The Pathogenesis of Staphylococcus aureus Eye Infections


Abstract
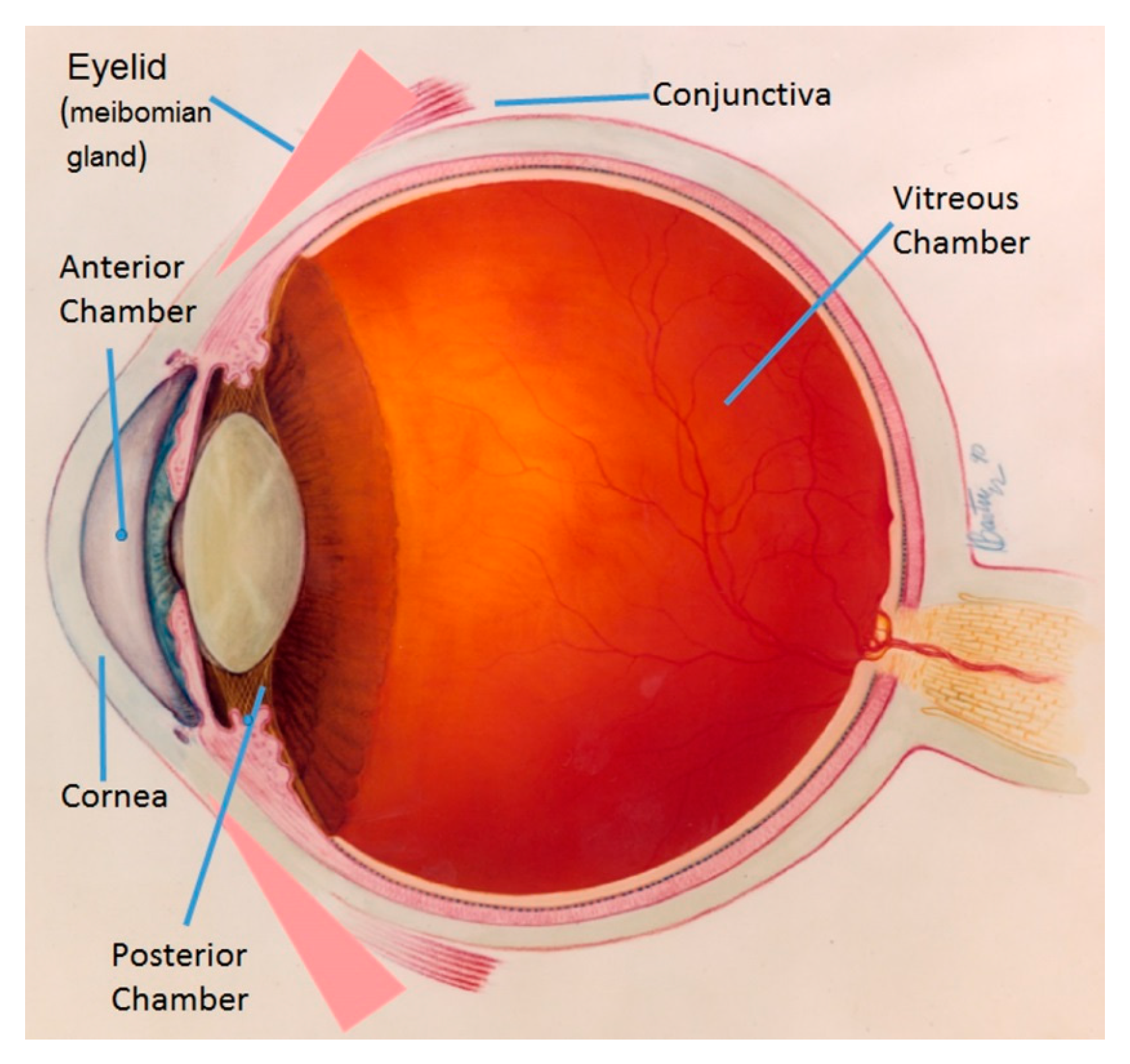
:1. Introduction
2. Meibomian Gland and Blepharitis
3. Dacryocystitis
4. Conjunctivitis
5. Keratitis
5.1. Introduction
5.2. Bacterial Virulence
5.3. Host Response
5.4. Conclusions
6. Endophthalmitis
6.1. Introduction
6.2. Bacterial Virulence
6.3. Host Response to Infection
6.4. Conclusions
7. Final Comment
Acknowledgment
Conflicts of Interest
References
- Shine, W.E.; Silvany, R.; McCulley, J.P. Relation of cholesterol-stimulated Staphylococcus aureus growth to chronic blepharitis. Investig. Ophthalmol. Vis. Sci. 1993, 34, 2291–2296. [Google Scholar]
- Azari, A.A.; Barney, N.P. Conjunctivitis: A systematic review of diagnosis and treatment. JAMA 2013, 310, 1721–1730. [Google Scholar] [CrossRef] [PubMed]
- Liesegang, T.J. Bacterial Keratitis. In The Cornea; Kaufman, H.E., Barron, B.A., McDonald, M.B., Eds.; Butterworth-Heinemann: Boston, MA, USA, 1999; ISBN 978-0-7506-7132-3. [Google Scholar]
- Wong, V.W.Y.; Lai, T.Y.Y.; Chi, S.C.C.; Lam, D.S.C. Pediatric Ocular surface infections: A 5-year review of demographics, clinical features, risk factors, microbiological results, and treatment. Cornea 2011, 30, 995–1002. [Google Scholar] [CrossRef] [PubMed]
- Kerr, N.; Stern, G.A. Bacterial keratitis associated with vernal keratoconjunctivitis. Cornea 1992, 11, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.L.; Hyndiuk, R.A. Contact lens-related infectious keratitis. Int. Ophthalmol. Clin. 1993, 33, 23–49. [Google Scholar] [CrossRef] [PubMed]
- Tacconelli, E.; Johnson, A.P. National guidelines for decolonization of methicillin-resistant Staphylococcus aureus carriers: The implications of recent experience in the Netherlands. J. Antimicrob. Chemother. 2011, 66, 2195–2198. [Google Scholar] [CrossRef] [PubMed]
- Durand, M.L. Endophthalmitis. Clin. Microbiol. Infect. 2013, 19, 227–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kluytmans, J.; van Belkum, A.; Verbrugh, H. Nasal carriage of Staphylococcus aureus: Epidemiology, underlying mechanisms, and associated risks. Clin. Microbiol. Rev. 1997, 10, 505–520. [Google Scholar] [PubMed]
- Rashid, Z.; Farzana, K.; Sattar, A.; Murtaza, G. Prevalence of nasal Staphylococcus aureus and methicillin-resistant Staphylococcus aureus in hospital personnel and associated risk factors. Acta Pol. Pharm. 2012, 69, 985–991. [Google Scholar] [PubMed]
- Yan, M.; Pamp, S.J.; Fukuyama, J.; Hwang, P.H.; Cho, D.-Y.; Holmes, S.; Relman, D.A. Nasal microenvironments and interspecific interactions influence nasal microbiota complexity and S. aureus carriage. Cell Host Microbe 2013, 14, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.A.; Jackson, C.R.; Fedorka-Cray, P.J.; Barrett, J.B.; Brousse, J.H.; Gustafson, J.; Kucher, M. Carriage of methicillin-resistant staphylococci by healthy companion animals in the US. Lett. Appl. Microbiol. 2014, 59, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Fluit, A.C. Livestock-associated Staphylococcus aureus. Clin. Microbiol. Infect. 2012, 18, 735–744. [Google Scholar] [CrossRef] [PubMed]
- Schaumburg, F.; Mugisha, L.; Kappeller, P.; Fichtel, C.; Köck, R.; Köndgen, S.; Becker, K.; Boesch, C.; Peters, G.; Leendertz, F. Evaluation of non-invasive biological samples to monitor Staphylococcus aureus colonization in great apes and lemurs. PLoS ONE 2013, 8, e78046. [Google Scholar] [CrossRef]
- Larsen, J.; Petersen, A.; Sørum, M.; Stegger, M.; van Alphen, L.; Valentiner-Branth, P.; Knudsen, L.; Larsen, L.; Feingold, B.; Price, L.; et al. Meticillin-resistant Staphylococcus aureus CC398 is an increasing cause of disease in people with no livestock contact in Denmark, 1999 to 2011. Eurosurveillance 2015, 20. [Google Scholar] [CrossRef] [PubMed]
- Köck, R.; Harlizius, J.; Bressan, N.; Laerberg, R.; Wieler, L.H.; Witte, W.; Deurenberg, R.H.; Voss, A.; Becker, K.; Friedrich, A.W. Prevalence and molecular characteristics of methicillin-resistant Staphylococcus aureus (MRSA) among pigs on German farms and import of livestock-related MRSA into hospitals. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 1375. [Google Scholar] [CrossRef] [PubMed]
- Speaker, M.G.; Milch, F.A.; Shah, M.K.; Eisner, W.; Kreiswirth, B.N. Role of external bacterial flora in the pathogenesis of acute postoperative endophthalmitis. Ophthalmology 1991, 98, 639–650. [Google Scholar] [CrossRef]
- McCulley, J.P.; Shine, W.E. Meibomian gland function and the tear lipid layer. Ocul. Surf. 2003, 1, 97–106. [Google Scholar] [CrossRef]
- Aronowicz, J.D.; Shine, W.E.; Oral, D.; Vargas, J.M.; McCulley, J.P. Short term oral minocycline treatment of meibomianitis. Br. J. Ophthalmol. 2006, 90, 856–860. [Google Scholar] [CrossRef] [PubMed]
- Wladis, E.J.; Bradley, E.A.; Bilyk, J.R.; Yen, M.T.; Mawn, L.A. Oral Antibiotics for Meibomian Gland-Related Ocular Surface Disease: A report by the American Academy of Ophthalmology. Ophthalmology 2016, 123, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Qiao, J.; Yan, X. Emerging treatment options for meibomian gland dysfunction. Clin. Ophthalmol. (Auckl. N. Z.) 2013, 7, 1797–1803. [Google Scholar] [CrossRef]
- Huber-Spitzy, V.; Steinkogler, F.J.; Huber, E.; Arocker-Mettinger, E.; Schiffbänker, M. Acquired dacryocystitis: Microbiology and conservative therapy. Acta Ophthalmol. 1992, 70, 745–749. [Google Scholar] [CrossRef]
- Sugita, J.; Yokoi, N.; Fullwood, N.J.; Quantock, A.J.; Takada, Y.; Nakamura, Y.; Kinoshita, S. The detection of bacteria and bacterial biofilms in punctal plug holes. Cornea 2001, 20, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcus epidermidis pathogenesis. Methods Mol. Biol. 2014, 1106, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Kugadas, A.; Christiansen, S.H.; Sankaranarayanan, S.; Surana, N.K.; Gauguet, S.; Kunz, R.; Fichorova, R.; Vorup-Jensen, T.; Gadjeva, M. Impact of microbiota on resistance to ocular Pseudomonas aeruginosa-induced keratitis. PLOS Pathog. 2016, 12, e1005855. [Google Scholar] [CrossRef] [PubMed]
- St. Leger, A.J.; Desai, J.V.; Drummond, R.A.; Kugadas, A.; Almaghrabi, F.; Silver, P.; Raychaudhuri, K.; Gadjeva, M.; Iwakura, Y.; Lionakis, M.S.; et al. An ocular commensal protects against corneal infection by driving an Interleukin-17 response from mucosal γδ T cells. Immunity 2017, 47, 148–158.e5. [Google Scholar] [CrossRef] [PubMed]
- Alfonso, S.A.; Fawley, J.D.; Lu, X.A. Conjunctivitis. Prim. Care Clin. Off. Pract. 2015, 42, 325–345. [Google Scholar] [CrossRef] [PubMed]
- Cronau, H.; Kankanala, R.R.; Mauger, T. Diagnosis and management of red eye in primary care. Am. Fam. Physician 2010, 81, 137–144. [Google Scholar] [PubMed]
- Fujishima, H.; Okada, N.; Dogru, M.; Baba, F.; Tomita, M.; Abe, J.; Matsumoto, K.; Saito, H. The role of Staphylococcal enterotoxin in atopic keratoconjunctivitis and corneal ulceration. Allergy 2012, 67, 799–803. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.-H.; Nam, K.-H.; Kweon, M.-N. Staphylococcus aureus accelerates an experimental allergic conjunctivitis by Toll-like receptor 2-dependent manner. Clin. Immunol. 2009, 131, 170–177. [Google Scholar] [CrossRef] [PubMed]
- McGilligan, V.E.; Gregory-Ksander, M.S.; Li, D.; Moore, J.E.; Hodges, R.R.; Gilmore, M.S.; Moore, T.C.B.; Dartt, D.A. Staphylococcus aureus activates the NLRP3 inflammasome in human and rat conjunctival goblet cells. PLoS ONE 2013, 8, e74010. [Google Scholar] [CrossRef] [PubMed]
- McCormick, C.C.; Caballero, A.R.; Balzli, C.L.; Tang, A.; Weeks, A.; O’Callaghan, R.J. Diverse virulence of Staphylococcus aureus strains for the conjunctiva. Curr. Eye Res. 2011, 36, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Tang, A.; Balzli, C.L.; Caballero, A.R.; McCormick, C.C.; Taylor, S.D.; O’Callaghan, R.J. Staphylococcus aureus infection of the rabbit cornea following topical administration. Curr. Eye Res. 2012, 37, 1075–1083. [Google Scholar] [CrossRef] [PubMed]
- Moreau, J.M.; Girgis, D.O.; Hume, E.B.H.; Dajcs, J.J.; Austin, M.S.; O’Callaghan, R.J. Phospholipase A2 in rabbit tears: A Host Defense against Staphylococcus aureus. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2347–2354. [Google Scholar]
- Davidson, H.J.; Kuonen, V.J. The tear film and ocular mucins. Vet. Ophthalmol. 2004, 7, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.J.; Fleiszig, S.M.J. Why does the healthy cornea resist Pseudomonas aeruginosa infection? Am. J. Ophthalmol. 2013, 155, 961–970.e2. [Google Scholar] [CrossRef] [PubMed]
- Gordon, Y.J.; Huang, L.C.; Romanowski, E.G.; Yates, K.A.; Proske, R.J.; McDermott, A.M. Human cathelicidin (LL-37), a multifunctional peptide, is expressed by ocular surface epithelia and has potent antibacterial and antiviralactivity. Curr. Eye Res. 2005, 30, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Yamada, A.; Gilmore, M.S. Host-pathogen interactions in the cornea. Jpn. J. Ophthalmol. 2010, 54, 191–193. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Epand, R.F.; Mishra, B.; Lushnikova, T.; Thomas, V.C.; Bayles, K.W.; Epand, R.M. Decoding the functional roles of cationic side chains of the major antimicrobial region of human cathelicidin LL-37. Antimicrob. Agents Chemother. 2012, 56, 845–856. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.C.; Jean, D.; Proske, R.J.; Reins, R.Y.; McDermott, A.M. Ocular surface expression and in vitro activity of antimicrobial peptides. Curr. Eye Res. 2007, 32, 595–609. [Google Scholar] [CrossRef] [PubMed]
- Haynes, R.J.; McElveen, J.E.; Dua, H.S.; Tighe, P.J.; Liversidge, J. Expression of human beta-defensins in intraocular tissues. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3026–3031. [Google Scholar]
- Li, Q.; Kumar, A.; Gui, J.-F.; Yu, F.-S.X. Staphylococcus aureus lipoproteins trigger human corneal epithelial innate response through toll-like receptor-2. Microb. Pathog. 2008, 44, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Cole, N.; Hume, E.B.; Khan, S.; Garthwaite, L.; Schubert, T.; Reeve, V.; Willcox, M.D.P. The corneal response to infection with Staphylococcus aureus in the absence of interleukin-4. Immunol. Cell Biol. 2007, 85, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Cole, N.; Hume, E.B.H.; Khan, S.; Garthwaite, L.; Conibear, T.C.R.; Willcox, M.D.P. The role of CXC chemokine receptor 2 in Staphylococcus aureus keratitis. Exp. Eye Res. 2014, 127, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Heimer, S.R.; Yamada, A.; Russell, H.; Gilmore, M. Response of corneal epithelial cells to Staphylococcus aureus. Virulence 2010, 1, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, J.L.; Willcox, M.D.P. Role of lactoferrin in the tear film. Biochimie 2009, 91, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Fluckinger, M.; Haas, H.; Merschak, P.; Glasgow, B.J.; Redl, B. Human tear lipocalin exhibits antimicrobial activity by scavenging microbial siderophores. Antimicrob. Agents Chemother. 2004, 48, 3367–3372. [Google Scholar] [CrossRef] [PubMed]
- Willcox, M.D.; Morris, C.A.; Thakur, A.; Sack, R.A.; Wickson, J.; Boey, W. Complement and complement regulatory proteins in human tears. Investig. Ophthalmol. Vis. Sci. 1997, 38, 1–8. [Google Scholar]
- Qu, X.-D.; Lehrer, R.I. Secretory Phospholipase A2 Is the principal bactericide for Staphylococci and other gram-positive bacteria in human tears. Infect. Immun. 1998, 66, 2791–2797. [Google Scholar] [PubMed]
- Pescosolido, N.; Barbato, A.; Pascarella, A.; Giannotti, R.; Genzano, M.; Nebbioso, M. Role of protease-inhibitors in ocular diseases. Molecules 2014, 19, 20557–20569. [Google Scholar] [CrossRef] [PubMed]
- Bräuer, L.; Kindler, C.; Jäger, K.; Sel, S.; Nölle, B.; Pleyer, U.; Ochs, M.; Paulsen, F.P. Detection of surfactant proteins A and D in human tear fluid and the human lacrimal system. Investig. Ophthalmol. Vis. Sci. 2007, 48, 3945–3953. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Abdel-Razek, O.; Hawgood, S.; Wang, G. Protective role of surfactant protein D in ocular Staphylococcus aureus infection. PLoS ONE 2015, 10, e0138597. [Google Scholar] [CrossRef] [PubMed]
- Jumblatt, M.M.; Imbert, Y.; Young, W.W.; Foulks, G.N.; Steele, P.S.; Demuth, D.R. Glycoprotein 340 in normal human ocular surface tissues and tear film. Infect. Immun. 2006, 74, 4058–4063. [Google Scholar] [CrossRef] [PubMed]
- McDermott, A.M. Antimicrobial compounds in tears. Exp. Eye Res. 2013, 117, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Girgis, D.O.; Dajcs, J.J.; O’Callaghan, R.J. Phospholipase A2 activity in normal and Staphylococcus aureus-infected rabbit eyes. Investig. Ophthalmol. Vis. Sci. 2003, 44, 197–202. [Google Scholar] [CrossRef]
- Tam, C.; Mun, J.J.; Evans, D.J.; Fleiszig, S.M.J. Cytokeratins mediate epithelial innate defense through their antimicrobial properties. J. Clin. Investig. 2012, 122, 3665–3677. [Google Scholar] [CrossRef] [PubMed]
- McDermott, A.M.; Redfern, R.L.; Zhang, B. Human ß-defensin 2 is up-regulated during re-epithelialization of the cornea. Curr. Eye Res. 2001, 22, 64–67. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Zhang, J.; Yu, F.-S.X. Toll-like receptor 2-mediated expression of β-defensin-2 in human corneal epithelial cells. Microbes Infect. 2006, 8, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Shirai, K.; Saika, S. Ocular surface mucins and local inflammation—Studies in genetically modified mouse lines. BMC Ophthalmol. 2015, 15, 154. [Google Scholar] [CrossRef] [PubMed]
- Kardon, R.; Price, R.E.; Julian, J.; Lagow, E.; Tseng, S.C.; Gendler, S.J.; Carson, D.D. Bacterial conjunctivitis in Muc1 null mice. Investig. Ophthalmol. Vis. Sci. 1999, 40, 1328–1335. [Google Scholar]
- Ricciuto, J.; Heimer, S.R.; Gilmore, M.S.; Argüeso, P. Cell Surface O-glycans limit Staphylococcus aureus adherence to corneal epithelial cells. Infect. Immun. 2008, 76, 5215–5220. [Google Scholar] [CrossRef] [PubMed]
- Ormerod, L.D.; Hertzmark, E.; Gomez, D.S.; Stabiner, R.G.; Schanzlin, D.J.; Smith, R.E. Epidemiology of microbial keratitis in Southern California: A multivariate analysis. Ophthalmology 1987, 94, 1322–1333. [Google Scholar] [CrossRef]
- Coster, D.J.; Badenoch, P.R. Host, microbial, and pharmacological factors affecting the outcome of suppurative keratitis. Br. J. Ophthalmol. 1987, 71, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Jett, B.D.; Gilmore, M.S. Internalization of Staphylococcus aureus by human corneal epithelial cells: Role of bacterial fibronectin-binding protein and host cell factors. Infect. Immun. 2002, 70, 4697–4700. [Google Scholar] [CrossRef] [PubMed]
- Rhem, M.N.; Lech, E.M.; Patti, J.M.; McDevitt, D.; Höök, M.; Jones, D.B.; Wilhelmus, K.R. The collagen-binding adhesin is a virulence factor in Staphylococcus aureus keratitis. Infect. Immun. 2000, 68, 3776–3779. [Google Scholar] [CrossRef] [PubMed]
- Alarcon, I.; Kwan, L.; Yu, C.; Evans, D.J.; Fleiszig, S.M.J. Role of the corneal epithelial basement membrane in ocular defense against Pseudomonas aeruginosa. Infect. Immun. 2009, 77, 3264–3271. [Google Scholar] [CrossRef] [PubMed]
- Mootz, J.M.; Benson, M.A.; Heim, C.E.; Crosby, H.A.; Kavanaugh, J.S.; Dunman, P.M.; Kielian, T.; Torres, V.J.; Horswill, A.R. Rot is a key regulator of Staphylococcus aureus biofilm formation. Mol. Microbiol. 2015, 96, 388–404. [Google Scholar] [CrossRef] [PubMed]
- Benson, M.A.; Lilo, S.; Wasserman, G.A.; Thoendel, M.; Smith, A.; Horswill, A.R.; Fraser, J.; Novick, R.P.; Shopsin, B.; Torres, V.J. Staphylococcus aureus regulates the expression and production of the staphylococcal superantigen-like secreted proteins in a Rot-dependent manner. Mol. Microbiol. 2011, 81, 659–675. [Google Scholar] [CrossRef] [PubMed]
- Benson, M.A.; Lilo, S.; Nygaard, T.; Voyich, J.M.; Torres, V.J. Rot and SaeRS cooperate to activate expression of the Staphylococcal superantigen-like exoproteins. J. Bacteriol. 2012, 194, 4355–4365. [Google Scholar] [CrossRef] [PubMed]
- Hume, E.B.H.; Dajcs, J.J.; Moreau, J.M.; Sloop, G.D.; Willcox, M.D.P.; O’Callaghan, R.J. Staphylococcus corneal virulence in a new topical model of infection. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2904–2908. [Google Scholar]
- Girgis, D.O.; Sloop, G.D.; Reed, J.M.; O’Callaghan, R.J. A new topical model of Staphylococcus corneal infection in the mouse. Investig. Ophthalmol. Vis. Sci. 2003, 44, 1591–1597. [Google Scholar] [CrossRef]
- Callegan, M.C.; Engel, L.S.; Hill, J.M.; O’Callaghan, R.J. Corneal virulence of Staphylococcus aureus: Roles of alpha-toxin and protein A in pathogenesis. Infect. Immun. 1994, 62, 2478–2482. [Google Scholar] [PubMed]
- O’Callaghan, R.J.; Callegan, M.C.; Moreau, J.M.; Green, L.C.; Foster, T.J.; Hartford, O.M.; Engel, L.S.; Hill, J.M. Specific roles of alpha-toxin and beta-toxin during Staphylococcus aureus corneal infection. Infect. Immun. 1997, 65, 1571–1578. [Google Scholar] [PubMed]
- Sloop, G.D.; Moreau, J.M.; Conerly, L.L.; Dajcs, J.J.; O’Callaghan, R.J. Acute inflammation of the eyelid and cornea in Staphylococcus keratitis in the rabbit. Investig. Ophthalmol. Vis. Sci. 1999, 40, 385–391. [Google Scholar]
- Girgis, D.; Sloop, G.; Reed, J.; O’Callaghan, R. Susceptibility of aged mice to Staphylococcus aureus keratitis. Curr. Eye Res. 2004, 29, 269–275. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, R.J.; McCormick, C.C.; Caballero, A.R.; Marquart, M.E.; Gatlin, H.P.; Fratkin, J.D. Age-related differences in rabbits during experimental Staphylococcus aureus keratitis. Investig. Ophthalmol. Vis. Sci. 2007, 48, 5125–5131. [Google Scholar] [CrossRef] [PubMed]
- Dajcs, J.J.; Thibodeaux, B.A.; Girgis, D.O.; O’Callaghan, R.J. Corneal virulence of Staphylococcus aureus in an experimental model of keratitis. DNA Cell Biol. 2002, 21, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Moreau, J.M.; Sloop, G.D.; Engel, L.S.; Hill, J.M.; O’Callaghan, R.J. Histopathological studies of staphylococcal alpha-toxin: Effects on rabbit corneas. Curr. Eye Res. 1997, 16, 1221–1228. [Google Scholar] [CrossRef] [PubMed]
- Girgis, D.O.; Sloop, G.D.; Reed, J.M.; O’Callaghan, R.J. Effects of toxin production in a murine model of Staphylococcus aureus keratitis. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2064–2070. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.Z.J.; Zhu, H.; Stapleton, F.; Hume, E.; Aliwarga, Y.; Thakur, A.; Willcox, M.D.P. Effects of α-toxin-deficient Staphylococcus aureus on the production of peripheral corneal ulceration in an animal model. Curr. Eye Res. 2005, 30, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.Z.; Zhu, H.; Thakur, A.; Willcox, M.D. Comparison of potential pathogenic traits of staphylococci that may contribute to corneal ulceration and inflammation. Aust. N. Z. J. Ophthalmol. 1999, 27, 234–236. [Google Scholar] [CrossRef] [PubMed]
- Kebaier, C.; Chamberland, R.R.; Allen, I.C.; Gao, X.; Broglie, P.M.; Hall, J.D.; Jania, C.; Doerschuk, C.M.; Tilley, S.L.; Duncan, J.A. Staphylococcus aureus α-hemolysin mediates virulence in a murine model of severe pneumonia through activation of the NLRP3 Inflammasome. J. Infect. Dis. 2012, 205, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Berube, B.J.; Wardenburg, J.B. Staphylococcus aureus α-toxin: Nearly a century of intrigue. Toxins 2013, 5, 1140–1166. [Google Scholar] [CrossRef] [PubMed]
- Menestrina, G.; Dalla Serra, M.; Prévost, G. Mode of action of β-barrel pore-forming toxins of the staphylococcal α-hemolysin family. Toxicon 2001, 39, 1661–1672. [Google Scholar] [CrossRef]
- Recsei, P.; Kreiswirth, B.; O’Reilly, M.; Schlievert, P.; Gruss, A.; Novick, R.P. Regulation of exoprotein gene expression in Staphylococcus aureus by agr. Mol. Gen. Genet. MGG 1986, 202, 58–61. [Google Scholar] [CrossRef] [PubMed]
- Novick, R.P.; Ross, H.F.; Projan, S.J.; Kornblum, J.; Kreiswirth, B.; Moghazeh, S. Synthesis of staphylococcal virulence factors is controlled by a regulatory RNA molecule. EMBO J. 1993, 12, 3967–3975. [Google Scholar] [PubMed]
- Xiong, Y.Q.; Willard, J.; Yeaman, M.R.; Cheung, A.L.; Bayer, A.S. Regulation of Staphylococcus aureus α-toxin gene (hla) expression by agr, sarA and sae In Vitro and in experimental infective endocarditis. J. Infect. Dis. 2006, 194, 1267–1275. [Google Scholar] [CrossRef] [PubMed]
- Tegmark, K.; Karlsson, A.; Arvidson, S. Identification and characterization of SarH1, a new global regulator of virulence gene expression in Staphylococcus aureus. Mol. Microbiol. 2000, 37, 398–409. [Google Scholar] [CrossRef] [PubMed]
- Berube, B.J.; Sampedro, G.R.; Otto, M.; Wardenburg, J.B. The psmα Locus Regulates production of Staphylococcus aureus alpha-toxin during Infection. Infect. Immun. 2014, 82, 3350–3358. [Google Scholar] [CrossRef] [PubMed]
- McCormick, C.C.; Caballero, A.R.; Balzli, C.L.; Tang, A.; O’Callaghan, R.J. Chemical inhibition of alpha-toxin, a key corneal virulence factor of Staphylococcus aureus. Investig. Ophthalmol. Vis. Sci. 2009, 50, 2848–2854. [Google Scholar] [CrossRef] [PubMed]
- Weeks, A.C.; Balzli, C.L.; Caballero, A.; Tang, A.; O’Callaghan, R. Identification and potency of cyclodextrin-lipid inhibitors of Staphylococcus aureus α-toxin. Curr. Eye Res. 2012, 37, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Hume, E.B.H.; Dajcs, J.J.; Moreau, J.M.; O’Callaghan, R.J. Immunization with alpha-toxin toxoid protects the cornea against tissue damage during experimental Staphylococcus aureus keratitis. Infect. Immun. 2000, 68, 6052–6055. [Google Scholar] [CrossRef] [PubMed]
- Caballero, A.R.; Foletti, D.L.; Bierdeman, M.A.; Tang, A.; Arana, A.M.; Hasa-Moreno, A.; Sangalang, E.R.B.; O’Callaghan, R.J. Effectiveness of alpha-toxin Fab monoclonal antibody therapy in limiting the pathology of Staphylococcus aureus keratitis. Ocul. Immunol. Inflamm. 2015, 23, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, T.; Zaidi, T.; Yoong, P.; Pier, G.B. Staphylococcus aureus corneal infections: Effect of the Panton-Valentine leukocidin (PVL) and antibody to PVL on virulence and pathology. Investig. Ophthalmol. Vis. Sci. 2013, 54, 4430–4438. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, K.; Kawai, Y.; Tanaka, Y.; Hirano, N.; Kaneko, J.; Tomita, N.; Ohta, M.; Kamio, Y.; Yao, M.; Tanaka, I. Crystal structure of the octameric pore of staphylococcal γ-hemolysin reveals the β-barrel pore formation mechanism by two components. Proc. Natl. Acad. Sci. USA 2011, 108, 17314–17319. [Google Scholar] [CrossRef] [PubMed]
- Gravet, A.; Colin, D.A.; Keller, D.; Giradot, R.; Monteil, H.; Prévost, G. Characterization of a novel structural member, LukE-LukD, of the bi-component staphylococcal leucotoxins family. FEBS Lett. 1998, 436, 202–208. [Google Scholar] [CrossRef]
- Szmigielski, S.; Prévost, G.; Monteil, H.; Colin, D.A.; Jeljaszewicz, J. Leukocidal toxins of staphylococci. Zentralblatt Für Bakteriologie 1999, 289, 185–201. [Google Scholar] [CrossRef]
- Sugawara, N.; Tomita, T.; Kamio, Y. Assembly of Staphylococcus aureus γ-hemolysin into a pore-forming ring-shaped complex on the surface of human erythrocytes. FEBS Lett. 1997, 410, 333–337. [Google Scholar] [CrossRef]
- Bierdeman, M.A.; Torres, A.M.; Caballero, A.R.; Tang, A.; O’Callaghan, R.J. Reactions with antisera and pathological effects of Staphylococcus aureus gamma-toxin in the cornea. Curr. Eye Res. 2017, 42, 1100–1107. [Google Scholar] [CrossRef] [PubMed]
- Hesje, C.K.; Sanfilippo, C.M.; Haas, W.; Morris, T.W. Molecular epidemiology of methicillin-resistant and methicillin-susceptible Staphylococcus aureus isolated from the eye. Curr. Eye Res. 2011, 36, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Amato, M.; Pershing, S.; Walvick, M.; Tanaka, S. Trends in ophthalmic manifestations of methicillin-resistant Staphylococcus aureus (MRSA) in a northern California pediatric population. J. Am. Assoc. Pediatr. Ophthalmol. Strabismus 2013, 17, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Mah, F.S.; Davidson, R.; Holland, E.J.; Hovanesian, J.; John, T.; Kanellopoulos, J.; Shamie, N.; Starr, C.; Vroman, D.; Kim, T. Current knowledge about and recommendations for ocular methicillin-resistant Staphylococcus aureus. J. Cataract Refract. Surg. 2014, 40, 1894–1908. [Google Scholar] [CrossRef] [PubMed]
- Asbell, P.A.; Sanfilippo, C.M.; Pillar, C.M.; DeCory, H.H.; Sahm, D.F.; Morris, T.W. Antibiotic resistance among ocular pathogens in the United States: Five-year results from the antibiotic resistance monitoring in ocular microorganisms (ARMOR) Surveillance Study. JAMA Ophthalmol. 2015, 133, 1445–1454. [Google Scholar] [CrossRef] [PubMed]
- Sueke, H.; Shankar, J.; Neal, T.; Winstanley, C.; Tuft, S.; Coates, R.; Horsburgh, M.J.; Kaye, S. lukSF-PV in Staphylococcus aureus keratitis isolates and association with clinical outcome. Investig. Ophthalmol. Vis. Sci. 2013, 54, 3410–3416. [Google Scholar] [CrossRef] [PubMed]
- Jett, B.D.; Gilmore, M.S. Host-parasite interactions in Staphylococcus aureus keratitis. DNA Cell Biol. 2002, 21, 397–404. [Google Scholar] [CrossRef]
- Zaidi, T.; Zaidi, T.; Cywes-Bentley, C.; Lu, R.; Priebe, G.P.; Pier, G.B. Microbiota-driven immune cellular maturation is essential for antibody-mediated adaptive immunity to Staphylococcus aureus infection in the eye. Infect. Immun. 2014, 82, 3483–3491. [Google Scholar] [CrossRef] [PubMed]
- Zegans, M.E.; Becker, H.I.; Budzik, J.; O’Toole, G. The role of bacterial biofilms in ocular infections. DNA Cell Biol. 2002, 21, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Sun, X.; Wang, Z.; Zhang, Y. Biofilm-forming capacity of Staphylococcus epidermidis, Staphylococcus aureus, and Pseudomonas aeruginosa from ocular infections. Investig. Ophthalmol. Vis. Sci. 2012, 53, 5624–5631. [Google Scholar] [CrossRef] [PubMed]
- Artini, M.; Cellini, A.; Scoarughi, G.L.; Papa, R.; Tilotta, M.; Palma, S.; Selan, L. Evaluation of contact lens multipurpose solutions on bacterial biofilm development. Eye Contact Lens 2015, 41, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Traidej, M.; Caballero, A.R.; Marquart, M.E.; Thibodeaux, B.A.; O’Callaghan, R.J. Molecular analysis of Pseudomonas aeruginosa protease IV expressed in Pseudomonas putida. Investig. Ophthalmol. Vis. Sci. 2003, 44, 190–196. [Google Scholar] [CrossRef]
- Thibodeaux, B.A.; Caballero, A.R.; Marquart, M.E.; Tommassen, J.; O’Callaghan, R.J. Corneal virulence of Pseudomonas aeruginosa elastase B and alkaline protease produced by Pseudomonas putida. Curr. Eye Res. 2007, 32, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Mondino, B.J.; Chou, H.J.; Sumner, H.L. Generation of complement membrane attack complex in normal human corneas. Investig. Ophthalmol. Vis. Sci. 1996, 37, 1576–1581. [Google Scholar]
- Ruan, X.; Chodosh, J.; Callegan, M.C.; Booth, M.C.; Lee, T.D.; Kumar, P.; Gilmore, M.S.; Pereira, H.A. Corneal expression of the inflammatory mediator CAP37. Investig. Ophthalmol. Vis. Sci. 2002, 43, 1414–1421. [Google Scholar]
- Hume, E.B.H.; Cole, N.; Garthwaite, L.L.; Khan, S.; Willcox, M.D.P. A protective role for IL-6 in Staphylococcal microbial keratitis. Investig. Ophthalmol. Vis. Sci. 2006, 47, 4926–4930. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhang, R.; Gadek, T.R.; O’Neill, C.A.; Pearlman, E. Corneal iinflammation is inhibited by the LFA-1 antagonist, lifitegrast (SAR 1118). J. Ocul. Pharmacol. Ther. 2012, 29, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Scott, I.U.; Flynn, H.W.; Feuer, W.; Pflugfelder, S.C.; Alfonso, E.C.; Forster, R.K.; Miller, D. Endophthalmitis associated with microbial keratitis. Ophthalmology 1996, 103, 1864–1870. [Google Scholar] [CrossRef]
- O’Neill, E.C.; Yeoh, J.; Fabinyi, D.C.A.; Cassidy, D.; Vajpayee, R.B.; Allen, P.; Connell, P.P. Risk factors, microbial profiles and prognosis of microbial keratitis-associated endophthalmitis in high-risk eyes. Graefes Arch. Clin. Exp. Ophthalmol. 2014, 252, 1457–1462. [Google Scholar] [CrossRef] [PubMed]
- The Endophthalmitis Vitrectomy Study Group. Microbiologic factors and visual outcome in the endophthalmitis vitrectomy study. Am. J. Ophthalmol. 1996, 122, 830–846. [Google Scholar] [CrossRef]
- Gentile, R.C.; Shukla, S.; Shah, M.; Ritterband, D.C.; Engelbert, M.; Davis, A.; Hu, D.-N. Microbiological spectrum and antibiotic sensitivity in endophthalmitis: A 25-Year Review. Ophthalmology 2014, 121, 1634–1642. [Google Scholar] [CrossRef] [PubMed]
- Valdez-García, J.E.; Climent, A.; Chávez-Mondragón, E.; Lozano-Ramírez, J.F. Anterior chamber bacterial contamination in cataract surgery. BMC Ophthalmol. 2014, 14, 57. [Google Scholar] [CrossRef] [PubMed]
- Shimada, H.; Arai, S.; Nakashizuka, H.; Hattori, T.; Yuzawa, M. Reduced anterior chamber contamination by frequent surface irrigation with diluted iodine solutions during cataract surgery. Acta Ophthalmol. 2017, 95, e373–e378. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Lenci, L.T.; Reddy, C.V.; Russell, S.R. Postoperative hemorrhagic occlusive retinal vasculitis associated with intracameral vancomycin prophylaxis during cataract surgery. J. Cataract Refract. Surg. 2016, 42, 1676–1680. [Google Scholar] [CrossRef] [PubMed]
- Witkin, A.J.; Shah, A.R.; Engstrom, R.E.; Kron-Gray, M.M.; Baumal, C.R.; Johnson, M.W.; Witkin, D.I.; Leung, J.; Albini, T.A.; Moshfeghi, A.A.; et al. Postoperative hemorrhagic occlusive retinal vasculitis: Expanding the clinical spectrum and possible association with vancomycin. Ophthalmology 2015, 122, 1438–1451. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, S.G.; Flynn, H.W.; Das, T.; Mieler, W.F. Ocular Infection: Endophthalmitis. Dev. Ophthalmol. 2016, 55, 176–188. [Google Scholar] [CrossRef] [PubMed]
- Booth, M.C.; Hatter, K.L.; Miller, D.; Davis, J.; Kowalski, R.; Parke, D.W.; Chodosh, J.; Jett, B.D.; Callegan, M.C.; Penland, R.; et al. Molecular epidemiology of Staphylococcus aureus and Enterococcus faecalis in endophthalmitis. Infect. Immun. 1998, 66, 356–360. [Google Scholar] [PubMed]
- Callegan, M.C.; Booth, M.C.; Jett, B.D.; Gilmore, M.S. Pathogenesis of gram-positive bacterial endophthalmitis. Infect. Immun. 1999, 67, 3348–3356. [Google Scholar] [PubMed]
- Arana, A.M.; Bierdeman, M.A.; Balzli, C.L.; Tang, A.; Caballero, A.R.; Patel, R.; O’Callaghan, R.J. Staphylococcus alpha-toxin action on the rabbit iris: Toxic effects and their inhibition. Curr. Eye Res. 2015, 40, 830–838. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kumar, A. Role of Staphylococcus aureus virulence factors in inducing inflammation and vascular permeability in a mouse model of bacterial endophthalmitis. PLoS ONE 2015, 10, e0128423. [Google Scholar] [CrossRef] [PubMed]
- Booth, M.C.; Atkuri, R.V.; Nanda, S.K.; Iandolo, J.J.; Gilmore, M.S. Accessory gene regulator controls Staphylococcus aureus virulence in endophthalmitis. Investig. Ophthalmol. Vis. Sci. 1995, 36, 1828–1836. [Google Scholar]
- Booth, M.C.; Cheung, A.L.; Hatter, K.L.; Jett, B.D.; Callegan, M.C.; Gilmore, M.S. Staphylococcal accessory regulator (sar) in conjunction with agr contributes to Staphylococcus aureus virulence in endophthalmitis. Infect. Immun. 1997, 65, 1550–1556. [Google Scholar] [PubMed]
- Giese, M.J.; Berliner, J.A.; Riesner, A.; Wagar, E.A.; Mondino, B.J. A comparison of the early inflammatory effects of an agr-/sar- versus a wild type strain of Staphylococcus aureus in a rat model of endophthalmitis. Curr. Eye Res. 1999, 18, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Sadaka, A.; Palmer, K.; Suzuki, T.; Gilmore, M.S. In vitro and in vivo models of Staphylococcus aureus endophthalmitis implicate specific nutrients in ocular infection. PLoS ONE 2014, 9, e110872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Supersac, G.; Piémont, Y.; Kubina, M.; Prévost, G.; Foster, T.J. Assessment of the role of gamma-toxin in experimental endophthalmitis using ahlg-deficient mutant of Staphylococcus aureus. Microb. Pathog. 1998, 24, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, J.A.; Speeg-Schatz, C.; Freitas, F.I.S.; Sahel, J.; Monteil, H.; Prévost, G. Channel-forming leucotoxins from Staphylococcus aureus cause severe inflammatory reactions in a rabbit eye model. J. Med. Microbiol. 1997, 46, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Bronner, S.; Jehl, F.; Peter, J.-D.; Ploy, M.-C.; Renault, C.; Arvis, P.; Monteil, H.; Prevost, G. Moxifloxacin efficacy and vitreous penetration in a rabbit model of Staphylococcus aureus endophthalmitis and effect on gene expression of leucotoxins and virulence regulator factors. Antimicrob. Agents Chemother. 2003, 47, 1621–1629. [Google Scholar] [CrossRef] [PubMed]
- Whiston, E.A.; Sugi, N.; Kamradt, M.C.; Sack, C.; Heimer, S.R.; Engelbert, M.; Wawrousek, E.F.; Gilmore, M.S.; Ksander, B.R.; Gregory, M.S. αB-Crystallin Protects Retinal Tissue during Staphylococcus aureus-Induced Endophthalmitis. Infect. Immun. 2008, 76, 1781–1790. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Kumar, A. Mitochondria mediates caspase-dependent and independent retinal cell death in Staphylococcus aureus endophthalmitis. Cell Death Discov. 2016, 2, 16034. [Google Scholar] [CrossRef] [PubMed]
- Lawin-Brüssel, C.A.; Refojo, M.F.; Kenyon, K.R. In vitro adhesion of Pseudomonas aeruginosa and Staphylococcus aureus to surface passivated poly(methyl methacrylate) intraocular lenses. J. Cataract Refract. Surg. 1992, 18, 598–601. [Google Scholar] [CrossRef]
- Portolés, M.; Refojo, M.F.; Leong, F.-L. Reduced bacterial adhesion to heparin-surface-modified intraocular lenses. J. Cataract Refract. Surg. 1993, 19, 755–759. [Google Scholar] [CrossRef]
- Doyle, A.; Beigi, B.; Early, A.; Blake, A.; Eustace, P.; Hone, R. Adherence of bacteria to intraocular lenses: A prospective study. Br. J. Ophthalmol. 1995, 79, 347–349. [Google Scholar] [CrossRef] [PubMed]
- Leid, J.G.; Costerton, J.W.; Shirtliff, M.E.; Gilmore, M.S.; Engelbert, M. Immunology of Staphylococcal biofilm infections in the eye: New tools to study biofilm endophthalmitis. DNA Cell Biol. 2002, 21, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Kohn, A.N. Bacterial cultures of lenses removed during cataract surgery. Am. J. Ophthalmol. 1978, 86, 162–166. [Google Scholar] [CrossRef]
- Beyer, T.L.; Vogler, G.; Sharma, D.; O’Donnell, F.E. Protective barrier effect of the posterior lens capsule in exogenous bacterial endophthalmitis—An experimental primate study. Investig. Ophthalmol. Vis. Sci. 1984, 25, 108–112. [Google Scholar] [CrossRef]
- Beyer, T.L.; O’Donnell, F.E.; Goncalves, V.; Singh, R. Role of the posterior capsule in the prevention of postoperative bacterial endophthalmitis: Experimental primate studies and clinical implications. Br. J. Ophthalmol. 1985, 69, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Streilein, J.W.; Ohta, K.; Mo, J.S.; Taylor, A.W. Ocular immune privilege and the impact of intraocular inflammation. DNA Cell Biol. 2002, 21, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Girgis, D.O.; Reed, J.M.; Monds, K.S.; Dajcs, J.J.; Marquart, M.E.; Thibodeaux, B.A.; O’Callaghan, R.J. Pathogenesis of Staphylococcus in the rabbit anterior chamber. Investig. Ophthalmol. Vis. Sci. 2005, 46, 1371–1378. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, R.P.; Romanowski, E.G.; Mah, F.S.; Sasaki, H.; Fukuda, M.; Gordon, Y.J. A comparison of moxifloxacin and levofloxacin topical prophylaxis in a fluoroquinolone-resistant Staphylococcus aureus rabbit model. Jpn. J. Ophthalmol. 2008, 52, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Chen, H.; Jiang, H.; Xu, Y.; Liu, T.; Xu, L. Prophylactic effect of topical fluoroquinolones in a rabbit model of Staphylococcus aureus endophthalmitis. J. Ocul. Pharmacol. Ther. 2012, 28, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Balzli, C.L.; Bartell, J.; Dajcs, J.J.; McCormick, C.C.; Caballero, A.R.; Stroman, D.; O’Callaghan, R.J. A Highly Virulent Staphylococcus aureus: Rabbit anterior chamber infection, characterization, and genetic analysis. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5114–5120. [Google Scholar] [CrossRef] [PubMed]
- Diamond, J.P.; Leeming, J.P.; Smart, A.D.; Duffus, P.; Hedges, A.J.; Easty, D.L. An antimicrobial effect associated with rabbit primary aqueous humour. Br. J. Ophthalmol. 1994, 78, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Diamond, D.J.P.; Moule, K.; Leeming, J.P.; Tavare, J.; Easty, D.L. Purification of an antimicrobial peptide from rabbit aqueous humour. Curr. Eye Res. 1998, 17, 783–787. [Google Scholar] [CrossRef] [PubMed]
- Perumal, N.; Manicam, C.; Steinicke, M.; Funke, S.; Pfeiffer, N.; Grus, F.H. Characterization of the human aqueous humour proteome: A comparison of the genders. PLoS ONE 2017, 12, e0172481. [Google Scholar] [CrossRef] [PubMed]
- Mondino, B.J.; Sidikaro, Y.; Sumner, H. Anaphylatoxin levels in human vitreous humor. Investig. Ophthalmol. Vis. Sci. 1988, 29, 1195–1198. [Google Scholar]
- Stainer, G.A.; Peyman, G.A.; Berkowitz, R.; Tessler, H.H. Intraocular lysozyme in experimental uveitis in rabbits: Aqueous and vitreous assay. Investig. Ophthalmol. Vis. Sci. 1976, 15, 312–315. [Google Scholar]
- Bernatchez, S.F.; Tabatabay, C.; Belin, D. Urokinase-type plasminogen activator in human aqueous humor. Investig. Ophthalmol. Vis. Sci. 1992, 33, 2687–2692. [Google Scholar]
- Suzuki, T.; Campbell, J.; Swoboda, J.G.; Walker, S.; Gilmore, M.S. Role of wall teichoic acids in Staphylococcus aureus endophthalmitis. Investig. Ophthalmol. Vis. Sci. 2011, 52, 3187–3192. [Google Scholar] [CrossRef] [PubMed]
- Aizuss, D.H.; Mondino, B.J.; Sumner, H.L.; Dethlefs, B.A. The complement system and host defense against Pseudomonas endophthalmitis. Investig. Ophthalmol. Vis. Sci. 1985, 26, 1262–1266. [Google Scholar]
- Giese, M.J.; Mondino, B.J.; Glasgow, B.J.; Sumner, H.L.; Adamu, S.A.; Halabi, H.P.; Chou, H.J. Complement system and host defense against staphylococcal endophthalmitis. Investig. Ophthalmol. Vis. Sci. 1994, 35, 1026–1032. [Google Scholar]
- Haynes, R.J.; Tighe, P.J.; Dua, H.S. Antimicrobial defensin peptides of the human ocular surface. Br. J. Ophthalmol. 1999, 83, 737–741. [Google Scholar] [CrossRef] [PubMed]
- Yamagata, M.; Rook, S.L.; Sassa, Y.; Ma, R.C.; Geraldes, P.; Goddard, L.; Clermont, A.; Gao, B.; Salti, H.; Gundel, R.; et al. Bactericidal/permeability-increasing protein’s signaling pathways and its retinal trophic and anti-angiogenic effects. FASEB J. 2006, 20, 2058–2067. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Kumar, A. Retinal photoreceptor expresses Toll-like receptors (TLRs) and elicits innate responses following TLR ligand and bacterial challenge. PLoS ONE 2015, 10, e0119541. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Pandey, R.K.; Miller, L.J.; Singh, P.K.; Kanwar, M. Muller Glia in Retinal innate immunity: A perspective on their roles in endophthalmitis. Crit. Rev. Immunol. 2013, 33. [Google Scholar] [CrossRef]
- Talreja, D.; Singh, P.K.; Kumar, A. In vivo role of TLR2 and MyD88 signaling in eliciting innate immune responses in Staphylococcal endophthalmitis. Investig. Ophthalmol. Vis. Sci. 2015, 56, 1719–1732. [Google Scholar] [CrossRef] [PubMed]
- Rajamani, D.; Singh, P.K.; Rottmann, B.G.; Singh, N.; Bhasin, M.K.; Kumar, A. Temporal retinal transcriptome and systems biology analysis identifies key pathways and hub genes in Staphylococcus aureus endophthalmitis. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Reviglio, V.E.; Sambuelli, R.H.; Olmedo, A.; Falco, M.; Echenique, J.; O’Brien, T.P.; Kuo, I.C. Secretory leukocyteprotease inhibitor is an inducible antimicrobial peptide expressed in Staphylococcus aureus endophthalmitis. Mediat. Inflamm. 2007, 2007. [Google Scholar] [CrossRef] [PubMed]
- Giese, M.J.; Sumner, H.L.; Berliner, J.A.; Mondino, B.J. Cytokine expression in a rat model of Staphylococcus aureus endophthalmitis. Investig. Ophthalmol. Vis. Sci. 1998, 39, 2785–2790. [Google Scholar]
- Rosenzweig, H.L.; Galster, K.T.; Planck, S.R.; Rosenbaum, J.T. NOD1 expression in the eye and functional contribution to IL-1β–dependent ocular inflammation in Mice. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1746–1753. [Google Scholar] [CrossRef] [PubMed]
- Giese, M.J.; Shum, D.C.; Rayner, S.A.; Mondino, B.J.; Berliner, J.A. Adhesion molecule expression in a rat model of Staphylococcus aureus endophthalmitis. Investig. Ophthalmol. Vis. Sci. 2000, 41, 145–153. [Google Scholar]
- Kumar, A.; Singh, C.N.; Glybina, I.V.; Mahmoud, T.H.; Yu, F.-S.X. Toll-like receptor 2 ligand—Induced protection against bacterial endophthalmitis. J. Infect. Dis. 2010, 201, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Kochan, T.; Singla, A.; Tosi, J.; Kumar, A. Toll-Like receptor 2 ligand pretreatment attenuates retinal microglial inflammatory response but enhances phagocytic activity toward Staphylococcus aureus. Infect. Immun. 2012, 80, 2076–2088. [Google Scholar] [CrossRef] [PubMed]
- Sugi, N.; Whiston, E.A.; Ksander, B.R.; Gregory, M.S. Increased resistance to Staphylococcus aureus endophthalmitis in BALB/c Mice: Fas ligand is required for resolution of inflammation but not for bacterial clearance. Infect. Immun. 2013, 81, 2217–2225. [Google Scholar] [CrossRef] [PubMed]
- Engelbert, M.; Gilmore, M.S. Fas ligand but not complement is critical for control of experimental Staphylococcus aureus endophthalmitis. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2479–2486. [Google Scholar] [CrossRef] [PubMed]
- Han, D.P. Intravitreal human immune globulin in a rabbit model of Staphylococcus aureus toxin-mediated endophthalmitis: A potential adjunct in the treatment of endophthalmitis. Trans. Am. Ophthalmol. Soc. 2004, 102, 305–320. [Google Scholar] [PubMed]
- Perkins, S.L.; Han, D.P.; Burke, J.M.; Schlievert, P.M.; Wirostko, W.J.; Tarasewicz, D.G.; Skumatz, C.M.B. Intravitreally injected human immunoglobulin attenuates the effects of Staphylococcus aureus culture supernatant in a rabbit model of toxin-mediated endophthalmitis. Arch. Ophthalmol. 2004, 122, 1499–1506. [Google Scholar] [CrossRef] [PubMed]
- Laventie, B.-J.; Rademaker, H.J.; Saleh, M.; de Boer, E.; Janssens, R.; Bourcier, T.; Subilia, A.; Marcellin, L.; van Haperen, R.; Lebbink, J.H.G.; et al. Heavy chain-only antibodies and tetravalent bispecific antibody neutralizing Staphylococcus aureus leukotoxins. Proc. Natl. Acad. Sci. USA 2011, 108, 16404–16409. [Google Scholar] [CrossRef] [PubMed]
- Ravindranath, R.M.; Mondino, B.J.; Adamu, S.A.; Pitchekian-Halabi, H.; Hasan, S.A.; Glasgow, B.J. Immunopathologic features of Staphylococcus aureus endophthalmitis in the rat. Investig. Ophthalmol. Vis. Sci. 1995, 36, 2482–2491. [Google Scholar]
- Engstrom, R.E.; Mondino, B.J.; Glasgow, B.J.; Pitchekian-Halabi, H.; Adamu, S.A. Immune response to Staphylococcus aureus endophthalmitis in a rabbit model. Investig. Ophthalmol. Vis. Sci. 1991, 32, 1523–1533. [Google Scholar]
- Giese, M.J.; Rayner, S.A.; Fardin, B.; Sumner, H.L.; Rozengurt, N.; Mondino, B.J.; Gordon, L.K. Mitigation of neutrophil infiltration in a rat model of early Staphylococcus aureus endophthalmitis. Investig. Ophthalmol. Vis. Sci. 2003, 44, 3077–3082. [Google Scholar] [CrossRef]
- Coburn, P.S.; Wiskur, B.J.; Astley, R.A.; Callegan, M.C. Blood–retinal barrier compromise and endogenous Staphylococcus aureus endophthalmitis. Investig. Ophthalmol. Vis. Sci. 2015, 56, 7303–7311. [Google Scholar] [CrossRef] [PubMed]
- Coburn, P.S.; Wiskur, B.J.; Miller, F.C.; LaGrow, A.L.; Astley, R.A.; Elliott, M.H.; Callegan, M.C. Bloodstream-to-eye infections are facilitated by outer blood-retinal barrier dysfunction. PLoS ONE 2016, 11, e0154560. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Aqueous Layer Protein | Activity of the Protein |
|---|---|
| Lysozyme | Comprises 20–30% of the protein; enzymatically cleaves cell wall peptidoglycan killing the bacteria, but not that of S. aureus [35] |
| Lactoferrin | Comprises 20–30% of the protein; binds iron, limiting bacterial growth and its cationic detergent effect can lyse bacteria [35,46] |
| Lipochalin | Binds iron, limiting bacterial growth [47] |
| Complement | Complement system in tears is functionally active [48] |
| Secretory phospholipase A2 | Lipolytic enzyme activity on phospholipids of bacterial membranes is lethal for gram-positive bacteria; a major defense against S. aureus [49] |
| Secretory leukocyte protease inhibitor (SLPI) | Cationic charge can bind bacterial membranes killing the organism and can inhibit some bacterial proteases [50] |
| Surfactants A and D | Bind and aggregate bacteria favoring their phagocytosis [51,52] |
| Glycoprotein 340 | Binds bacteria favoring their phagocytosis [53] |
| Antimicrobial peptides | Have bactericidal effects [54] |
| Eye Protections to Be Avoided by Bacteria | Bacterial Binding Proteins | Epithelial Cell Response | Inflammation and Tissue Damage Mediators | |
|---|---|---|---|---|
| Tear Film | Mucous Layer | |||
| PLA2 Mucins Surfactant Protein Immunoglobulin Leukocytes Complement Cytokines AMP’s SLPI Lactoferrin Lipochalin | Mucins Leukocytes Immunoglobulins Cytokines | Fibronectin Binding Protein Collagen Binding Protein | IL-1α IL-1β IL-6 IL-8 IL-17 TNF-α LL-37 | Teichoic Acid Alpha-Toxin Gamma-Toxin PVL ** SSL ** S. aureus elastase ** |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
O’Callaghan, R.J. The Pathogenesis of Staphylococcus aureus Eye Infections. Pathogens 2018, 7, 9. https://doi.org/10.3390/pathogens7010009
O’Callaghan RJ. The Pathogenesis of Staphylococcus aureus Eye Infections. Pathogens. 2018; 7(1):9. https://doi.org/10.3390/pathogens7010009
Chicago/Turabian StyleO’Callaghan, Richard J. 2018. "The Pathogenesis of Staphylococcus aureus Eye Infections" Pathogens 7, no. 1: 9. https://doi.org/10.3390/pathogens7010009
APA StyleO’Callaghan, R. J. (2018). The Pathogenesis of Staphylococcus aureus Eye Infections. Pathogens, 7(1), 9. https://doi.org/10.3390/pathogens7010009