
Evaluation of Antibacterial and Antibiofilm Activity of Rice Husk Extract against Staphylococcus aureus


 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Extract Sample Preparation
2.2. Bacterial Strains
2.3. Antimicrobial Testing
2.4. Growth Curve Kinetics
2.5. Inhibition of Biofilm Formation
2.6. Biofilm Disaggregation Assay
2.7. Confocal Laser Scanning Microscopy (CLSM)
2.8. Toxicity in Galleria mellonella Larvae
2.9. Checkerboard Test
2.10. Statistical Analysis
3. Results
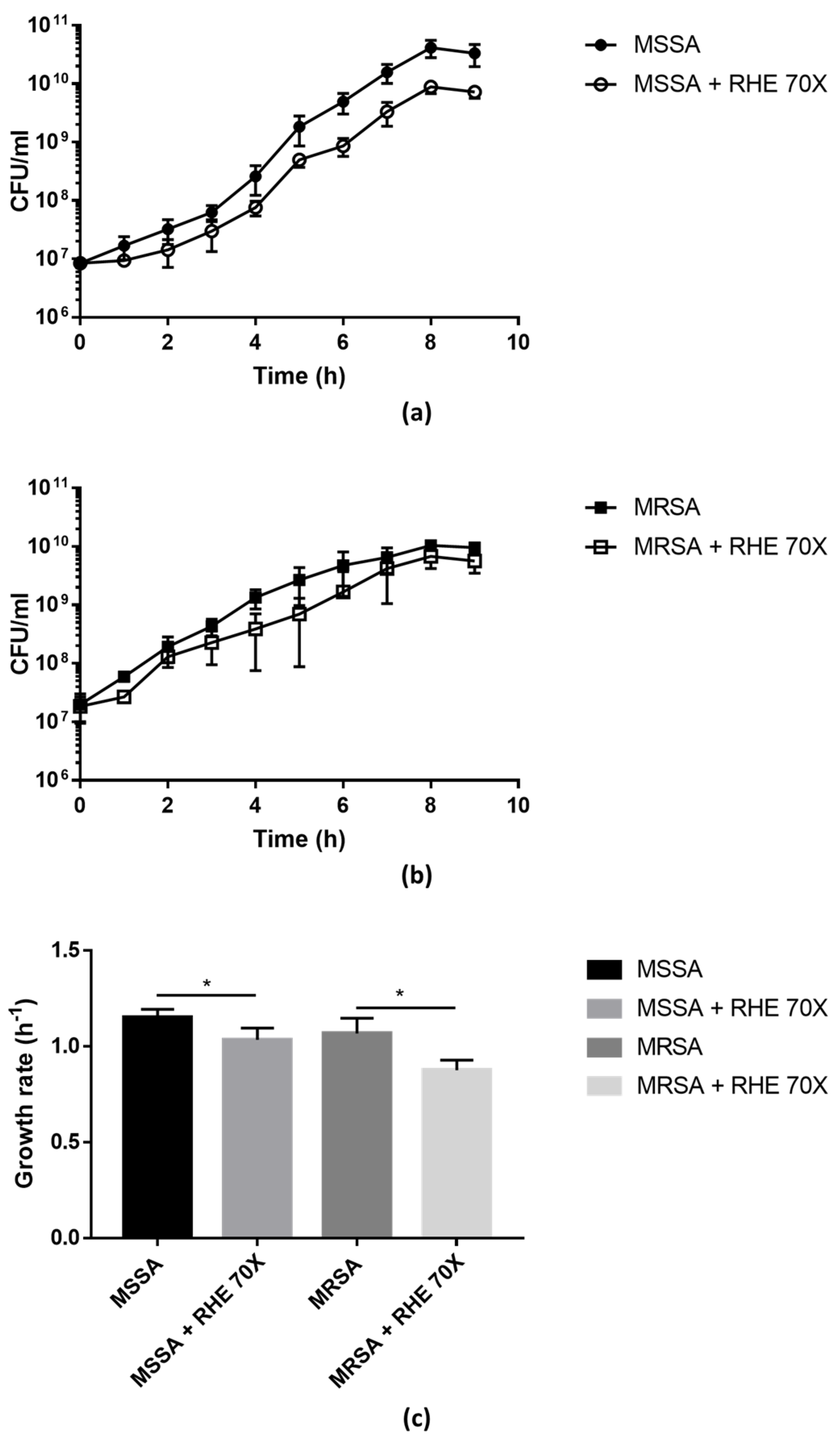
3.1. Antimicrobial Activity
3.2. Antibiofilm Activity
3.3. Toxicity
3.4. Interaction with Antibiotics
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheung, G.Y.C.; Bae, J.S.; Otto, M. Pathogenicity and virulence of Staphylococcus aureus. Virulence 2021, 12, 547–569. [Google Scholar] [CrossRef]
- Gould, I.M.; David, M.Z.; Esposito, S.; Garaud, J.; Lina, G.; Mazzei, T.; Peters, G. New insights into meticillin-resistant Staphylococcus aureus (MRSA) pathogenesis, treatment and resistance. Int. J. Antimicrob. Agents 2012, 39, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G.J. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed]
- Van Hal, S.J.; Jensen, S.O.; Vaska, V.L.; Espedido, B.A.; Paterson, D.L.; Gosbell, I.B. Predictors of mortality in Staphylococcus aureus Bacteremia. Clin. Microbiol. Rev. 2012, 25, 362–386. [Google Scholar] [CrossRef]
- Abraham, L.; Bamberger, D.A. Staphylococcus aureus Bacteremia: Contemporary Management. Mo Med. 2020, 117, 341–345. [Google Scholar]
- O’Toole, G.A. Microtiter Dish Biofilm Formation Assay. J. Vis. Exp. 2011, 47, e2437. [Google Scholar] [CrossRef]
- Howden, B.P.; Giulieri, S.G.; Lung, T.W.F.; Baines, S.L.; Sharkey, L.K.; Lee, J.Y.H.; Hachani, A.; Monk, I.R.; Stinear, T.P. Staphylococcus aureus host interactions and adaptation. Nat. Rev. Microbiol. 2023, 21, 380–395. [Google Scholar] [CrossRef]
- Archer, N.K.; Mazaitis, M.J.; Costerton, J.W.; Leid, J.G.; Powers, M.E.; Shirtliff, M.E. Staphylococcus aureus biofilms Properties, regulation and roles in human disease. Virulence 2011, 2, 445–459. [Google Scholar] [CrossRef]
- Cordeiro, L.; Figueiredo, P.; Souza, H.; Sousa, A.; Andrade-Júnior, F.; Barbosa-Filho, J.; Lima, E. Antibacterial and Antibiofilm Activity of Myrtenol against Staphylococcus aureus. Pharmaceuticals 2020, 13, 133. [Google Scholar] [CrossRef]
- Gao, Y.; Guo, X.; Liu, Y.; Fang, Z.; Zhang, M.; Zhang, R.; You, L.; Li, T.; Liu, R.H. A full utilization of rice husk to evaluate phytochemical bioactivities and prepare cellulose nanocrystals. Sci. Rep. 2018, 8, 10482. [Google Scholar] [CrossRef]
- Manso, T.; Lores, M.; de Miguel, T. Antimicrobial Activity of Polyphenols and Natural Polyphenolic Extracts on Clinical Isolates. Antibiotics 2022, 11, 46. [Google Scholar] [CrossRef]
- Frosi, I.; Vallelonga, D.; Colombo, R.; Milanese, C.; Papetti, A. Valorization of Rice Hushk (Oryza Sativa L) as a source of in Vitro antiglycative and antioxidant agents. Foods 2023, 12, 529. [Google Scholar] [CrossRef] [PubMed]
- Minh, T.N.; Xuan, T.D.; Ahmad, A.; Elzaawely, A.A.; Teschke, R.; Van, T.M. Efficacy from Different Extractions for Chemical Profile and Biological Activities of Rice Husk. Sustainability 2018, 10, 1356. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Nguyen, M.T.; Nguyen, H.D.; Pham, P.D.; Thach, U.D.; Trinh, B.T.D.; Nguyen, L.T.T.; Dang, S.V.; Do, A.T.; Do, B.H. Antioxidant and Antimicrobial Activities of the Extracts from Different Garcinia Species. Evid-Based Complement. Altern. Med. 2021, 2021, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, T.; Chatterjee, T.K.; Majumdar, D.; Chakrabarti, P. Antibacterial effect of silver nanoparticles and the modeling of bacterial growth kinetics using a modified Gompertz model. Biochim. Biophys. Acta (BBA)—Gen. Subj. 2015, 1850, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Papetti, A.; Signoretto, C.; Spratt, D.A.; Pratten, J.; Lingstrom, P.; Zaura, E.; Ofek, I.; Wilson, M.; Pruzzo, C.; Gazzani, G. Components in Lentinus edodes mushroom with anti-biofilm activity directed against bacteria involved in caries and gingivitis. Food Funct. 2018, 9, 3489–3499. [Google Scholar] [CrossRef]
- Stepanović, S.; Vuković, D.; Hola, V.; Di Bonaventura, G.; Djukić, S.; Cirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef]
- Neu, T.R.; Lawrence, J.R. Development and structure of microbial biofilms in river water studied by confocal laser scanning microscopy. FEMS Microbiol. Ecol. 1997, 24, 11–25. [Google Scholar] [CrossRef]
- Jia-Yun Tsai, C.; San Loh, J.M.; Proft, P. Galleria mellonella infection models for the study of bacterial diseases and for antimicrobial drug testing. Virulence 2016, 7, 214–229. [Google Scholar] [CrossRef]
- Sopirala, M.M.; Mangino, J.E.; Gebreyes, W.A.; Biller, B.; Bannerman, T.; Balada-Llasat, J.M.; Pancholi, P. Synergy testing by Etest, microdilution checkerboard, and time-kill methods for pan-drug-resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 2010, 54, 4678–4683. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. 2017. Available online: http://www.who.int/medicines/publications/global-priority-list-antibiotic-resistant-bacteria/en/ (accessed on 27 February 2017).
- Cascioferro, S.; Carbone, D.; Parrino, B.; Pecoraro, C.; Giovannetti, E.; Cirrincione, G.; Diana, P. Therapeutic Strategies to Counteract Antibiotic Resistance in MRSA Biofilm-Associated Infections. ChemMedChem 2021, 16, 65–80. [Google Scholar] [CrossRef]
- Altemini, A.; Lakhssassi, N.; Baharlouei, A.; Watson, D.; Lightfoot, D. Phytochemicals: Extraction, isolation, and identification of bioactive compounds from plant extracts. Plants 2017, 6, 42. [Google Scholar] [CrossRef]
- Agourram, A.; Ghirardello, D.; Rantsiou, K.; Zeppa, G.; Belviso, S.; Romane, A.; Oufdou, K.; Giordano, M. Phenolic content, antioxidant potential, and antimicrobial activities of fruit and vegetable by product extracts. Int. J. Food Propert 2013, 16, 1092–1104. [Google Scholar] [CrossRef]
- Gil-Chávez, J.G.; Villa, J.A.; Ayala-Zavala, F.J.; Heredia, B.J.; Sepulveda, D.; Yahia, E.M.; González-Aguilar, G.A. Technologies for extraction and production of bioactive compounds to be used as nutraceuticals and food ingredients: An overview. Compr. Rev. Food Sci. Food Saf. 2013, 12, 5–23. [Google Scholar] [CrossRef]
- Baiano, A. Recovery of biomolecules from food wastes—A review. Molecules 2014, 19, 14821–14842. [Google Scholar] [CrossRef] [PubMed]
- Lakshmana Prabu, S.; Umamaheswari, A.; Grace Felciya, S.J. Investigation on the biofilm eradication potential of selected medicinal plants against methicillin-resistant Staphylococcus aureus. Biotechnol. Rep. 2020, 28, e00523. [Google Scholar] [CrossRef]
- Akinduti, P.A.; Emoh-Robinson, V.; Obamoh-Triumphant, H.F.; Obafemi, Y.D.; Banjo, T.T. Antibacterial activities of plant leaf extracts against multi-antibiotic resistant Staphylococcus aureus associated with skin and soft tissue infections. BMC Complement. Med. Ther. 2022, 22, 47. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos Ramos, F.; Midon Martins, S.; Sagini, N.; Brandão Quines, C.; de Oliveira Pereira, F.S.; de Ávila, A.S.; Zanzarind, D.; Pilau, E.D.; de Lima Postiga, I.A.; Tostes, J.; et al. Soybeans agroindustrial residues as Stahylococcus epidermidis and S. aureus biofilm inhibitors. Ind. Crops Prod. 2021, 170, 113713. [Google Scholar] [CrossRef]
- Meilina, H.; Faisal, M. Antibacterial activity of liquid smoke powder from rice husk. Int. J. GEOMATE 2022, 23, 89–96. [Google Scholar] [CrossRef]
- Butstat, S.; Siriamornpun, S. Phenolic acids and antioxidant activities in husk of different Thai rice varieties. Food Sci. Technol. Int. 2010, 16, 329–335. [Google Scholar] [CrossRef]
- Irakli, M.; Kleisiaris, F.; Kadoglidou, K.; Katsantionis, D. Optimizing extraction conditions of free and bound phenolic compounds from rice by-products and their antioxidant effects. Foods 2018, 7, 93. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, S.A.Z.; Irfan, A.; Zaheer, S.; Sultan, A.; Shajahan, S.; Rubab, S.L.; Ain, Q.; Acevedo, R. Proximate composition of orange peel, pea peel and rice husk wastes and their potential use as antimicrobial agents and antioxidants. Vegetos 2021, 34, 470–476. [Google Scholar] [CrossRef]
- Fu, J.; Zhang, Y.; Lin, S.; Zhang, W.; Shu, G.; Lin, J.; Li, H.; Xu, F.; Tang, H.; Peng, G.; et al. Strategies for Interfering with Bacterial Early Stage Biofilms. Front. Microbiol. 2021, 12, 675843. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Hu, W.; Tian, Z.; Yuan, D.; Yi, G.; Zhou, Y.; Cheng, Q.; Zhu, J.; Li, M. Developing natural products as potential anti-biofilm agents. Chin. Med. 2019, 14, 11. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| RHE Concentration | MSSA | MRSA |
|---|---|---|
| 20× | 0 | 0 |
| 40× | 4 | 3 |
| 60× | 10 | 9 |
| 70× | 13 | 12 |
| 80× | 15 | 14 |
| RHE Concentration | MSSA (%) | MRSA (%) |
|---|---|---|
| 20× | 0 ± 0.2 | 0 ± 0.5 |
| 40× | 5 ± 1.2 | 8 ± 1.0 |
| 60× | 25 ± 4.2 | 30 ± 2.2 |
| 70× | 45 ± 3.7 | 50 ± 3.2 |
| 80× | 70 ± 5.9 | 75 ± 3.8 |
| RHE Concentration | MSSA (%) | MRSA (%) |
|---|---|---|
| 20× | 0 ± 0.3 | 0 ± 0.4 |
| 40× | 3 ± 0.7 | 3 ± 0.9 |
| 60× | 9 ± 1,0 | 8 ± 1.1 |
| 70× | 15 ± 1.7 | 12 ± 2.0 |
| 80× | 20 ± 1.8 | 20 ± 1.5 |
| Treatment | 24 h | 48 h | 72 h |
|---|---|---|---|
| Sterile saline solution | 0/10 | 0/10 | 0/10 |
| Ethanol | 0/10 | 0/10 | 1/10 |
| RHE 60× | 0/10 | 0/10 | 0/10 |
| RHE 70× | 0/10 | 0/10 | 0/10 |
| RHE 80× | 0/10 | 0/10 | 1/10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burlacchini, G.; Sandri, A.; Papetti, A.; Frosi, I.; Boschi, F.; Lleo, M.M.; Signoretto, C. Evaluation of Antibacterial and Antibiofilm Activity of Rice Husk Extract against Staphylococcus aureus. Pathogens 2024, 13, 80. https://doi.org/10.3390/pathogens13010080
Burlacchini G, Sandri A, Papetti A, Frosi I, Boschi F, Lleo MM, Signoretto C. Evaluation of Antibacterial and Antibiofilm Activity of Rice Husk Extract against Staphylococcus aureus. Pathogens. 2024; 13(1):80. https://doi.org/10.3390/pathogens13010080
Chicago/Turabian StyleBurlacchini, Gloria, Angela Sandri, Adele Papetti, Ilaria Frosi, Federico Boschi, Maria M. Lleo, and Caterina Signoretto. 2024. "Evaluation of Antibacterial and Antibiofilm Activity of Rice Husk Extract against Staphylococcus aureus" Pathogens 13, no. 1: 80. https://doi.org/10.3390/pathogens13010080
APA StyleBurlacchini, G., Sandri, A., Papetti, A., Frosi, I., Boschi, F., Lleo, M. M., & Signoretto, C. (2024). Evaluation of Antibacterial and Antibiofilm Activity of Rice Husk Extract against Staphylococcus aureus. Pathogens, 13(1), 80. https://doi.org/10.3390/pathogens13010080