
Complement Suppresses the Initial Type 1 Interferon Response to Ocular Herpes Simplex Virus Type 1 Infection in Mice



Abstract
:1. Introduction
2. Results
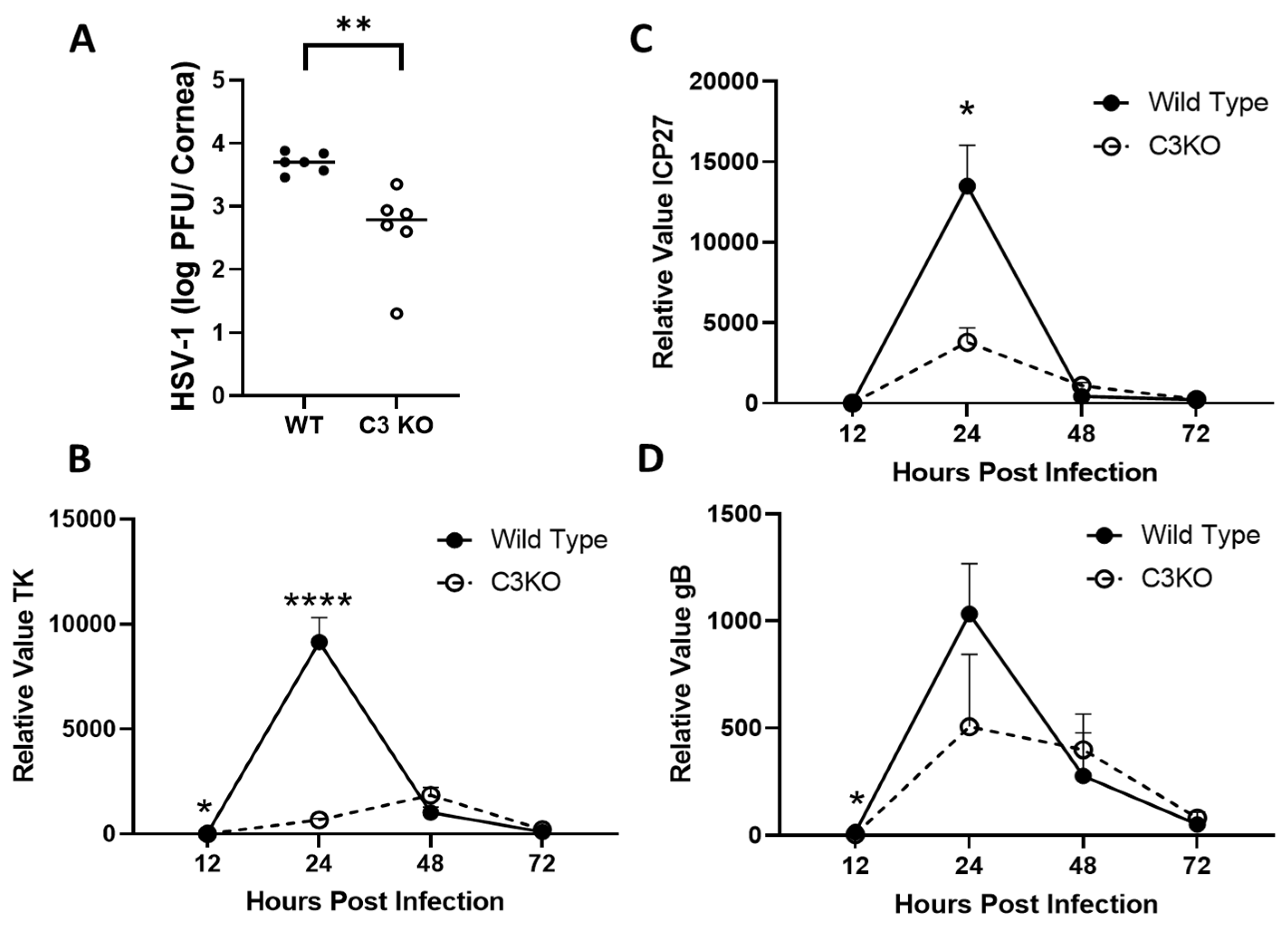
2.1. C3KO Mice Possess Less Infectious Virus and Express Lower Levels of HSV-1 Lytic Genes Compared to Wild-Type Mice 12–24 h Post Infection
2.2. C3 Levels in the Cornea following HSV-1 Infection Peak at 12 h PI
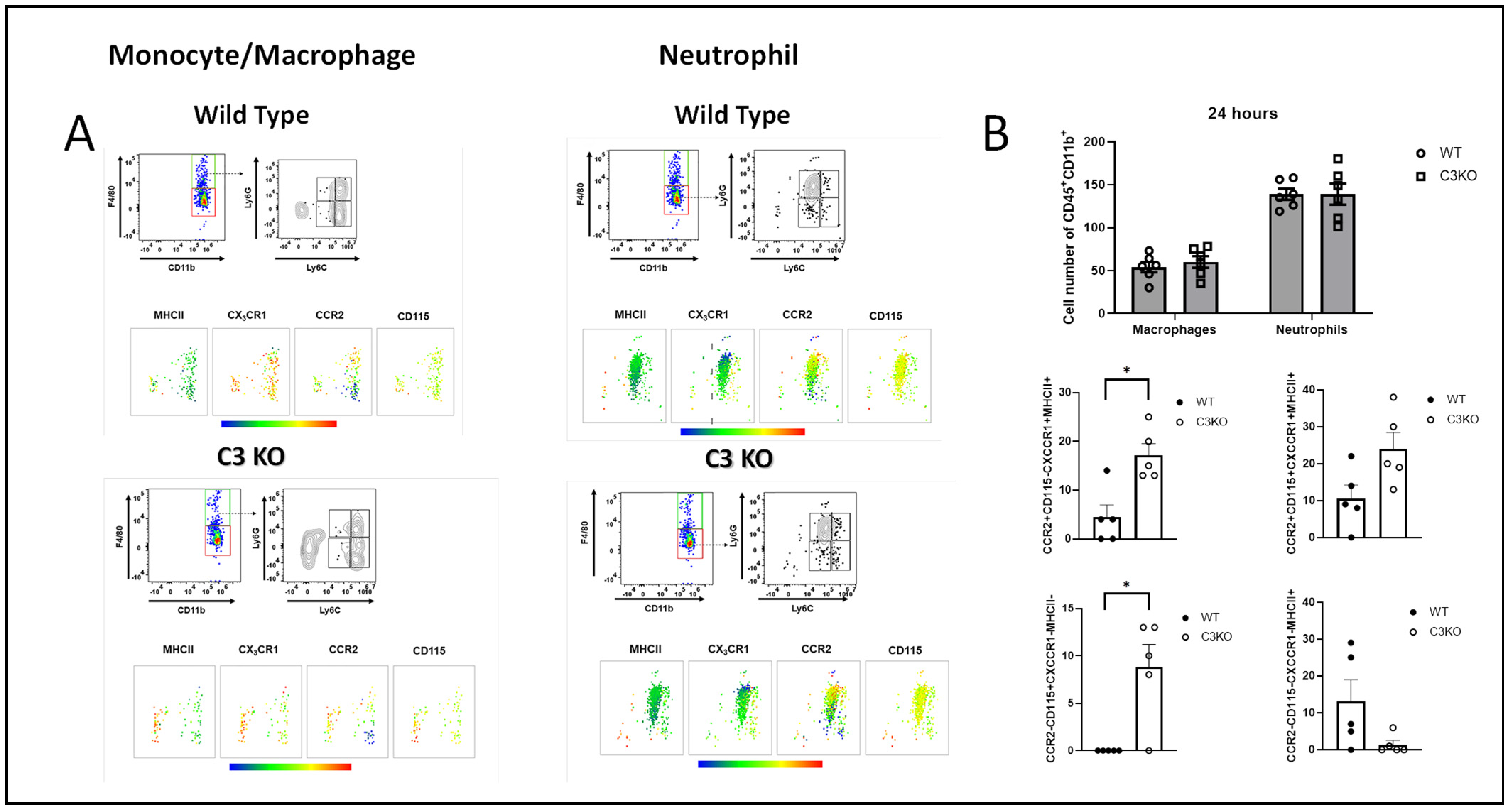
2.3. Myeloid Cell Infiltrate into the Cornea Is Similar between WT and C3KO Mice Following HSV-1 Infection
2.4. CCL3/Macrophage Inflammatory Protein (MIP)-1α Is Elevated in the Cornea of WT Mice 12 h PI
2.5. IFN-β and Downstream Effector Molecule RNase L Are Elevated in the Cornea of C3KO Mice Early Post Infection
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Cells, Virus, and Viral Plaque Assay
4.3. Ocular Infection
4.4. C3 ELISA and Cytokine/Chemokine Multiplex Suspension Array
4.5. Real-Time Reverse Transcriptase (RT)-Polymerase Chain Reaction (PCR)
4.6. Flow Cytometry
4.7. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nonaka, M.; Kimura, A. Genomic view of the evolution of the complement system. Immunogenetics 2006, 58, 701–713. [Google Scholar] [CrossRef] [PubMed]
- Ricklin, D.; Hajishengallis, G.; Yang, K.; Lambris, J.D. Complement: A key system for immune surveillance and homeostasis. Nat. Immunol. 2010, 11, 785–797. [Google Scholar] [CrossRef]
- Bubeck, D. The making of a macromolecular machine: Assembly of the membrane attack complex. Biochemistry 2014, 53, 1908–1915. [Google Scholar] [CrossRef]
- Tam, J.C.; Bidgood, S.R.; McEwan, W.A.; James, L.C. Intracellular sensing of complement C3 activates cell autonomous immunity. Science 2014, 345, 1256070. [Google Scholar] [CrossRef] [PubMed]
- Liszewski, M.K.; Elvington, M.; Kulkarni, H.S.; Atkinson, J.P. Complement’s hidden arsenal: New insights and novel functions inside the cell. Mol. Immunol. 2017, 84, 2–9. [Google Scholar] [CrossRef]
- Bennett, K.M.; Rooijakkers, S.H.; Gorham, R.D., Jr. Let’s Tie the Knot: Marriage of Complement and Adaptive Immunity in Pathogen Evasion, for Better or Worse. Front. Microbiol. 2017, 8, 89. [Google Scholar] [CrossRef] [PubMed]
- Mondino, B.J.; Hoffman, D.B. Hemolytic complement activity in normal human donor corneas. Arch. Ophthalmol. 1980, 98, 2041–2044. [Google Scholar] [CrossRef] [PubMed]
- Rothman, B.; Despins, A.; Webb, S.; Taylor, D.; Sundarraj, N.; O’Rourke, J.; Kreutzer, D. Cytokine regulation of C3 and C5 production by human corneal fibroblasts. Exp. Eye Res. 1991, 53, 353–361. [Google Scholar] [CrossRef]
- Mondino, B.J.; Chou, H.J.; Sumner, H.L. Generation of complement membrane attack complex in normal human corneas. Investig. Ophthalmol. Vis. Sci. 1996, 37, 1576–1581. [Google Scholar]
- Montalvo, V.; Chan, C.-C.; Gery, I.; Campos, M.M.; Wawrousek, E.F.; Bush, R.A.; Lambris, J.D. Complement deposits on ocular tissues adjacent to sites of inflammation. Curr. Eye Res. 2007, 32, 917–922. [Google Scholar] [CrossRef]
- Mohammed, M.R.S.; Krishnan, S.; Amrathlal, R.S.; Jayapal, J.M.; Namperumalsamy, V.P.; Prajna, L.; Kuppamuthu, D. Local Activation of the Alternative Pathway of Complement System in Mycotic Keratitis Patient Tear. Front. Cell. Infect. Microbiol. 2020, 10, 205. [Google Scholar] [CrossRef] [PubMed]
- Bora, N.S.; Gobleman, C.L.; Atkinson, J.P.; Pepose, J.S.; Kaplan, H.J. Differential expression of the complement regulatory proteins in the human eye. Investig. Ophthalmol. Vis. Sci. 1993, 34, 3579–3584. [Google Scholar]
- Clark, S.J.; Bishop, P.N. The eye as a complement dysregulation hotspot. Semin. Immunopathol. 2018, 40, 65–74. [Google Scholar] [CrossRef]
- McCormick, I.; James, C.; Welton, N.J.; Mayaud, P.; Turner, K.M.E.; Gottlieb, S.L.; Foster, A.; Looker, K.J. Incidence of Herpes Simplex Virus Keratitis and Other Ocular Disease: Global Review and Estimates. Ophthalmic Epidemiol. 2022, 29, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Summers, B.C.; Margolis, T.P.; Leib, D.A. Herpes simplex virus type 1 corneal infection results in periocular disease by zosteriform spread. J. Virol. 2001, 75, 5069–5075. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.; Gangappa, S.; Kanangat, S.; Rouse, B.T. On the essential involvement of neutrophils in the immunopathologic disease: Herpetic stromal keratitis. J. Immunol. 1997, 158, 1383–1391. [Google Scholar] [CrossRef] [PubMed]
- Gaddipati, S.; Rao, P.; Jerome, A.D.; Burugula, B.B.; Gerard, N.P.; Suvas, S. Loss of Neurokinin-1 Receptor Alters Ocular Surface Homeostasis and Promotes an Early Development of Herpes Stromal Keratitis. J. Immunol. 2016, 197, 4021–4033. [Google Scholar] [CrossRef]
- Cheng, H.; Tumpey, T.M.; Staats, H.F.; van Rooijen, N.; Oakes, J.E.; Lausch, R.N. Role of macrophages in restricting herpes simplex virus type 1 growth after ocular infection. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1402–1409. [Google Scholar]
- Conrady, C.D.; Zheng, M.; Mandal, N.A.; van Rooijen, N.; Carr, D.J. IFN-alpha-driven CCL2 production recruits inflammatory monocytes to infection site in mice. Mucosal Immunol. 2013, 6, 45–55. [Google Scholar] [CrossRef]
- Jeon, S.; Rowe, A.M.; Carroll, K.L.; Harvey, S.A.K.; Hendricks, R.L. PD-L1/B7-H1 Inhibits Viral Clearance by Macrophages in HSV-1-Infected Corneas. J. Immunol. 2018, 200, 3711–3719. [Google Scholar] [CrossRef]
- Frank, G.M.; Buela, K.A.; Maker, D.M.; Harvey, S.A.; Hendricks, R.L. Early responding dendritic cells direct the local NK response to control herpes simplex virus 1 infection within the cornea. J. Immunol. 2012, 188, 1350–1359. [Google Scholar] [CrossRef]
- Royer, D.J.; Zheng, M.; Conrady, C.D.; Carr, D.J. Granulocytes in Ocular HSV-1 Infection: Opposing Roles of Mast Cells and Neutrophils. Investig. Ophthalmol. Vis. Sci. 2015, 56, 3763–3775. [Google Scholar] [CrossRef] [PubMed]
- Hendricks, R.L.; Weber, P.C.; Taylor, J.L.; Koumbis, A.; Tumpey, T.M.; Glorioso, J.C. Endogenously produced interferon alpha protects mice from herpes simplex virus type 1 corneal disease. J. Gen. Virol. 1991, 72 Pt 7, 1601–1610. [Google Scholar] [CrossRef]
- Carr, D.J.; Veress, L.A.; Noisakran, S.; Campbell, I.L. Astrocyte-targeted expression of IFN-alpha1 protects mice from acute ocular herpes simplex virus type 1 infection. J. Immunol. 1998, 161, 4859–4865. [Google Scholar] [CrossRef] [PubMed]
- Antony, F.; Pundkar, C.; Sandey, M.; Jaiswal, A.K.; Mishra, A.; Kumar, A.; Channappanavar, R.; Suryawanshi, A. IFN-λ Regulates Neutrophil Biology to Suppress Inflammation in Herpes Simplex Virus-1-Induced Corneal Immunopathology. J. Immunol. 2021, 206, 1866–1877. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.H.; Yan, X.T.; Oakes, J.E.; Lausch, R.N. Protective antibody therapy is associated with reduced chemokine transcripts in herpes simplex virus type 1 corneal infection. J. Virol. 1996, 70, 1277–1281. [Google Scholar] [CrossRef] [PubMed]
- Carr, D.J.; Tomanek, L. Herpes simplex virus and the chemokines that mediate the inflammation. Curr. Top. Microbiol. Immunol. 2006, 303, 47–65. [Google Scholar]
- Tumpey, T.M.; Cheng, H.; Cook, D.N.; Smithies, O.; Oakes, J.E.; Lausch, R.N. Absence of macrophage inflammatory protein-1alpha prevents the development of blinding herpes stromal keratitis. J. Virol. 1998, 72, 3705–3710. [Google Scholar] [CrossRef]
- Yun, H.; Rowe, A.M.; Lathrop, K.L.; Harvey, S.A.; Hendricks, R.L. Reversible nerve damage and corneal pathology in murine herpes simplex stromal keratitis. J. Virol. 2014, 88, 7870–7880. [Google Scholar] [CrossRef]
- Chucair-Elliott, A.J.; Jinkins, J.; Carr, M.M.; Carr, D.J. IL-6 Contributes to Corneal Nerve Degeneration after Herpes Simplex Virus Type I Infection. Am. J. Pathol. 2016, 186, 2665–2678. [Google Scholar] [CrossRef]
- Chucair-Elliott, A.J.; Gurung, H.R.; Carr, M.M.; Carr, D.J.J. Colony Stimulating Factor-1 Receptor Expressing Cells Infiltrating the Cornea Control Corneal Nerve Degeneration in Response to HSV-1 Infection. Investig. Ophthalmol. Vis. Sci. 2017, 58, 4670–4682. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Antony, F.; Rouse, B.T.; Suryawanshi, A. Role of Innate Interferon Responses at the Ocular Surface in Herpes Simplex Virus-1-Induced Herpetic Stromal Keratitis. Pathogens 2023, 12, 437. [Google Scholar] [CrossRef] [PubMed]
- Hook, L.M.; Lubinski, J.M.; Jiang, M.; Pangburn, M.K.; Friedman, H.M. Herpes simplex virus type 1 and 2 glycoprotein C prevents complement-mediated neutralization induced by natural immunoglobulin M antibody. J. Virol. 2006, 80, 4038–4046. [Google Scholar] [CrossRef] [PubMed]
- Friedman, H.M.; Wang, L.; Fishman, N.O.; Lambris, J.D.; Eisenberg, R.J.; Cohen, G.H.; Lubinski, J. Immune evasion properties of herpes simplex virus type 1 glycoprotein gC. J. Virol. 1996, 70, 4253–4260. [Google Scholar] [CrossRef] [PubMed]
- Lubinski, J.M.; Wang, L.; Soulika, A.M.; Burger, R.; Wetsel, R.A.; Colten, H.; Cohen, G.H.; Eisenberg, R.J.; Lambris, J.D.; Friedman, H.M. Herpes simplex virus type 1 glycoprotein gC mediates immune evasion in vivo. J. Virol. 1998, 72, 8257–8263. [Google Scholar] [CrossRef]
- Da Costa, X.J.; Brockman, M.A.; Alicot, E.; Ma, M.; Fischer, M.B.; Zhou, X.; Knipe, D.M.; Carroll, M.C. Humoral response to herpes simplex virus is complement-dependent. Proc. Natl. Acad. Sci. USA 1999, 96, 12708–12712. [Google Scholar] [CrossRef] [PubMed]
- Royer, D.J.; Echegaray-Mendez, J.; Lin, L.; Gmyrek, G.B.; Mathew, R.; Saban, D.R.; Perez, V.L.; Carr, D.J.J. Complement and CD4(+) T cells drive context-specific corneal sensory neuropathy. Elife 2019, 8, e48378. [Google Scholar] [CrossRef]
- Filiberti, A.; Gmyrek, G.B.; Berube, A.N.; Royer, D.J.; Carr, D.J.J. An intact complement system dampens cornea inflammation during acute primary HSV-1 infection. Sci. Rep. 2021, 11, 10247. [Google Scholar] [CrossRef]
- Secombes, C.J.; Zou, J. Evolution of Interferons and Interferon Receptors. Front. Immunol. 2017, 8, 209. [Google Scholar] [CrossRef]
- Fattahi, F.; Grailer, J.J.; Parlett, M.; Lu, H.; Malan, E.A.; Abe, E.; Russell, M.W.; Frydrych, L.M.; Delano, M.J.; Zetoune, F.S.; et al. Requirement of Complement C6 for Intact Innate Immune Responses in Mice. J. Immunol. 2020, 205, 251–260. [Google Scholar] [CrossRef]
- Vollstedt, S.; Franchini, M.; Alber, G.; Ackermann, M.; Suter, M. Interleukin-12- and gamma interferon-dependent innate immunity are essential and sufficient for long-term survival of passively immunized mice infected with herpes simplex virus type 1. J. Virol. 2001, 75, 9596–9600. [Google Scholar] [CrossRef] [PubMed]
- Rossol-Voth, R.; Rossol, S.; Schütt, K.H.; Corridori, S.; de Cian, W.; Falke, D. In vivo protective effect of tumour necrosis factor alpha against experimental infection with herpes simplex virus type 1. J. Gen. Virol. 1991, 72 Pt 1, 143–147. [Google Scholar] [CrossRef]
- Sergerie, Y.; Rivest, S.; Boivin, G. Tumor necrosis factor-alpha and interleukin-1 beta play a critical role in the resistance against lethal herpes simplex virus encephalitis. J. Infect. Dis. 2007, 196, 853–860. [Google Scholar] [CrossRef]
- Halford, W.P.; Veress, L.A.; Gebhardt, B.M.; Carr, D.J. Immunization with HSV-1 antigen rapidly protects against HSV-1-induced encephalitis and is IFN-gamma independent. J. Interferon Cytokine Res. 1998, 18, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Härle, P.; Sainz, B.; Carr, D.J., Jr.; Halford, W.P. The immediate-early protein, ICP0, is essential for the resistance of herpes simplex virus to interferon-alpha/beta. Virology 2002, 293, 295–304. [Google Scholar] [CrossRef]
- Melroe, G.T.; DeLuca, N.A.; Knipe, D.M. Herpes simplex virus 1 has multiple mechanisms for blocking virus-induced interferon production. J. Virol. 2004, 78, 8411–8420. [Google Scholar] [CrossRef] [PubMed]
- Christensen, M.H.; Jensen, S.B.; Miettinen, J.J.; Luecke, S.; Prabakaran, T.; Reinert, L.S.; Mettenleiter, T.; Chen, Z.J.; Knipe, D.M.; Sandri-Goldin, R.M.; et al. HSV-1 ICP27 targets the TBK1-activated STING signalsome to inhibit virus-induced type I IFN expression. Embo J. 2016, 35, 1385–1399. [Google Scholar] [CrossRef]
- Verpooten, D.; Ma, Y.; Hou, S.; Yan, Z.; He, B. Control of TANK-binding kinase 1-mediated signaling by the gamma(1)34.5 protein of herpes simplex virus 1. J. Biol. Chem. 2009, 284, 1097–1105. [Google Scholar] [CrossRef]
- Xing, J.; Ni, L.; Wang, S.; Wang, K.; Lin, R.; Zheng, C. Herpes simplex virus 1-encoded tegument protein VP16 abrogates the production of beta interferon (IFN) by inhibiting NF-κB activation and blocking IFN regulatory factor 3 to recruit its coactivator CBP. J. Virol. 2013, 87, 9788–9801. [Google Scholar] [CrossRef]
- Meurs, E.F.; Watanabe, Y.; Kadereit, S.; Barber, G.N.; Katze, M.G.; Chong, K.; Williams, B.R.; Hovanessian, A.G. Constitutive expression of human double-stranded RNA-activated p68 kinase in murine cells mediates phosphorylation of eukaryotic initiation factor 2 and partial resistance to encephalomyocarditis virus growth. J. Virol. 1992, 66, 5805–5814. [Google Scholar] [CrossRef]
- He, B.; Gross, M.; Roizman, B. The gamma(1)34.5 protein of herpes simplex virus 1 complexes with protein phosphatase 1alpha to dephosphorylate the alpha subunit of the eukaryotic translation initiation factor 2 and preclude the shutoff of protein synthesis by double-stranded RNA-activated protein kinase. Proc. Natl. Acad. Sci. USA 1997, 94, 843–848. [Google Scholar] [PubMed]
- Castelli, J.C.; Hassel, B.A.; Wood, K.A.; Li, X.-L.; Amemiya, K.; Dalakas, M.C.; Torrence, P.F.; Youle, R.J. A study of the interferon antiviral mechanism: Apoptosis activation by the 2-5A system. J. Exp. Med. 1997, 186, 967–972. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.; Paranjape, J.; Brown, T.L.; Nie, H.; Naik, S.; Dong, B.; Chang, A.; Trapp, B.; Fairchild, R.; Colmenares, C.; et al. Interferon action apoptosis are defective in mice devoid of 2′,5′-oligoadenylate-dependent RNase L. Embo J. 1997, 16, 6355–6363. [Google Scholar] [CrossRef] [PubMed]
- Royer, D.J.; Carr, D.J. A STING-dependent innate-sensing pathway mediates resistance to corneal HSV-1 infection via upregulation of the antiviral effector tetherin. Mucosal Immunol. 2016, 9, 1065–1075. [Google Scholar] [CrossRef]
- Sharma, N.; Wang, C.; Kessler, P.; Sen, G.C. Herpes simplex virus 1 evades cellular antiviral response by inducing microRNA-24, which attenuates STING synthesis. PLoS Pathog. 2021, 17, e1009950. [Google Scholar] [CrossRef]
- Lebel, M.; Langlois, M.-P.; Daudelin, J.-F.; Tarrab, E.; Savard, P.; Leclerc, D.; Lamarre, A. Complement Component 3 Regulates IFN-α Production by Plasmacytoid Dendritic Cells following TLR7 Activation by a Plant Virus-like Nanoparticle. J. Immunol. 2017, 198, 292–299. [Google Scholar] [CrossRef]
- Fung, K.Y.; Mangan, N.E.; Cumming, H.; Horvat, J.C.; Mayall, J.R.; Stifter, S.A.; De Weerd, N.; Roisman, L.C.; Rossjohn, J.; Robertson, S.A.; et al. Interferon-ε protects the female reproductive tract from viral and bacterial infection. Science 2013, 339, 1088–1092. [Google Scholar] [CrossRef]
- Härle, P.; Cull, V.; Agbaga, M.P.; Silverman, R.; Williams, B.R.; James, C.; Carr, D.J.J. Differential effect of murine alpha/beta interferon transgenes on antagonization of herpes simplex virus type 1 replication. J. Virol. 2002, 76, 6558–6567. [Google Scholar] [CrossRef]
- Härle, P.; Cull, V.; Guo, L.; Papin, J.; Lawson, C.; Carr, D.J. Transient transfection of mouse fibroblasts with type I interferon transgenes provides various degrees of protection against herpes simplex virus infection. Antivir. Res. 2002, 56, 39–49. [Google Scholar] [CrossRef]
- Stark, G.R.; Kerr, I.M.; Williams, B.R.; Silverman, R.H.; Schreiber, R.D. How cells respond to interferons. Annu. Rev. Biochem. 1998, 67, 227–264. [Google Scholar] [CrossRef]
- Al-Khatib, K.; Williams, B.R.; Silverman, R.H.; Halford, W.; Carr, D.J. Distinctive roles for 2’,5’-oligoadenylate synthetases and double-stranded RNA-dependent protein kinase R in the in vivo antiviral effect of an adenoviral vector expressing murine IFN-beta. J. Immunol. 2004, 172, 5638–5647. [Google Scholar] [CrossRef] [PubMed]
- Austin, B.A.; James, C.; Silverman, R.H.; Carr, D.J. Critical role for the oligoadenylate synthetase/RNase L pathway in response to IFN-beta during acute ocular herpes simplex virus type 1 infection. J. Immunol. 2005, 175, 1100–1106. [Google Scholar] [CrossRef]
- Smith, S.E.; Busse, D.C.; Binter, S.; Weston, S.; Soria, C.D.; Laksono, B.M.; Clare, S.; Van Nieuwkoop, S.; Hoogen, B.G.V.D.; Clement, M.; et al. Interferon-Induced Transmembrane Protein 1 Restricts Replication of Viruses That Enter Cells via the Plasma Membrane. J. Virol. 2019, 93, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Lenschow, D.J.; Lai, C.; Frias-Staheli, N.; Giannakopoulos, N.V.; Lutz, A.; Wolff, T.; Osiak, A.; Levine, B.; Schmidt, R.E.; García-Sastre, A.; et al. IFN-stimulated gene 15 functions as a critical antiviral molecule against influenza, herpes, and Sindbis viruses. Proc. Natl. Acad. Sci. USA 2007, 104, 1371–1376. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.-Y.; Gowripalan, A.; Croft, S.N.; Smith, S.A.; Helbig, K.J.; Man, S.M.; Tscharke, D.C. Viperin has species-specific roles in response to herpes simplex virus infection. J. Gen. Virol. 2021, 102, 001638. [Google Scholar] [CrossRef]
- Hendricks, R.L.; Tumpey, T.M.; Finnegan, A. IFN-gamma and IL-2 are protective in the skin but pathologic in the corneas of HSV-1-infected mice. J. Immunol. 1992, 149, 3023–3028. [Google Scholar] [CrossRef]
- Rowe, A.M.; St Leger, A.J.; Jeon, S.; Dhaliwal, D.K.; Knickelbein, J.E.; Hendricks, R.L. Herpes keratitis. Prog. Retin. Eye Res. 2013, 32, 88–101. [Google Scholar] [CrossRef]
- Bryant-Hudson, K.; Conrady, C.D.; Carr, D.J. Type I interferon and lymphangiogenesis in the HSV-1 infected cornea—Are they beneficial to the host? Prog. Retin. Eye Res. 2013, 36, 281–291. [Google Scholar] [CrossRef]
- Jha, P.; Sohn, J.-H.; Xu, Q.; Nishihori, H.; Wang, Y.; Nishihori, S.; Manickam, B.; Kaplan, H.J.; Bora, P.S.; Bora, N.S. The complement system plays a critical role in the development of experimental autoimmune anterior uveitis. Investig. Ophthalmol. Vis. Sci. 2006, 47, 1030–1038. [Google Scholar] [CrossRef]
- Bora, N.S.; Jha, P.; Bora, P.S. The role of complement in ocular pathology. Semin. Immunopathol. 2008, 30, 85–95. [Google Scholar] [CrossRef]
- Stuart, P.M.; Morris, J.E.; Sidhu, M.; Keadle, T.L. CCL3 protects mice from corneal pathology during recurrent HSV-1 infection. Front. Biosci. 2008, 13, 4407–4415. [Google Scholar] [CrossRef] [PubMed]
- Berube, A.; Gmyrek, G.B.; Royer, D.J.; Carr, D.J.J. Tripartite-Motif 21 (TRIM21) Deficiency Results in a Modest Loss of Herpes Simplex Virus (HSV)-1 Surveillance in the Trigeminal Ganglia Following Cornea Infection. Viruses 2022, 14, 589. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| ICP27 | 5′-GCA TCC TTC GTG TTT GTC AT-3′ | 5′-ACC AAG GGT CGC GTA GTC-3′ |
| TK | 5′ATA CCG ACG ATC TGC GAC CT-3′ | 5′-TTA TTG CCG TCA TAG CGC GG-3′ |
| gB | 5′-TCT GCA CCA TGA CCA AGT G-3′ | 5′-TGG TGA AGG TCC TCC ATA TG-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carr, D.J.J.; Filiberti, A.; Gmyrek, G.B. Complement Suppresses the Initial Type 1 Interferon Response to Ocular Herpes Simplex Virus Type 1 Infection in Mice. Pathogens 2024, 13, 74. https://doi.org/10.3390/pathogens13010074
Carr DJJ, Filiberti A, Gmyrek GB. Complement Suppresses the Initial Type 1 Interferon Response to Ocular Herpes Simplex Virus Type 1 Infection in Mice. Pathogens. 2024; 13(1):74. https://doi.org/10.3390/pathogens13010074
Chicago/Turabian StyleCarr, Daniel J. J., Adrian Filiberti, and Grzegorz B. Gmyrek. 2024. "Complement Suppresses the Initial Type 1 Interferon Response to Ocular Herpes Simplex Virus Type 1 Infection in Mice" Pathogens 13, no. 1: 74. https://doi.org/10.3390/pathogens13010074
APA StyleCarr, D. J. J., Filiberti, A., & Gmyrek, G. B. (2024). Complement Suppresses the Initial Type 1 Interferon Response to Ocular Herpes Simplex Virus Type 1 Infection in Mice. Pathogens, 13(1), 74. https://doi.org/10.3390/pathogens13010074