Genome Characterization of a Pathogenic Porcine Rotavirus B Strain Identified in Buryat Republic, Russia in 2015

 ,
,
Abstract
:1. Introduction
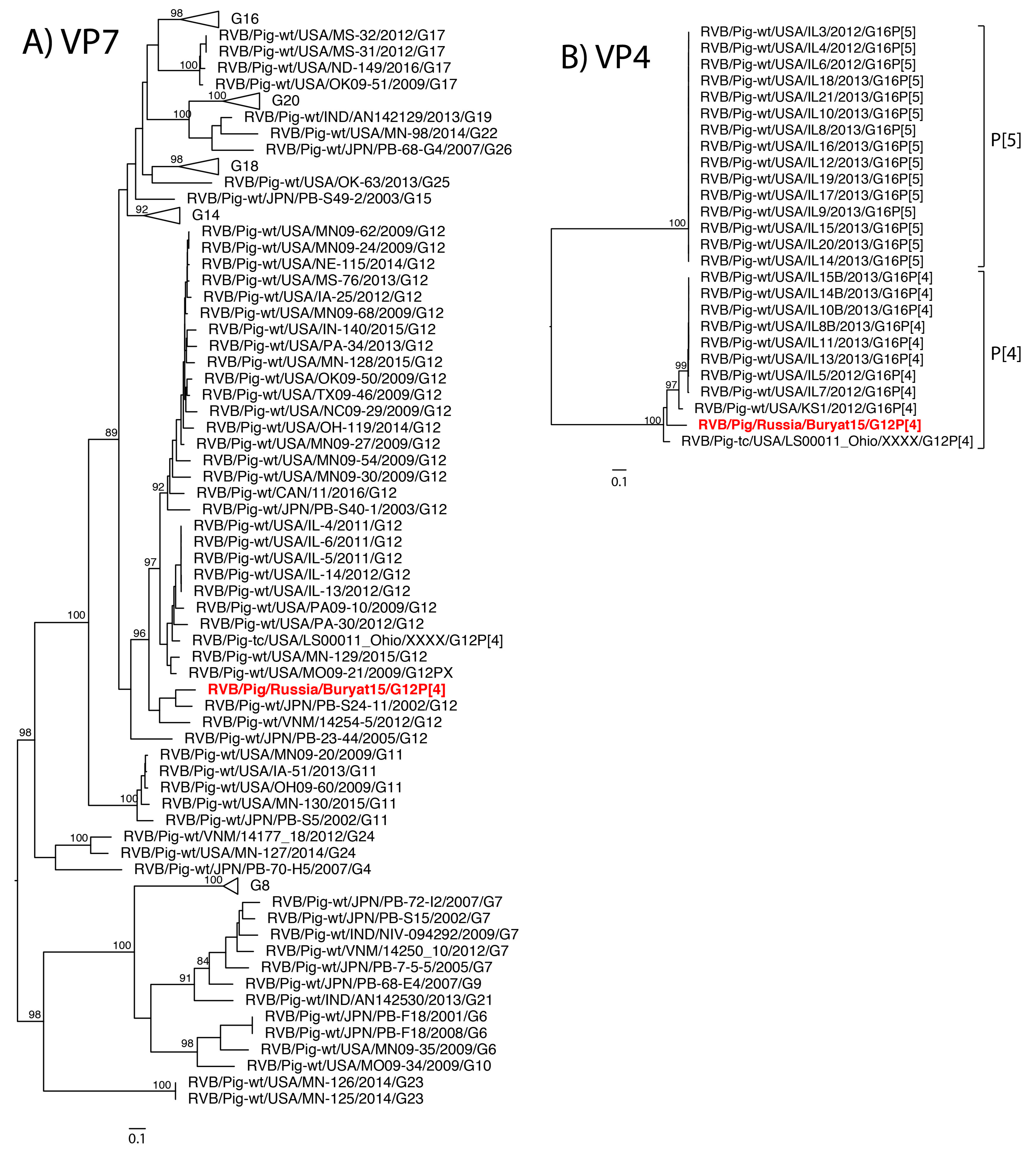
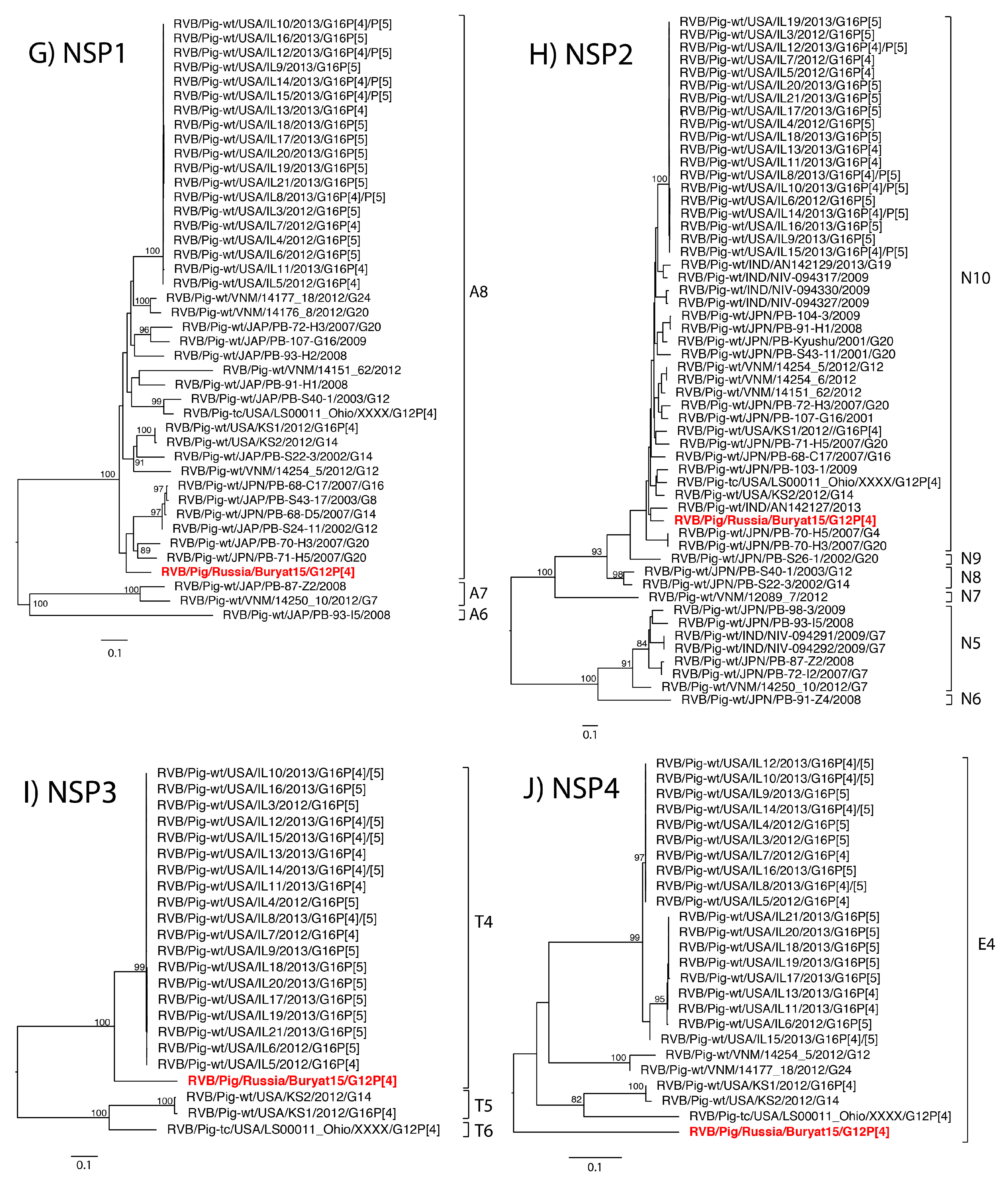
2. Results
3. Discussion
4. Materials and Methods
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bishop, R.; Davidson, G.P.; Holmes, I.H.; Ruck, B.J. Virus particles in epithelial cells of duodenal mucosa from children with acute non-bacterial gastroenteritis. Lancet 1973, 302, 1281–1283. [Google Scholar] [CrossRef]
- Flewett, T.H.; Bryden, A.S.; Davies, H. Virus particles in gastroenteritis. Lancet 1973, 2, 1497. [Google Scholar] [CrossRef]
- Rodger, S.M.; Craven, J.A.; Williams, I. Demonstration of Reovirus-like Particles in Intestinal Contents of Piglets with Diarrhoea. Aust. Vet. J. 1975, 51, 536. [Google Scholar] [CrossRef] [PubMed]
- Flewett, T.H.; Bryden, A.S.; Davies, H.; Woode, G.N.; Bridger, J.C.; Derrick, J.M. Relation between viruses from acute gastroenteritis of children and newborn calves. Lancet 1974, 304, 61–63. [Google Scholar] [CrossRef]
- Flewett, T.H.; Woode, G.N. The rotaviruses. Arch. Virol. 1978, 57, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Janke, B.H.; Nelson, J.K.; Benfield, D.A.; Nelson, E.A. Relative prevalence of typical and atypical strains among rotaviruses from diarrheic pigs in conventional swine herds. J. Vet. Diagn. Investig. 1990, 2, 308–311. [Google Scholar] [CrossRef] [PubMed]
- Estes, M.K.; Greenberg, H.B. Rotaviruses. In Fields Virology, 6th ed.; Wolters Kluwer Health/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 1347–1401. [Google Scholar]
- Matthijnssens, J.; Otto, P.H.; Ciarlet, M.; Desselberger, U.; Van Ranst, M.; Johne, R. VP6-sequence-based cutoff values as a criterion for rotavirus species demarcation. Arch. Virol. 2012, 157, 1177–1182. [Google Scholar] [CrossRef] [PubMed]
- Bányai, K.; Kemenesi, G.; Budinski, I.; Földes, F.; Zana, B.; Marton, S.; Varga-Kugler, R.; Oldal, M.; Kurucz, K.; Jakab, F. Candidate new rotavirus species in Schreiber’s bats, Serbia. Infect. Genet. Evol. 2016, 48, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Mihalov-Kovács, E.; Gellért, Á.; Marton, S.; Farkas, S.L.; Fehér, E.; Oldal, M.; Jakab, F.; Martella, V.; Bányai, K. Candidate new rotavirus species in sheltered dogs, Hungary. Emerg. Infect. Dis. 2015, 21, 660–663. [Google Scholar] [CrossRef] [PubMed]
- Marthaler, D.; Homwong, N.; Rossow, K.; Culhane, M.; Goyal, S.; Collins, J.; Matthijnssens, J.; Ciarlet, M. Rapid detection and high occurrence of porcine rotavirus A, B, and C by RT-qPCR in diagnostic samples. J. Virol. Methods 2014, 209, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Marthaler, D.; Rossow, K.; Culhane, M.; Goyal, S.; Collins, J.; Matthijnssens, J.; Nelson, M.; Ciarlet, M. Widespread Rotavirus H in Commercially Raised Pigs, United States. Emerg. Infect. Dis. 2014, 20, 1203–1206. [Google Scholar] [CrossRef] [PubMed]
- Pedley, S.; Bridger, J.C.; Chasey, D.; McCrae, M.A. Definition of two new groups of atypical rotaviruses. J. Gen. Virol. 1986, 67 Pt 1, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Saif, L.J.; Bohl, E.H.; Theil, K.W.; Cross, R.F.; House, J.A. Rotavirus-like, calicivirus-like, and 23-nm virus-like particles associated with diarrhea in young pigs. J. Clin. Microbiol. 1980, 12, 105–111. [Google Scholar] [PubMed]
- Bridger, J.C.; Brown, J.F. Prevalence of antibody to typical and atypical rotaviruses in pigs. Vet. Rec. 1985, 116, 50. [Google Scholar] [CrossRef] [PubMed]
- Theil, K.W.; Saif, L.J.; Moorhead, P.D.; Whitmoyer, R.E. Porcine Rotavirus-Like Virus (Group B Rotavirus): Characterization and Pathogenicity for Gnotobiotic Pigs. J. Clin. Microbiol. 1985, 21, 340–345. [Google Scholar] [PubMed]
- Homwong, N.; Diaz, A.; Rossow, S.; Ciarlet, M.; Marthaler, D. Three-Level Mixed-Effects Logistic Regression Analysis Reveals Complex Epidemiology of Swine Rotaviruses in Diagnostic Samples from North America. PLoS ONE 2016, 11, e0154734. [Google Scholar] [CrossRef] [PubMed]
- Kuga, K.; Miyazaki, A.; Suzuki, T.; Takagi, M.; Hattori, N.; Katsuda, K.; Mase, M.; Sugiyama, M.; Tsunemitsu, H. Genetic diversity and classification of the outer capsid glycoprotein VP7 of porcine group B rotaviruses. Arch. Virol. 2009, 154, 1785. [Google Scholar] [CrossRef] [PubMed]
- Otto, P.H.; Rosenhain, S.; Elschner, M.C.; Hotzel, H.; Machnowska, P.; Trojnar, E.; Hoffmann, K.; Johne, R. Detection of rotavirus species A, B and C in domestic mammalian animals with diarrhoea and genotyping of bovine species A rotavirus strains. Vet. Microbiol. 2015, 179, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Lahon, A.; Ingle, V.C.; Birade, H.S.; Raut, C.G.; Chitambar, S.D. Molecular characterization of group B rotavirus circulating in pigs from India: Identification of a strain bearing a novel VP7 genotype, G21. Vet. Microbiol. 2014, 174, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Molinari, B.L.D.; Possatti, F.; Lorenzetti, E.; Alfieri, A.F.; Alfieri, A.A. Unusual outbreak of post-weaning porcine diarrhea caused by single and mixed infections of rotavirus groups A, B, C, and H. Vet. Microbiol. 2016, 193, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Geyer, A.; Sebata, T.; Peenze, I.; Steele, A.D. Group B and C porcine rotaviruses identified for the first time in South Africa. J. S. Afr. Vet. Assoc. 1996, 67, 115–116. [Google Scholar] [PubMed]
- Madson, D. Comparative Analysis: The Pathogenesis of Disease Induced in Piglets by Group A, B and C Rotaviruses, Singularly or Concurrently. Available online: https://www.pork.org/research/comparative-analysis-the-pathogenesis-of-disease-induced-in-piglets-by-group-a-b-and-c-rotaviruses-singularly-or-concurrently/ (accessed on 1 January 2013).
- Shepherd, F.K.; Murtaugh, M.P.; Chen, F.; Culhane, M.R.; Marthaler, D.G. Longitudinal Surveillance of Porcine Rotavirus B Strains from the United States and Canada and In Silico Identification of Antigenically Important Sites. Pathogens 2017, 6, 64. [Google Scholar] [CrossRef] [PubMed]
- Zhirakovskaia, E.; Tikunov, A.; Klemesheva, V.; Loginovskikh, N.; Netesov, S.; Tikunova, N. First genetic characterization of rotavirus C in Russia. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2016, 39, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Soma, J.; Miyazaki, A.; Tsunemitsu, H. Phylogenetic analysis of nonstructural protein 5 (NSP5) gene sequences in porcine rotavirus B strains. Infect. Genet. Evol. 2012, 12, 1661–1668. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Soma, J.; Kuga, K.; Miyazaki, A.; Tsunemitsu, H. Sequence and phylogenetic analyses of nonstructural protein 2 genes of species B porcine rotaviruses detected in Japan during 2001–2009. Virus Res. 2012, 165, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Marthaler, D.; Rossow, K.; Gramer, M.; Collins, J.; Goyal, S.; Tsunemitsu, H.; Kuga, K.; Suzuki, T.; Ciarlet, M.; Matthijnssens, J. Detection of substantial porcine group B rotavirus genetic diversity in the United States, resulting in a modified classification proposal for G genotypes. Virology 2012, 433, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Bwogi, J.; Jere, K.C.; Karamagi, C.; Byarugaba, D.K.; Namuwulya, P.; Baliraine, F.N.; Desselberger, U.; Iturriza-Gomara, M. Whole genome analysis of selected human and animal rotaviruses identified in Uganda from 2012 to 2014 reveals complex genome reassortment events between human, bovine, caprine and porcine strains. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Aung, M.S.; Nahar, S.; Aida, S.; Paul, S.K.; Hossain, M.A.; Ahmed, S.; Haque, N.; Ghosh, S.; Malik, Y.S.; Urushibara, N.; et al. Distribution of two distinct rotavirus B (RVB) strains in the north-central Bangladesh and evidence for reassortment event among human RVB revealed by whole genomic analysis. Infect. Genet. Evol. 2017, 47, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Marthaler, D.; Suzuki, T.; Rossow, K.; Culhane, M.; Collins, J.; Goyal, S.; Tsunemitsu, H.; Ciarlet, M.; Matthijnssens, J. VP6 genetic diversity, reassortment, intragenic recombination and classification of rotavirus B in American and Japanese pigs. Vet. Microbiol. 2014, 172, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Hasebe, A.; Miyazaki, A.; Tsunemitsu, H. Analysis of genetic divergence among strains of porcine rotavirus C, with focus on VP4 and VP7 genotypes in Japan. Virus Res. 2015, 197, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinform. Oxf. Engl. 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Predicted Epitope Location | 33 | 34 | 36 | 37 | 39 | 40 | 65 | 66 | 67 | 89 | 90 | 91 | 92 | 130 | 158 | 159 | 160 | 161 | 179 | 180 | 181 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Consensus | D | D | N | D | K | Q | X | N | Y | K | Y | A | Y | D | P | D | R | R | S | N | N |
| Russia/Buryat15/2015 | . | . | . | . | . | . | N | . | . | . | . | E | . | . | . | . | . | . | . | . | . |
| JPN/PB-S24-11/2002 | . | . | . | . | E | . | D | . | . | . | . | E | . | . | . | . | . | . | . | . | . |
| JPN/PB-S40-1/2003 | . | . | T | E | . | . | V | S | . | . | . | . | . | . | . | . | . | . | . | . | . |
| CAN/11/2016 | . | . | T | . | . | . | D | Q | . | . | . | . | . | . | . | N | . | . | . | S | . |
| USA/PA-30/2012 | . | . | T | . | . | K | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| USA/MN-129/2015 | . | . | T | . | . | K | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| USA/MN-128/2015 | . | . | . | . | . | . | E | . | . | . | . | . | . | . | . | N | . | . | . | . | . |
| USA/IL-13/2012 | . | . | T | . | . | K | N | D | . | . | . | . | . | . | . | . | . | . | . | . | . |
| USA/IL-5/2011 | . | . | T | . | . | K | N | D | . | . | . | . | . | . | . | . | . | . | . | . | . |
| USA/IL-6/2011 | . | . | T | . | . | K | N | D | . | . | . | . | . | . | . | . | . | . | . | . | . |
| USA/IL-4/2011 | . | . | T | . | . | K | N | D | . | . | . | . | . | . | . | . | . | . | . | . | . |
| USA/IL-14/2012 | . | . | T | . | . | K | N | D | . | . | . | . | . | . | . | . | . | . | . | . | . |
| USA/IN-140/2015 | . | . | . | . | . | . | G | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| USA/OH-119/2014 | . | . | . | . | . | . | Q | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| USA/PA-34/2013 | . | . | . | . | . | . | E | . | . | . | . | . | . | . | . | N | . | . | . | . | . |
| USA/IA-25/2012 | . | . | . | . | . | . | D | . | . | . | . | . | . | . | . | N | . | . | . | . | . |
| USA/NE-115/2014 | . | . | . | . | . | . | E | . | . | . | . | . | . | . | . | N | . | . | . | . | . |
| USA/MS-76/2013 | . | . | . | . | . | . | E | . | . | . | . | . | . | . | . | N | . | . | . | . | . |
| USA/PA09-10/2009 | . | . | T | . | . | K | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| USA/MO09-21/2009 | . | . | T | . | . | K | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| USA/OK09-50/2009 | . | . | . | . | . | . | E | . | . | . | . | . | . | . | . | N | . | . | . | . | . |
| USA/MN09-54/2009 | . | . | . | . | . | . | Q | . | . | . | . | . | . | . | . | . | . | . | . | S | . |
| USA/MN09-30/2009 | . | . | . | . | . | . | I | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| USA/MN09-27/2009 | . | . | . | . | . | . | Q | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| USA/MN09-24/2009 | . | . | . | . | . | . | E | . | . | . | . | . | . | . | . | N | . | . | . | . | . |
| USA/MN09-68/2009 | . | . | . | . | . | . | E | . | . | . | . | . | . | . | . | N | . | . | . | . | . |
| USA/LS00011_Ohio/XXXX | . | . | T | . | . | K | S | D | . | . | . | . | . | . | . | . | . | . | . | . | . |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alekseev, K.P.; Penin, A.A.; Mukhin, A.N.; Khametova, K.M.; Grebennikova, T.V.; Yuzhakov, A.G.; Moskvina, A.S.; Musienko, M.I.; Raev, S.A.; Mishin, A.M.; et al. Genome Characterization of a Pathogenic Porcine Rotavirus B Strain Identified in Buryat Republic, Russia in 2015. Pathogens 2018, 7, 46. https://doi.org/10.3390/pathogens7020046
Alekseev KP, Penin AA, Mukhin AN, Khametova KM, Grebennikova TV, Yuzhakov AG, Moskvina AS, Musienko MI, Raev SA, Mishin AM, et al. Genome Characterization of a Pathogenic Porcine Rotavirus B Strain Identified in Buryat Republic, Russia in 2015. Pathogens. 2018; 7(2):46. https://doi.org/10.3390/pathogens7020046
Chicago/Turabian StyleAlekseev, Konstantin P., Aleksey A. Penin, Alexey N. Mukhin, Kizkhalum M. Khametova, Tatyana V. Grebennikova, Anton G. Yuzhakov, Anna S. Moskvina, Maria I. Musienko, Sergey A. Raev, Alexandr M. Mishin, and et al. 2018. "Genome Characterization of a Pathogenic Porcine Rotavirus B Strain Identified in Buryat Republic, Russia in 2015" Pathogens 7, no. 2: 46. https://doi.org/10.3390/pathogens7020046
APA StyleAlekseev, K. P., Penin, A. A., Mukhin, A. N., Khametova, K. M., Grebennikova, T. V., Yuzhakov, A. G., Moskvina, A. S., Musienko, M. I., Raev, S. A., Mishin, A. M., Kotelnikov, A. P., Verkhovsky, O. A., Aliper, T. I., Nepoklonov, E. A., Herrera-Ibata, D. M., Shepherd, F. K., & Marthaler, D. G. (2018). Genome Characterization of a Pathogenic Porcine Rotavirus B Strain Identified in Buryat Republic, Russia in 2015. Pathogens, 7(2), 46. https://doi.org/10.3390/pathogens7020046