016, 7, 27. [Google Scholar] [CrossRef]
- Simitzis, P.E. Enrichment of Animal Diets with Essential Oils—A Great Perspective on Improving Animal Performance and Quality Characteristics of the Derived Products. Medicines 2017, 4, 35. [Google Scholar] [CrossRef]
- Voreades, N.; Kozil, A.; Weir, T.L. Diet and the development of the human intestinal microbiome. Front. Microbiol. 2014, 5, 494. [Google Scholar] [CrossRef]
- Cowan, M.M. Plant products as antimicrobial agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef]
- Irshad, M.; Subhani, M.A.; Ali, S.; Hussain, A. Biological importance of essential oils. Essent. Oils-Oils Nat. 2020, 1, 235–280. [Google Scholar]
- Wilson, R. Aromatherapy: Essential Oils for Vibrant Health and Beauty; Penguin: London, UK, 2002. [Google Scholar]
- Van Wyk, B.-E.; Wink, M. Medicinal Plants of the World; CABI: Wallingford, UK, 2018. [Google Scholar]
- Khamees, A.H. Phytochemical and pharmacological analysis for seeds of two varieties of Iraqi Raphanus sativus. Int. J. Pharm. Sci. Rev. Res. 2017, 43, 237–242. [Google Scholar]
- Celikel, N.; Kavas, G. Antimicrobial properties of some essential oils against some pathogenic microorganisms. Czech J. Food Sci. 2008, 26, 174. [Google Scholar] [CrossRef]
- Holley, R.A.; Patel, D. Improvement in shelf-life and safety of perishable foods by plant essential oils and smoke antimicrobials. Food Microbiol. 2005, 22, 273–292. [Google Scholar] [CrossRef]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Man, A.; Santacroce, L.; Iacob, R.; Mare, A.; Man, L. Antimicrobial Activity of Six Essential Oils Against a Group of Human Pathogens: A Comparative Study. Pathogens 2019, 8, 15. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Adame-Gallegos, J.R.; Andrade-Ochoa, S.; Nevarez-Moorillon, G.V. Potential Use of Mexican Oregano Essential Oil against Parasite, Fungal and Bacterial Pathogens. J. Essent. Oil Bear. Plants 2016, 19, 553–567. [Google Scholar] [CrossRef]
- Dorman, H.D.; Deans, S.G. Antimicrobial agents from plants: Antibacterial activity of plant volatile oils. J. Appl. Microbiol. 2000, 88, 308–316. [Google Scholar] [CrossRef]
- Miguel, M.G. Antioxidant and anti-inflammatory activities of essential oils: A short review. Molecules 2010, 15, 9252–9287. [Google Scholar] [CrossRef]
- Trombetta, D.; Castelli, F.; Sarpietro, M.G.; Venuti, V.; Cristani, M.; Daniele, C.; Saija, A.; Mazzanti, G.; Bisignano, G. Mechanisms of antibacterial action of three monoterpenes. Antimicrob. Agents Chemother. 2005, 49, 2474–2478. [Google Scholar] [CrossRef]
- Sun, J.; Cheng, Z.; Zhao, Y.; Wang, Y.; Wang, H.; Ren, Z. Influence of increasing levels of oregano essential oil on intestinal morphology, intestinal flora and performance of Sewa sheep. Ital. J. Anim. Sci. 2022, 21, 463–472. [Google Scholar] [CrossRef]
- Santos, F.; Rao, V. Antiinflammatory and antinociceptive effects of 1, 8-cineole a terpenoid oxide present in many plant essential oils. Phytother. Res. 2000, 14, 240–244. [Google Scholar] [CrossRef]
- Vasta, V.; Luciano, G. The effects of dietary consumption of plants secondary compounds on small ruminants’ products quality. Small Rumin. Res. 2011, 101, 150–159. [Google Scholar] [CrossRef]
- Lei, Z.; Zhang, K.; Li, C.; Jiao, T.; Wu, J.; Wei, Y.; Tian, K.; Li, C.; Tang, D.; Davis, D.I.; et al. Ruminal metagenomic analyses of goat data reveals potential functional microbiota by supplementation with essential oil-cobalt complexes. BMC Microbiol. 2019, 19, 30. [Google Scholar] [CrossRef]
- Guarda, A.; Rubilar, J.F.; Miltz, J.; Galotto, M.J. The antimicrobial activity of microencapsulated thymol and carvacrol. Int. J. Food Microbiol. 2011, 146, 144–150. [Google Scholar] [CrossRef]
- Bovell-Benjamin, A.C. Sweet potato: A review of its past, present, and future role in human nutrition. Adv. Food Nutr. Res. 2007, 52, 1–59. [Google Scholar]
- Sun, H.; Mu, B.; Song, Z.; Ma, Z.; Mu, T. The in vitro antioxidant activity and inhibition of intracellular reactive oxygen species of sweet potato leaf polyphenols. Oxidative Med. Cell. Longev. 2018, 2018, 9017828. [Google Scholar] [CrossRef]
- Karna, P.; Gundala, S.R.; Gupta, M.V.; Shamsi, S.A.; Pace, R.D.; Yates, C.; Narayan, S.; Aneja, R. Polyphenol-rich sweet potato greens extract inhibits proliferation and induces apoptosis in prostate cancer cells in vitro and in vivo. Carcinogenesis 2011, 32, 1872–1880. [Google Scholar] [CrossRef]
- Islam, S. Sweetpotato (Ipomoea batatas L.) leaf: Its potential effect on human health and nutrition. J. Food Sci. 2006, 71, R13–R121. [Google Scholar] [CrossRef]
- Dewijanti, I.D.; Banjarnahor, S.D.; Triyuliani, T.; Maryani, F.; Meilawati, L. Antioxidant activity, phenolic and flavonoids total of ethanolic extract of Ipomoea batata L. leaves (white, yellow, orange, and purple). In AIP Conference Proceedings; AIP Publishing: Long Island, NY, USA, 2017; p. 020020. [Google Scholar]
- Mbaeyi-Nwaoha, I.E.; Emejulu, V.N. Evaluation of Phytochemical Composition and Antimicrobial Activity of Sweet Potato (Ipomoea batatas) Leaf. Pak. J. Nutr. 2013, 12, 575–586. [Google Scholar] [CrossRef]
- Horton, M.; Robbins, J. The Potential of the Arkansas Sweet Potato Industry: A Matter of Volatility. J. Bus. Adm. Online 2007. [Google Scholar]
- Li, J.; Huang, S.-Y.; Deng, Q.; Li, G.; Su, G.; Liu, J.; Wang, H.-M.D. Extraction and characterization of phenolic compounds with antioxidant and antimicrobial activities from pickled radish. Food Chem. Toxicol. 2020, 136, 111050. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.-W.; Song, K.-Y.; Chon, J.-W.; Jeong, D.; Seo, K.-H. Antimicrobial Action of Raphanus raphanistrum subsp. sativus (radish) Extracts against Foodborne Bacteria Present in Various Milk Products: A Preliminary Study. J. Milk. Sci. Biotechnol. 2019, 37, 187–195. [Google Scholar] [CrossRef]
- Kaymak, H.C.; Yilmaz, S.O.; Ercisli, S.; Guvenc, I. Antibacterial Activities of Red Colored Radish Types (Raphanus sativus L.). Rom. Biotechnol. Lett. 2018, 23, 13744. [Google Scholar]
- Vargas, S.R.; Perez, G.R.; Perez, G.S.; Zavala, S.M.; Perez, G.C. Antiurolithiatic activity of Raphanus sativus aqueous extract on rats. J. Ethnopharmacol. 1999, 68, 335–338. [Google Scholar] [CrossRef] [PubMed]
- Ahn, K.; Ji, H.; Kim, H.-E.; Cho, H.; Sun, Q.; Shi, S.; He, Y.; Kim, B.-G.; Kim, O. Raphanus sativus L. Seed Extracts Induce Apoptosis and Reduce Migration of Oral Squamous Cell Carcinoma KB and KBCD133+ Cells by Downregulation of β-Catenin; International System for Agricultural Science and Technology: Rome, Italy, 2020. [Google Scholar]
- Kang, H.; Lee, S.; Eom, H.; Yu, J.; Lee, S. Anti-inflammatory and antitumor phenylpropanoid sucrosides from the seeds of Raphanus sativus. Planta Medica 2015, 81, PX12. [Google Scholar] [CrossRef]
- Gutiérrez, R.M.; Perez, R.L. Raphanus sativus (Radish): Their Chemistry and Biology. Sci. World J. 2004, 4, 811–837. [Google Scholar] [CrossRef]
- Choi, K.-C.; Cho, S.-W.; Kook, S.-H.; Chun, S.-R.; Bhattarai, G.; Poudel, S.B.; Kim, M.-K.; Lee, K.-Y.; Lee, J.-C. Intestinal anti-inflammatory activity of the seeds of Raphanus sativus L. in experimental ulcerative colitis models. J. Ethnopharmacol. 2016, 179, 55–65. [Google Scholar] [CrossRef]
- Roberts, T.; Ortel, C.; Hoegenauer, K.; Wright, H.; Durre, T. Understanding Cover Crops; University of Arkansas: Fayetteville, AR, USA, 2018. [Google Scholar]
- Henderson, A.J.; Ollila, C.A.; Kumar, A.; Borresen, E.C.; Raina, K.; Agarwal, R.; Ryan, E.P. Chemopreventive properties of dietary rice bran: Current status and future prospects. Adv. Nutr. 2012, 3, 643–653. [Google Scholar] [CrossRef]
- Ravichanthiran, K.; Ma, Z.F.; Zhang, H.; Cao, Y.; Wang, C.W.; Muhammad, S.; Aglago, E.K.; Zhang, Y.; Jin, Y.; Pan, B. Phytochemical profile of brown rice and its nutrigenomic implications. Antioxidants 2018, 7, 71. [Google Scholar] [CrossRef]
- Komiyama, Y.; Andoh, A.; Fujiwara, D.; Ohmae, H.; Araki, Y.; Fujiyama, Y.; Mitsuyama, K.; Kanauchi, O. New prebiotics from rice bran ameliorate inflammation in murine colitis models through the modulation of intestinal homeostasis and the mucosal immune system. Scand. J. Gastroenterol. 2011, 46, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.; Savary, B.J.; Teoh, K.; Chen, M.-H.; McClung, A.; Lee, S.-O. In vitro fermentation patterns of rice bran components by human gut microbiota. Nutrients 2017, 9, 1237. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Chen, H.; Gao, Y.; An, N.; Li, X.; Pan, X.; Yang, X.; Tian, L.; Sun, J.; Xiong, X. Gut microbiota-derived short-chain fatty acids and hypertension: Mechanism and treatment. Biomed. Pharmacother. 2020, 130, 110503. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Henderson, A.; Forster, G.M.; Goodyear, A.W.; Weir, T.L.; Leach, J.E.; Dow, S.W.; Ryan, E.P. Dietary rice bran promotes resistance to Salmonella enterica serovar Typhimurium colonization in mice. BMC Microbiol. 2012, 12, 71. [Google Scholar] [CrossRef] [PubMed]
- Herfel, T.; Jacobi, S.; Lin, X.; Van Heugten, E.; Fellner, V.; Odle, J. Stabilized rice bran improves weaning pig performance via a prebiotic mechanism. J. Anim. Sci. 2013, 91, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Bodie, A.R.; Micciche, A.C.; Atungulu, G.G.; Rothrock Jr, M.J.; Ricke, S.C. Current trends of rice milling byproducts for agricultural applications and alternative food production systems. Front. Sustain. Food Syst. 2019, 3, 47. [Google Scholar] [CrossRef]
- Rubinelli, P.M.; Kim, S.A.; Park, S.H.; Roto, S.M.; Nealon, N.J.; Ryan, E.P.; Ricke, S.C. Differential effects of rice bran cultivars to limit Salmonella Typhimurium in chicken cecal in vitro incubations and impact on the cecal microbiome and metabolome. PLoS ONE 2017, 12, e0185002. [Google Scholar] [CrossRef]
- Moldenhauer, K.; Sha, X.; Berger, G.; Hardke, J.; Norman, R.; Wilson Jr, C.; Wamishe, Y.; Cartwright, R.; Blocker, M.; McCarty, D. LaKast, a high yielding, very short season, long-grain rice variety. BR Wells Rice Res. Stud.-Ark. Agric. Exp. Stn. Univ. Ark. Syst. 2014, 617, 74–80. [Google Scholar]
- Kim, S.A.; Rubinelli, P.M.; Park, S.H.; Ricke, S.C. Ability of Arkansas LaKast and LaKast Hybrid Rice Bran to Reduce Salmonella Typhimurium in Chicken Cecal Incubations and Effects on Cecal Microbiota. Front. Microbiol. 2018, 9, 134. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daniel, I.K.; Njue, O.M.; Sanad, Y.M. Antimicrobial Effects of Plant-Based Supplements on Gut Microbial Diversity in Small Ruminants. Pathogens 2024, 13, 31. https://doi.org/10.3390/pathogens13010031
Daniel IK, Njue OM, Sanad YM. Antimicrobial Effects of Plant-Based Supplements on Gut Microbial Diversity in Small Ruminants. Pathogens. 2024; 13(1):31. https://doi.org/10.3390/pathogens13010031
Chicago/Turabian StyleDaniel, Ian K., Obadiah M. Njue, and Yasser M. Sanad. 2024. "Antimicrobial Effects of Plant-Based Supplements on Gut Microbial Diversity in Small Ruminants" Pathogens 13, no. 1: 31. https://doi.org/10.3390/pathogens13010031
APA StyleDaniel, I. K., Njue, O. M., & Sanad, Y. M. (2024). Antimicrobial Effects of Plant-Based Supplements on Gut Microbial Diversity in Small Ruminants. Pathogens, 13(1), 31. https://doi.org/10.3390/pathogens13010031