
Mycoplasma gallisepticum and Mycoplasma synoviae in Turkeys in Poland

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extraction
2.3. Real-Time PCR
2.4. PCR and Sequence Analysis
2.5. Presence of Other Bacterial Pathogens
2.6. Statistical Analysis
3. Results
3.1. Isolation and Identification
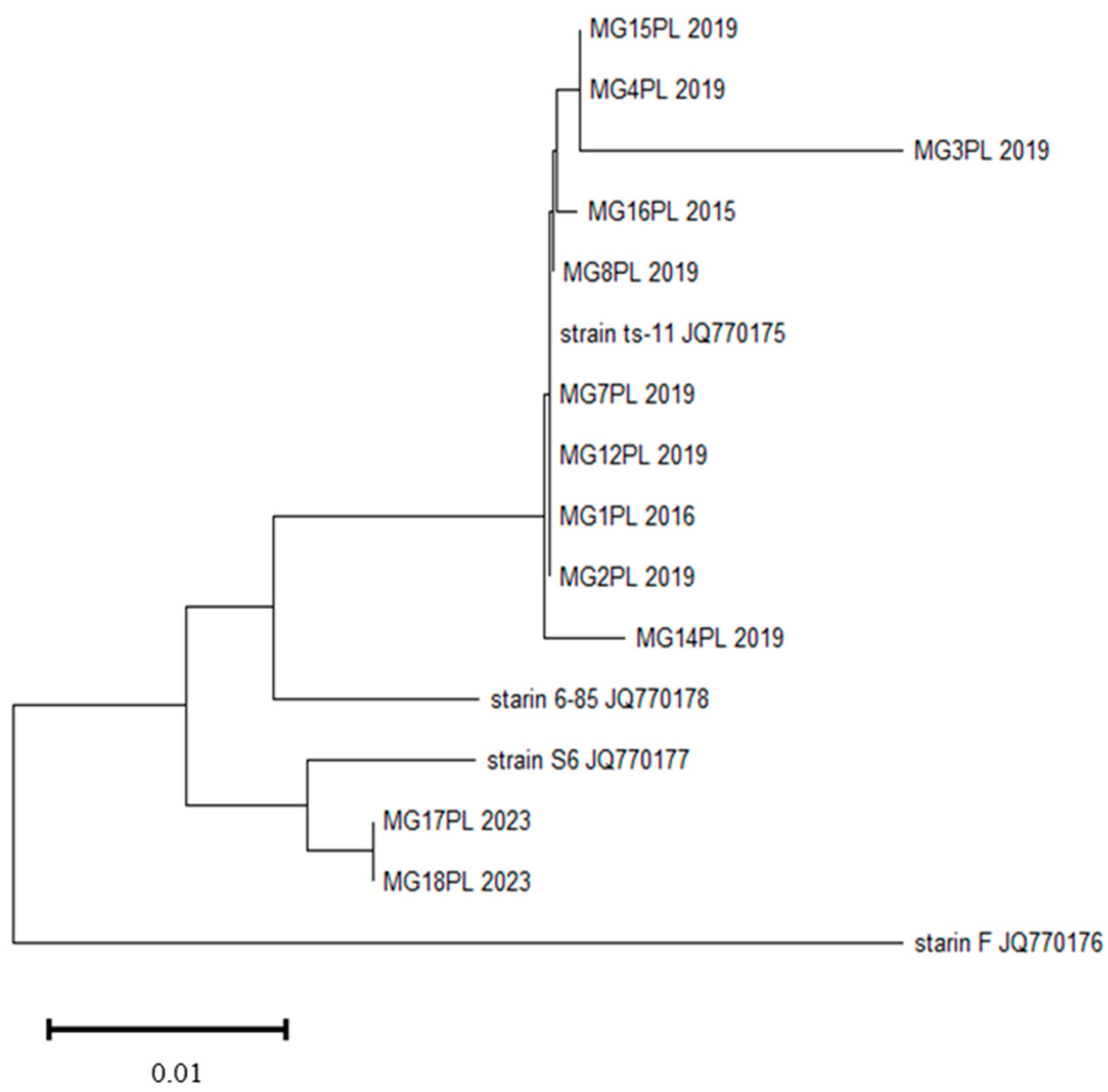
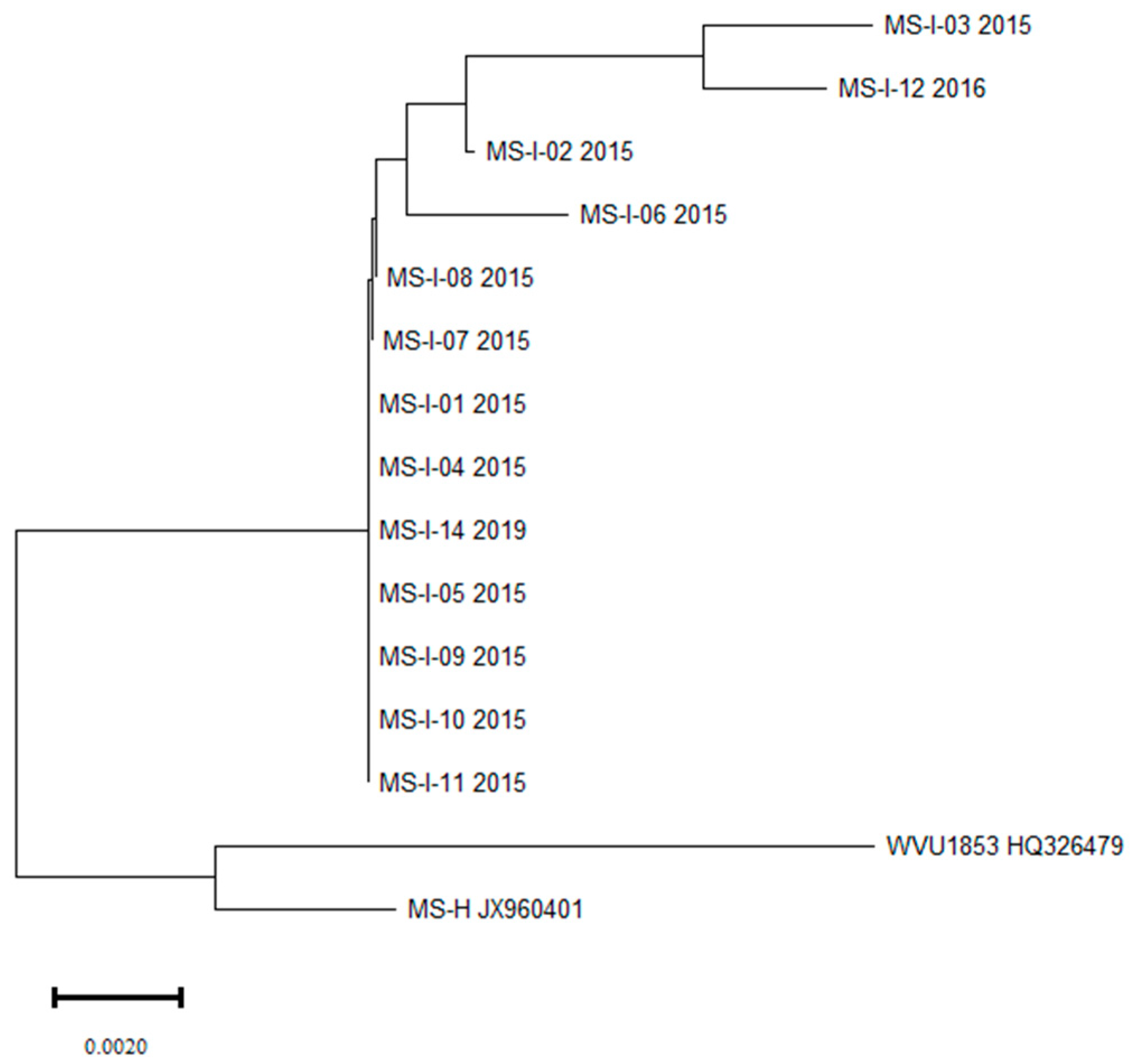
3.2. Molecular Characterisation and Phylogenetic Relationship
3.3. Multi-Infection with Bacterial Pathogens
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Levisohn, S.; Kleven, S.H. Avian Mycoplasmosis (Mycoplasma gallisepticum). Rev. Sci. Tech. 2000, 19, 425–442. [Google Scholar] [CrossRef] [PubMed]
- Feberwee, A.; de Wit, S.; Dijkman, R. Clinical Expression, Epidemiology, and Monitoring of Mycoplasma gallisepticum and Mycoplasma synoviae: An Update. Avian Pathol. 2022, 51, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, H.; Carpenter, A.T.E.; Yamamoto, R. Economic Impact of Mycoplasma gallisepticum and M. synoviae in Commercial Layer Flocks. Avian Dis. 1987, 31, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ishfaq, M.; Fan, Q.; Chen, C.; Li, J. A Respiratory Commensal Bacterium Acts as a Risk Factor for Mycoplasma gallisepticum Infection in Chickens. Vet. Immunol. Immunopathol. 2020, 230, 110127. [Google Scholar] [CrossRef] [PubMed]
- Landman, W.J.M.; Corbanie, E.A.; Feberwee, A.; van Eck, J.H.H. Aerosolization of Mycoplasma synoviae Compared with Mycoplasma gallisepticum and Enterococcus faecalis. Avian Pathol. 2004, 33, 210–215. [Google Scholar] [CrossRef]
- Sivaseelan, S.; Balachandran, P.; Balasubramaniam, G.A.; Madheswaran, R. Synergistic Pathological Effect of Mycoplasma gallisepticum with Ornithobacterium rhinotracheale Infection in Layer Chicken. Indian J. Anim. Sci. 2015, 85, 32–36. [Google Scholar] [CrossRef]
- Sid, H.; Hartmann, S.; Petersen, H.; Ryll, M.; Rautenschlein, S. Mycoplasma gallisepticum Modifies the Pathogenesis of Influenza A Virus in the Avian Tracheal Epithelium. Int. J. Med. Microbiol. 2016, 306, 174–186. [Google Scholar] [CrossRef]
- Walker, R.T. Bacterial Phylogeny with Particular Reference to Mycoplasma. J. Biol. Educ. 1990, 24, 77–80. [Google Scholar] [CrossRef]
- Kleven, S.H. Mycoplasmas in the Etiology of Multifactorial Respiratory Disease. Poult. Sci. 1998, 77, 1146–1149. [Google Scholar] [CrossRef]
- Bradbury, J.M. Avian Mycoplasma Infections: Prototype of Mixed Infections with Mycoplasmas, Bacteria and Viruses. Ann. Inst. Pasteur Microbiol. 1984, 135, 83–89. [Google Scholar] [CrossRef]
- Morrow, C.J.; Bradbury, J.M.; Gentle, M.J.; Thorp, B.H. The Development of Lameness and Bone Deformity in the Broiler Following Experimental Infection with Mycoplasma gallisepticum or Mycoplasma synoviae. Avian Pathol. 1997, 26, 169–187. [Google Scholar] [CrossRef] [PubMed]
- Wyrzykowski, B.; Albaric, O.; Moreau, S.; Nguyen, F.; Fleurance, R.; Belluco, S.; Wyers, M.; Colle, M.A. Retrospective Study of Mycoplasma gallisepticum Meningoencephalitis in Six Turkey Flocks in Western France. J. Comp. Pathol. 2013, 148, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Feberwee, A.; de Wit, J.J.; Landman, W.J.M. Induction of Eggshell Apex Abnormalities by Mycoplasma synoviae: Field and Experimental Studies. Avian Pathol. 2009, 38, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Ferguson-Noel, N.; Noormohammadi, A.H. Mycoplasma synoviae Infection. In Diseases of Poultry; Swayne, D.E., Glisson, J.R., McDougald, L.R., Nolan, L.K., Suarez, D.L., Nair, V.L., Eds.; Wiley: Ames, IO, USA, 2013; pp. 900–906. [Google Scholar]
- Lockaby, S.B.; Hoerr, F.J. Virulence of Mycoplasma synoviae in Poultry: A Review. Worlds Poult. Sci. J. 1999, 55, 175–185. [Google Scholar] [CrossRef]
- Sawicka, A.; Durkalec, M.; Tomczyk, G.; Kursa, O. Occurrence of Mycoplasma gallisepticum in Wild Birds: A Systematic Review and Meta-analysis. PLoS ONE 2020, 15, e0231545. [Google Scholar] [CrossRef] [PubMed]
- Michiels, T.; Welby, S.; Vanrobaeys, M.; Quinet, C.; Rouffaer, L.; Lens, L.; Martel, A.; Butaye, P. Prevalence of Mycoplasma gallisepticum and Mycoplasma synoviae in Commercial Poultry, Racing Pigeons and Wild Birds in Belgium. Avian Pathol. 2016, 45, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Lierz, M.; Hagen, N.; Lueschow, D.; Hafez, H.M. Use of Polymerase Chain Reactions to Detect Mycoplasma gallisepticum, Mycoplasma imitans, Mycoplasma iowae, Mycoplasma meleagridis and Mycoplasma synoviae in Birds of Prey. Avian Pathol. 2008, 37, 471–476. [Google Scholar] [CrossRef]
- Citti, C.; Blanchard, A. Mycoplasmas and Their Host: Emerging and Re-Emerging Minimal Pathogens. Trends Microbiol. 2013, 21, 196–203. [Google Scholar] [CrossRef]
- Pitcher, D.G.; Nicholas, R.A.J. Mycoplasma Host Specificity: Fact or Fiction? Vet. J. 2005, 170, 300–306. [Google Scholar] [CrossRef]
- Dawood, A.; Algharib, S.A.; Zhao, G.; Zhu, T.; Qi, M.; Delai, K.; Hao, Z.; Marawan, M.A.; Shirani, I.; Guo, A. Mycoplasmas as Host Pantropic and Specific Pathogens: Clinical Implications, Gene Transfer, Virulence Factors, and Future Perspectives. Front. Cell Infect. Microbiol. 2022, 12, 855731. [Google Scholar]
- Cizelj, I.; Dušanić, D.; Benčina, D.; Narat, M. Mycoplasma and Host Interaction: In Vitro Gene Expression Modulation in Mycoplasma synoviae and Infected Chicken Chondrocytes. Acta Vet. Hung. 2016, 64, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Kreizinger, Z.; Sulyok, K.M.; Grózner, D.; Beko, K.; Dán, Á.; Szabó, Z.; Gyuranecz, M. Development of Mismatch Amplification Mutation Assays for the Differentiation of MS1 Vaccine Strain from Wild-Type Mycoplasma synoviae and MS-H Vaccine Strains. PLoS ONE 2017, 12, e0175969. [Google Scholar] [CrossRef] [PubMed]
- Kreizinger, Z.; Sulyok, K.M.; Pásztor, A.; Erdélyi, K.; Felde, O.; Povazsán, J.; Korösi, L.; Gyuranecz, M. Rapid, Simple and Cost-Effective Molecular Method to Differentiate the Temperature Sensitive (Ts+) MS-H Vaccine Strain and Wild-Type Mycoplasma synoviae Isolates. PLoS ONE 2015, 10, e0133554. [Google Scholar] [CrossRef] [PubMed]
- Bekő, K.; Kreizinger, Z.; Sulyok, K.M.; Kovács, Á.B.; Grózner, D.; Catania, S.; Bradbury, J.; Lysnyansky, I.; Olaogun, O.M.; Czanik, B.; et al. Genotyping Mycoplasma gallisepticum by Multilocus Sequence Typing. Vet. Microbiol. 2019, 231, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.A.; Marenda, M.S.; Markham, P.F.; Noormohammadi, A.H. Complementation of the Mycoplasma synoviae MS-H Vaccine Strain with Wild-Type Obg Influencing Its Growth Characteristics. PLoS ONE 2018, 13, e0194528. [Google Scholar] [CrossRef] [PubMed]
- Ghorashi, S.A.; Bradbury, J.M.; Ferguson-Noel, N.M.; Noormohammadi, A.H. Comparison of Multiple Genes and 16S-23S RRNA Intergenic Space Region for Their Capacity in High Resolution Melt Curve Analysis to Differentiate Mycoplasma gallisepticum Vaccine Strain Ts-11 from Field Strains. Vet. Microbiol. 2013, 167, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Raviv, Z.; Kleven, S.H. The Development of Diagnostic Real-Time TaqMan PCRs for the Four Pathogenic Avian Mycoplasmas. Avian Dis. 2009, 53, 103–107. [Google Scholar] [CrossRef]
- Ferguson, N.M.; Hepp, D.; Sun, S.; Ikuta, N.; Levisohn, S.; Kleven, S.H.; García, M. Use of Molecular Diversity of Mycoplasma gallisepticum by Gene-Targeted Sequencing (GTS) and Random Amplified Polymorphic DNA (RAPD) Analysis for Epidemiological Studies. Microbiology 2005, 151, 1883–1893. [Google Scholar] [CrossRef]
- WOAH. OIE Avian Mycoplasmosis (Mycoplasma gallisepticum, M. synoviae). In OIE Terrestial Manual; WOAH: Paris, France, 2018; Volume 3, pp. 68–70. [Google Scholar]
- de Jong, A.; Youala, M.; Klein, U.; El Garch, F.; Simjee, S.; Moyaert, H.; Rose, M.; Gautier-Bouchardon, A.V.; Catania, S.; Ganapathy, K.; et al. Minimal Inhibitory Concentration of Seven Antimicrobials to Mycoplasma gallisepticum and Mycoplasma synoviae Isolates from Six European Countries. Avian Pathol. 2021, 50, 161–173. [Google Scholar] [CrossRef]
- Feberwee, A.; de Vries, T.S.; Landman, W.J.M. Seroprevalence of Mycoplasma synoviae in Dutch Commercial Poultry Farms. Avian Pathol. 2008, 37, 629–633. [Google Scholar] [CrossRef]
- Evans, R.D.; Hafez, Y.S. Evaluation of a Mycoplasma gallisepticum Strain Exhibiting Reduced Virulence for Prevention and Control of Poultry Mycoplasmosis. Avian Dis. 1992, 36, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Whithear, K.G.; Soeripto; Harringan, K.E.; Ghiocas, E. Safety of Temperature Sensitive Mutant Mycoplasma gallisepticum Vaccine. Aust. Vet. J. 1990, 67, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Morrow, C.J.; Bell, I.G.; Walker, S.B.; Markham, P.F.; Thorp, B.H.; Whithear, K.G. Isolation of Mycoplasma synoviae from Infectious Synovitis of Chickens. Aust. Vet. J. 1990, 67, 121–124. [Google Scholar] [CrossRef] [PubMed]
- El Gazzar, M.; Laibinis, V.A.; Ferguson-Noel, N. Characterization of a Ts-11–like Mycoplasma gallisepticum Isolate from Commercial Broiler Chickens. Avian Dis. Dig. 2011, 6, e41–e42. [Google Scholar] [CrossRef]
- Brandao, M.D.M.; Santos, F.F.; Machado, L.S.; Verinaud, M.S.; Oliveira, J.M.; Soares, N.M.; Nascimento, E.R.; Pereira, V.L.A. The Effect of Eggshell Apex Abnormalities on Table Egg Quality during Storage in 2 Seasons of the Year. Poult. Sci. 2014, 93, 2657–2662. [Google Scholar] [CrossRef]
- Lara, L.J.; Rostagno, M.H. Impact of Heat Stress on Poultry Production. Animals 2013, 3, 356–369. [Google Scholar] [CrossRef]
- Kers, J.G.; Velkers, F.C.; Fischer, E.A.J.; Hermes, G.D.A.; Stegeman, J.A.; Smidt, H. Host and Environmental Factors Affecting the Intestinal Microbiota in Chickens. Front. Microbiol. 2018, 9, 235. [Google Scholar] [CrossRef]
- Landman, W.J.M. Is Mycoplasma synoviae outrunning Mycoplasma gallisepticum? A viewpoint from the Netherlands. Avian Pathol. 2014, 43, 2–8. [Google Scholar] [CrossRef]
- Landman, W.J.M.; Feberwee, A. Field Studies on the Association between Amyloid Arthropathy and Mycoplasma synoviae Infection, and Experimental Reproduction of the Condition in Brown Layers. Avian Pathol. 2001, 30, 629–639. [Google Scholar] [CrossRef]
- Kursa, O.; Paku, A.; Tomczyk, G.; Sawicka, A. Eggshell Apex Abnormalities Caused by Two Different Mycoplasma synoviae Genotypes and Evaluation of Eggshell Anomalies by Full-Field Optical Coherence Tomography. BMC Vet. Res. 2019, 15, 1. [Google Scholar] [CrossRef]
- Landman, W.J.; Feberwee, A. Aerosol-Induced Mycoplasma synoviae Arthritis: The Synergistic Effect of Infectious Bronchitis Virus Infection. Avian Pathol. 2004, 33, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Landman, W.J.M.; Feberwee, A.; Veldman, K.T.; Mevius, D.J. Epidemiology: Study on the Vertical Transmission of Arthropathic and Amyloidogenic Enterococcus faecalis in a Flock of Brown Layer Chickens. Vet. Q. 2001, 23, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Śmietanka, K.; Minta, Z.; Świętoń, E.; Olszewska, M.; Jóźwiak, M.; Domańska-Blicharz, K.; Wyrostek, K.; Tomczyk, G.; Pikuła, A. Avian Influenza H9N2 Subtype in Poland—Characterization of the Isolates and Evidence of Concomitant Infections. Avian Pathol. 2014, 43, 427–436. [Google Scholar] [CrossRef]
- Marouf, S.; Khalf, M.A.; Alorabi, M.; El-Shehawi, A.M.; El-Tahan, A.M.; El-Hack, M.E.A.; El-Saadony, M.T.; Salem, H.M. Mycoplasma gallisepticum: A Devastating Organism for the Poultry Industry in Egypt. Poult. Sci. 2022, 101, 101658. [Google Scholar] [CrossRef]
- Wei, X.; Zhong, Q.; Wang, D.; Yan, Z.; Liang, H.; Zhou, Q.; Chen, F. Epidemiological Investigations and Multilocus Sequence Typing of Mycoplasma gallisepticum Collected in China. Poult. Sci. 2023, 102, 102930. [Google Scholar] [CrossRef] [PubMed]
- Galluzzo, P.; Migliore, S.; Galuppo, L.; Condorelli, L.; Hussein, H.A.; Licitra, F.; Coltraro, M.; Sallemi, S.; Antoci, F.; Cascone, G.; et al. First Molecular Survey to Detect Mycoplasma gallisepticum and Mycoplasma synoviae in Poultry Farms in a Strategic Production District of Sicily (South-Italy). Animals 2022, 12, 962. [Google Scholar] [CrossRef]
- Al-dabbagh, S.Y.A.; Rasheed, B.Y.; Al-Jumaa, Z.M. Molecular Diagnosis of Mycoplasma gallisepticum in Turkey in Mosul City. Vet. Pract. 2021, 22. [Google Scholar]
- Abbas, N.; Suleman, M.; Khan, N.A.; Ali, I.; Rauf, M.; Ur Rahman, S. Prevalence of Mycoplasma gallisepticum in Poultry and Wild Life Birds Suspected of Chronic Respiratory Disease in Northern Pakistan. Pak. J. Zool. 2018, 50, 1071–1077. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Year | Number of Samples from Broilers | MG-Positive Samples | MS-Positive Samples | ||
|---|---|---|---|---|---|
| % | n | % | n | ||
| 2015 | 61 | 1.64% | 1 | 19.67% | 12 |
| 2016 | 15 | 6.67% | 1 | 6.67% | 1 |
| 2017 | 10 | - | - | - | - |
| 2018 | 6 | - | - | - | - |
| 2019 | 16 | 50% | 8 | 6.25% | 1 |
| 2020 | 5 | - | - | - | |
| 2021 | 11 | - | - | - | |
| 2022 | 0 | - | - | - | |
| 2023 | 20 | 10% | 2 | - | - |
| Total | 144 | 8.33% | 12 | 9.72% | 14 |
| Year | MS | MG | ORT | GA | E. faecalis | E. faecium | S. aureus | R. anatipestifer |
|---|---|---|---|---|---|---|---|---|
| 2015 | 12 | 1 | 10 | 2 | 2 | 0 | 0 | 0 |
| 2016 | 1 | 1 | 2 | 1 | 2 | 0 | 0 | 0 |
| 2019 | 1 | 8 | 9 | 4 | 0 | 0 | 0 | 0 |
| 2023 | 0 | 2 | 2 | 1 | 2 | 2 | 1 | 1 |
| Total | 14 | 12 | 23 | 8 | 6 | 2 | 1 | 1 |
| Total% | 9.72 | 8.33 | 88.46 | 30.77 | 23.08 | 7.69 | 3.85 | 3.85 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kursa, O.; Tomczyk, G.; Sieczkowska, A.; Kostka, S.; Sawicka-Durkalec, A. Mycoplasma gallisepticum and Mycoplasma synoviae in Turkeys in Poland. Pathogens 2024, 13, 78. https://doi.org/10.3390/pathogens13010078
Kursa O, Tomczyk G, Sieczkowska A, Kostka S, Sawicka-Durkalec A. Mycoplasma gallisepticum and Mycoplasma synoviae in Turkeys in Poland. Pathogens. 2024; 13(1):78. https://doi.org/10.3390/pathogens13010078
Chicago/Turabian StyleKursa, Olimpia, Grzegorz Tomczyk, Agata Sieczkowska, Sylwia Kostka, and Anna Sawicka-Durkalec. 2024. "Mycoplasma gallisepticum and Mycoplasma synoviae in Turkeys in Poland" Pathogens 13, no. 1: 78. https://doi.org/10.3390/pathogens13010078
APA StyleKursa, O., Tomczyk, G., Sieczkowska, A., Kostka, S., & Sawicka-Durkalec, A. (2024). Mycoplasma gallisepticum and Mycoplasma synoviae in Turkeys in Poland. Pathogens, 13(1), 78. https://doi.org/10.3390/pathogens13010078