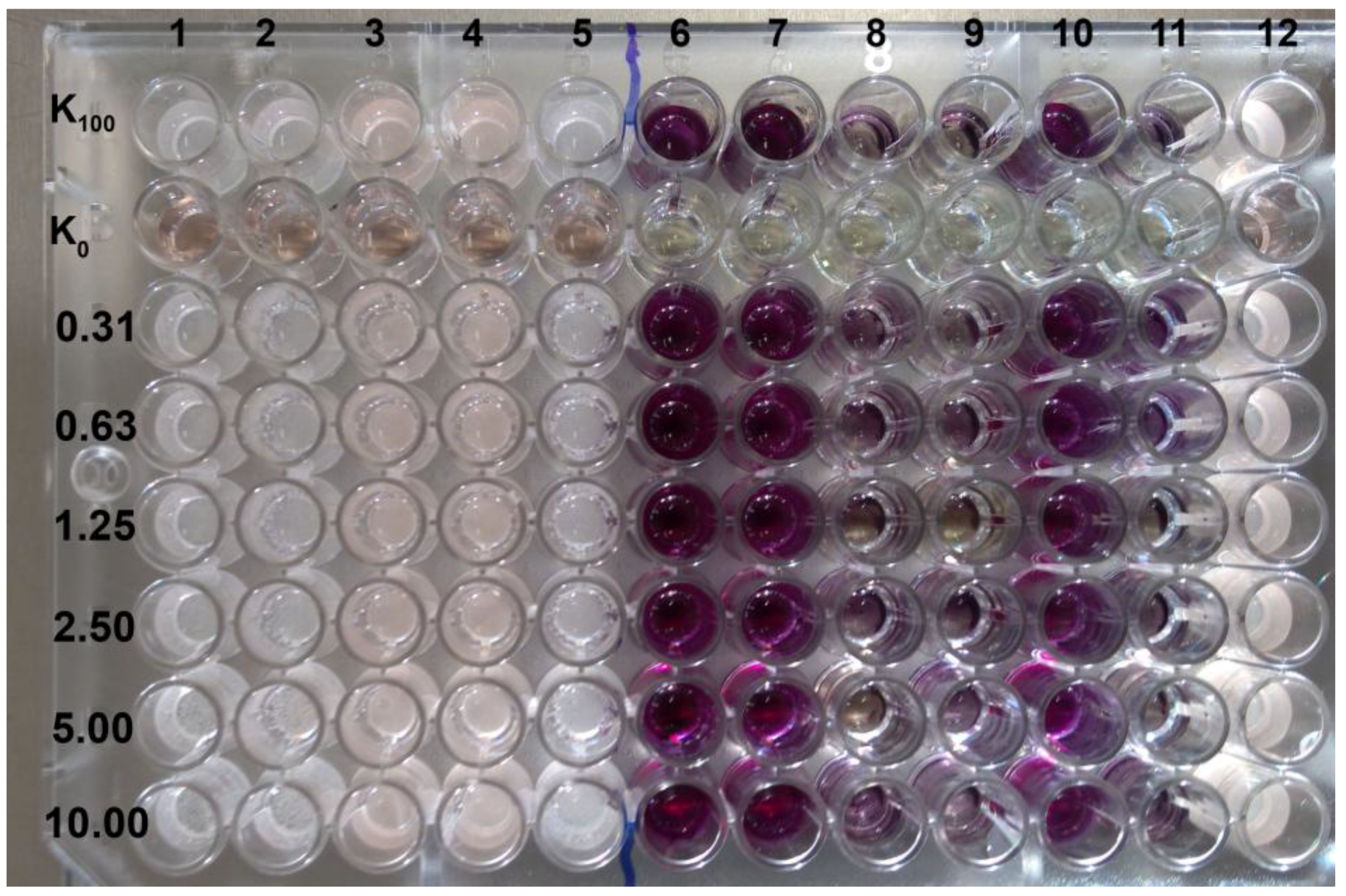
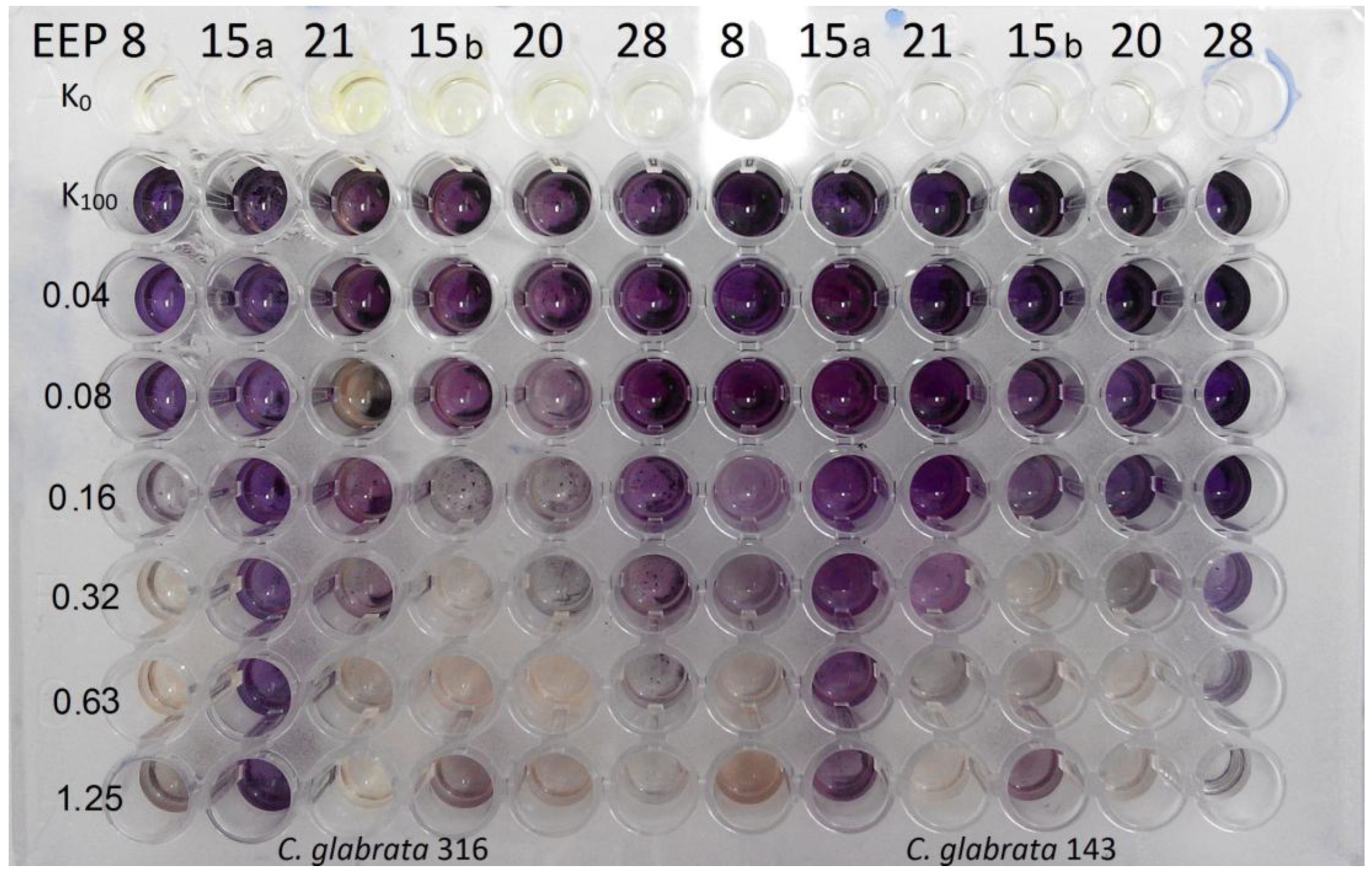
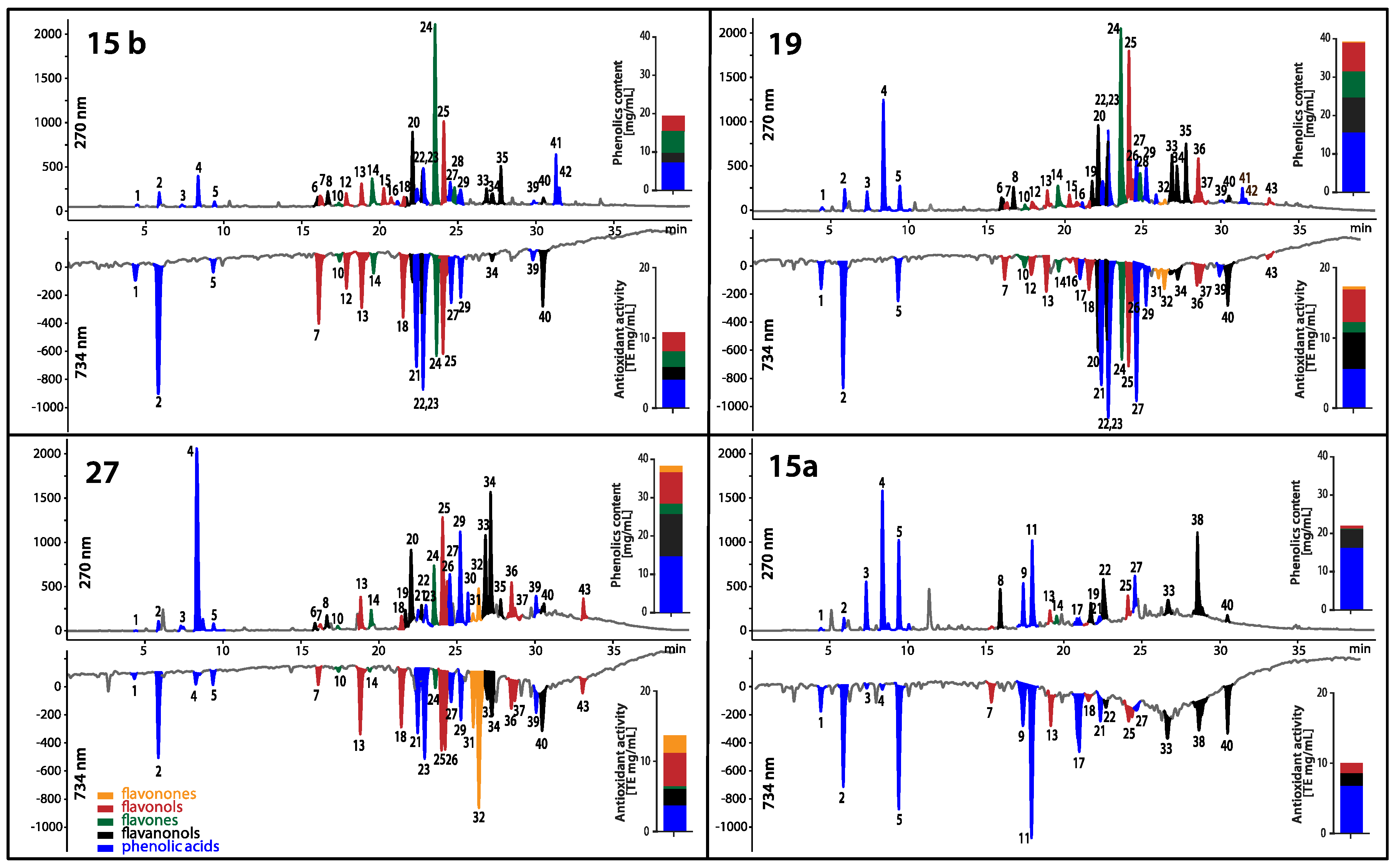
Searching for any new ways of improvement of current antifungal therapy is the main goal of many scientific teams in the world. Natural products continue to be an inexhaustible resource of compounds of potent antimicrobial activity. In line with this tendency, we have focused our attention on ethanolic extracts of honey bee propolis (EEPs), which is known for its antimicrobial activity without serious side effects. Our previous research confirmed a high antifungal potential of propolis produced in some Polish apiaries [
13]; however, a very limited number of propolis samples (
n = 4, collected in the south part of Poland) and
Candida spp. strains were investigated in that study. Herein, we examined activity of 50 samples of propolis obtained from different geographical origins in northern Poland against a large collection of
C. albicans clinical isolates. The preliminary, screening tests revealed that definitely most of the investigated propolis samples exhibited promising activity against
C. albicans and
C. glabrata reference strains. However, some important differences in activity were observed among the samples tested. Some of them, eight in the case of
C. albicans ATCC 10231 and three in the case of
C. glabrata DSM 11226, were able to kill (according to MFC value) the human pathogenic yeasts of the
Candida genus at the concentration 0.31%
v/
v, whilst 7 and 19 EEPs, respectively, were not able to eliminate these strains respectively at the highest tested concentration (2.5%
v/
v). Moreover, slightly higher resistance was observed in the case of
C. albicans strains B4 and Gu5 (fluconazole resistant, overproducing efflux pumps) in comparison to the corresponding strains B3 and Gu4 (fluconazole sensitive, not overproducing efflux pumps). This phenomenon was not observed in our previous study and could suggest that some important active ingredients of propolis can be effectively eliminated from the yeasts’ cells overproducing drug efflux transporters. This issue still needs explanation and is a subject of our current research. The activity of six selected (the most active) EEPs was additionally tested against the group of 69
C. albicans clinical isolates. Most of them were killed at concentrations of 0.31 or 0.63%
v/
v, and resistance to the concentration of 2.5 % (
v/
v) was observed only in the case of one strain for only one EEP. High antifungal potential and some differences in the activity of propolis samples (due to differences of its chemical composition) collected in different regions of The World were also provided by other researchers. For example, Siqueira and coworkers stated that fungicidal activities of Brazilian red propolis were in the range of concentrations of 64–512 μg/mL for
C. albicans and and 64–256 μg/mL for
C. glabrata [
14]. The group of Tobaldini investigated activity of Brazilian green propolis and found that MIC values for this product ranged from 220 to 880 μg/mL [
15]. High antifungal potential of propolis collected in Brazil was also revealed by other authors [
16,
17,
18]. In addition, the groups of Santos [
19,
20] and Pina [
21] successfully treated oral candidiasis with extracts of Brazilian propolis. Haghdoost reported activity of Iran propolis for which MIC and MFC values against
C. albicans isolates were 360.6 μg/mL and 1250.1 μg/mL, respectively [
22]. The MFC values for ethanolic extracts of propolis collected in South Africa varied from 147 to 3125 μg/mL [
23]. On the other hand, Massaro stated that propolis from eastern Australia was not active against
C. albicans ATCC 10231 [
24]. Taking into account the method of preparing the EEPs in our studies (extraction of dry, raw material with 70% ethanol at 3:1 (
v/
w), and about 20% of the raw material was removed as an insoluble waste), the mass concentration of 0.31% (
v/
v) solution (equal to MIC of the most active products) can be estimated as about 630 μg/mL. Thus, the activity of the propolis collected in Polish apiaries is satisfactory and comparable to the activity of the products from other geographical regions.
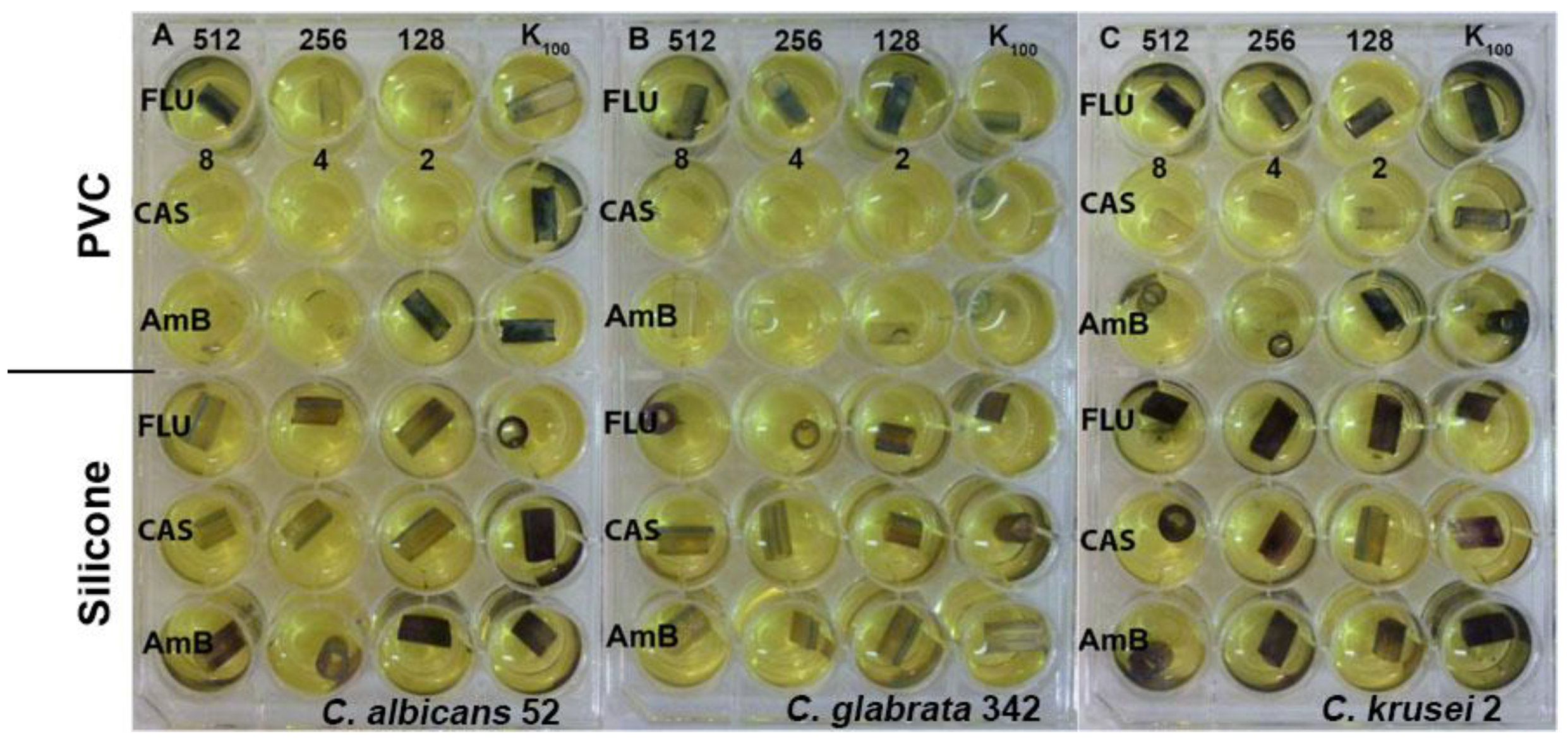
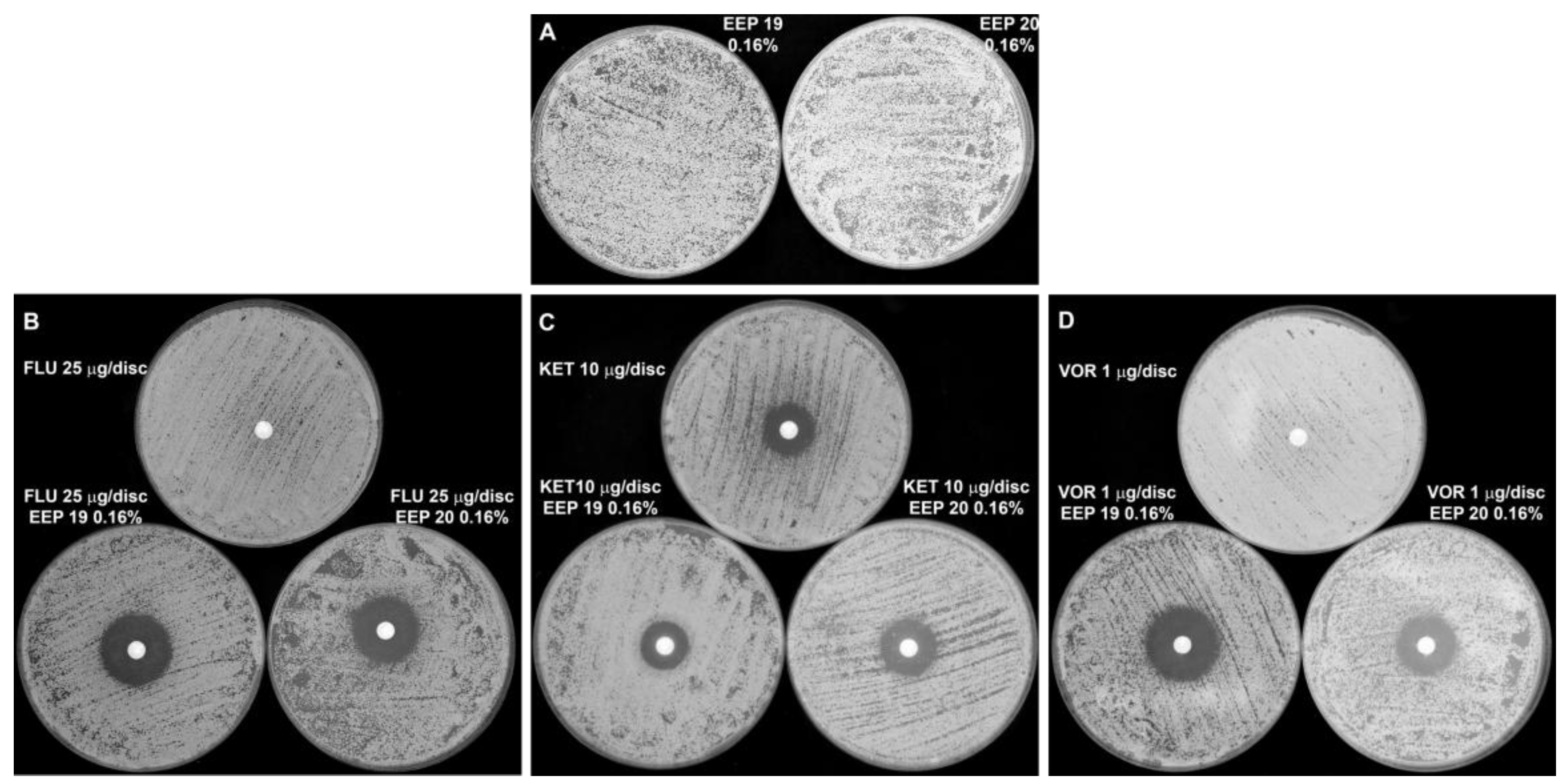
We also investigated the activity of our EEPs in combination with known antimycotics. Searching for a possible synergistic combination is very important because overuse of antimicrobial drugs often leads to resistance and the need for using higher and higher doses of antifungals, which can involve serious side effects. The problem of drug resistance is especially common for fluconazole, which is considered the least toxic of the currently used antifungals and thus the most extensively used. Combinational therapy with a natural compound possesses the ability to reverse fluconazole resistance and can overcome fluconazole incompetence. The synergistic activity of Brazilian propolis with fluconazole was observed by Pippi and coworkers [
17]. In this study, we were able to demonstrate a synergistic effect of Polish propolis in combination with fluconazole and voriconazole. Not evident potentiation of ketoconazole activity was observed. However, this agent alone exhibited quite high activity with a relatively large zone of growth inhibition, which could cover the influence of EEP components. In addition, we proved that addition of EEP in very small concentrations, not exceeding 0.04%, can reduce the effective dose of the fluconazole up to 256 times. Another important remark, which we observed in our study, was that the addition of EEP to the medium allowed elimination of the so called ‘trailing growth’. The term ‘trailing’ has been used to describe the reduced but persistent growth that some isolates of
Candida spp. exhibit at drug concentrations above the MIC in broth dilution tests with azole antifungal agents [
25]. This phenomenon is the most prevalent in
C. albicans isolates. According to Zomorodian et al., trailing growth appears in around 90% of isolates exposed to the activity of ketoconazole, itraconazole or fluconazole and in 78% of isolates exposed to voriconazole. Occasionally, it was also recorded for
C. tropicalis and
C. dubliniensis [
26]. The existence of trailing growth can be the cause of recurrent infections ant treatment failure, thus the use of EEPs in combined therapy would be reasonable.
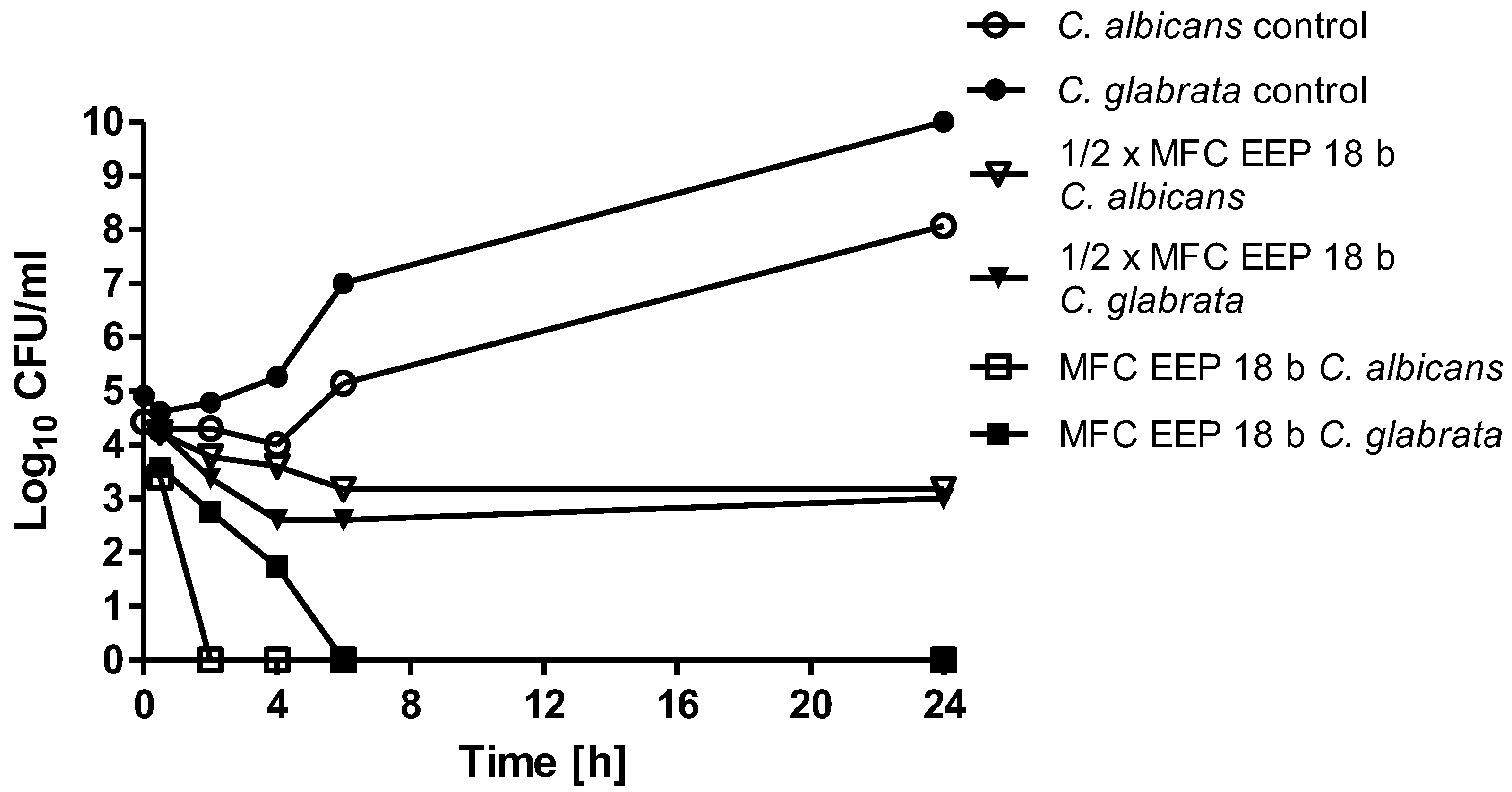
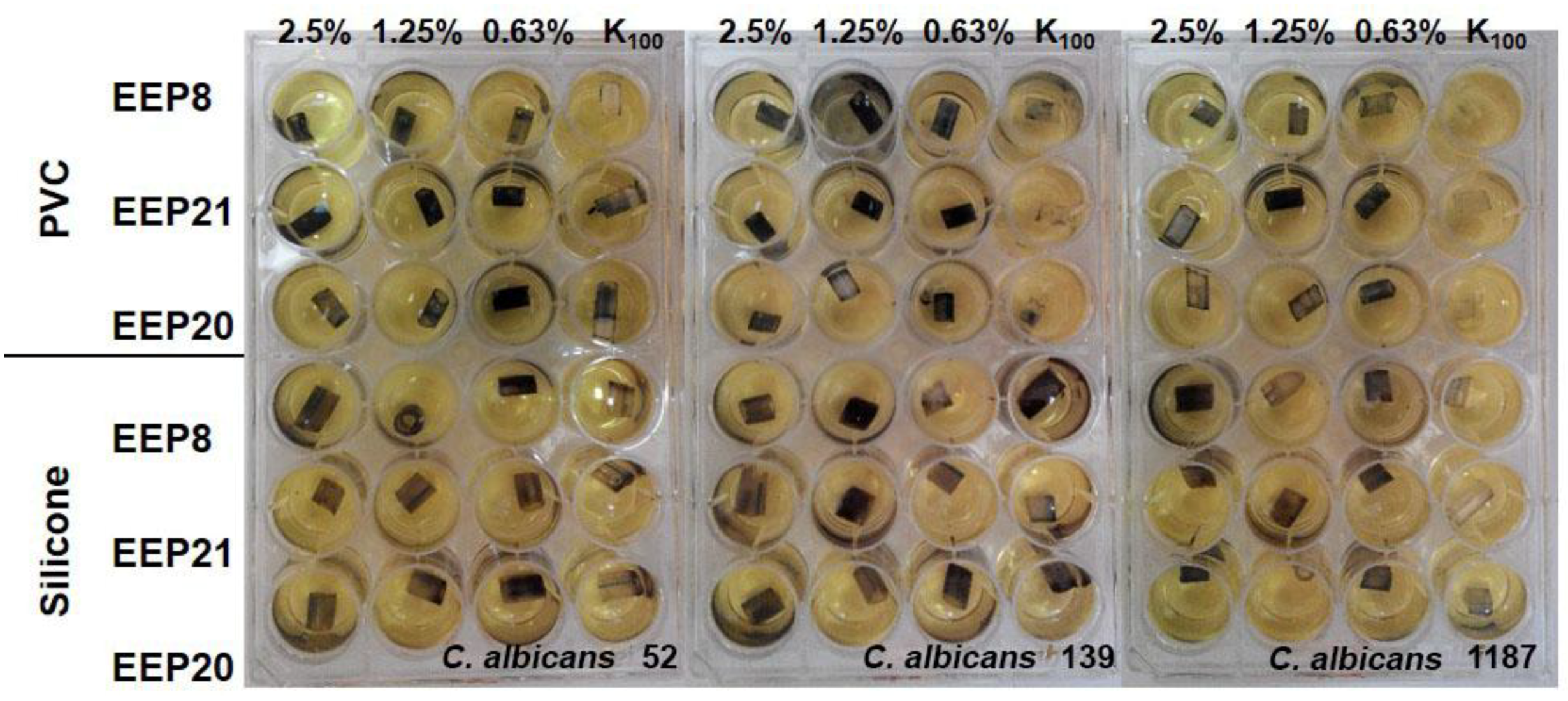
Biofilm formation by
Candida spp. is a key factor in the survival of these cells in the host and is responsible for colonization of tissues and indwelling devices. The possible mechanisms of biofilm resistance to antimicrobial agents include impeded drug penetration through the extracellular structure, phenotypic switching and induction of the expression of resistance genes [
27]. In our research, we revealed that eradication of biofilm in at least 50% requires EEPs concentrations up to 5% (
v/
v) (in individual cases higher). This confirms the conclusions drawn by other researchers about higher resistance of cells living in a biofilm matrix. Capoci et al. [
28] tested activity of Brazilian propolis extracts toward 29 clinical isolates of
C. albicans. They stated that concentration of EEP corresponding to ½× MIC (273.43 μg/mL) was able to reduce biofilm formation in 26% to 95%, depending on the isolate type [
19]. Freires and coworkers revealed that EEPs disrupted biofilm structures of
Candida spp. at a concentration of 500 μg/mL [
18]. Thus, propolis seems to be a promising preparation in the treatment of fungal biofilm.
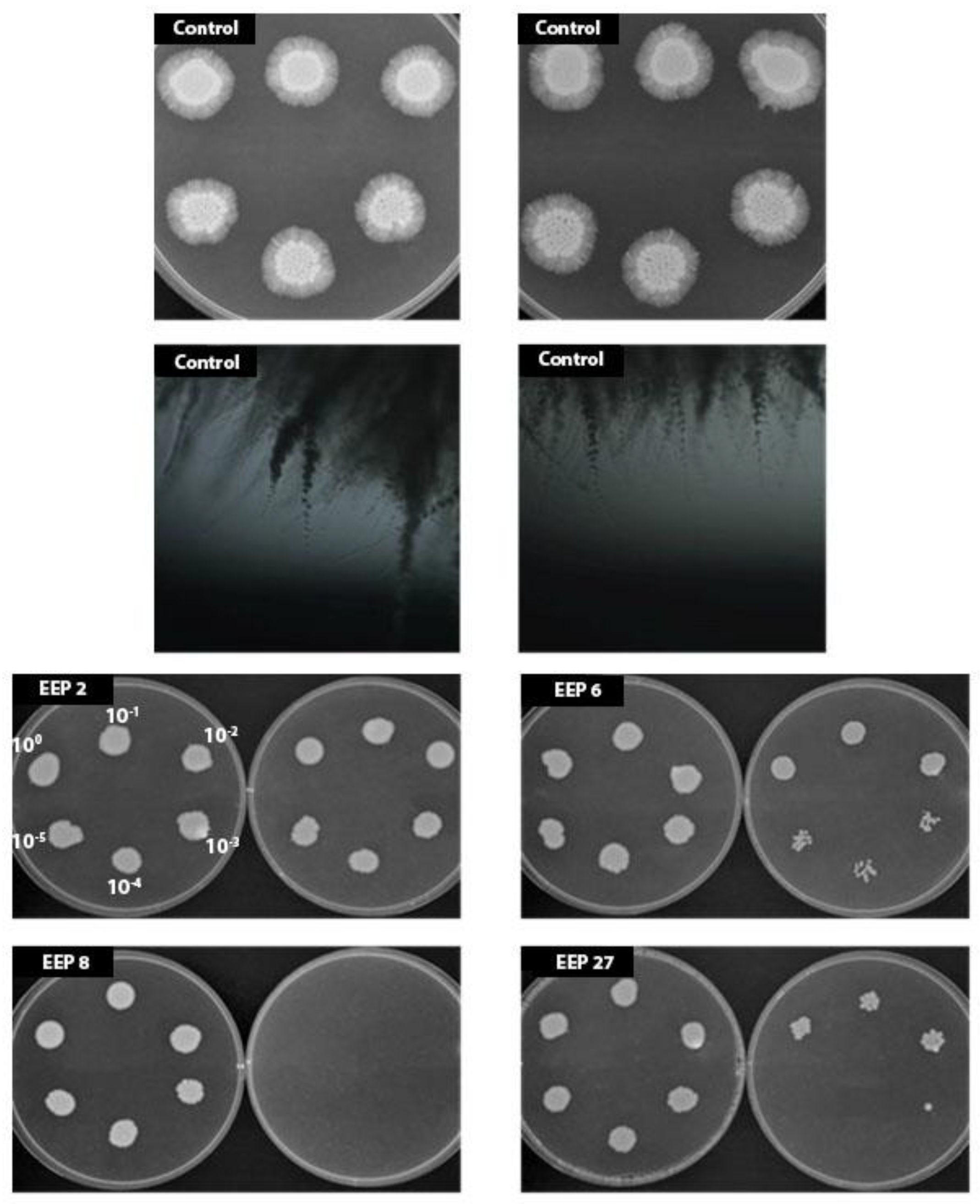
Another important phenomenon, phenotypic switching, is common for
C. albicans strains, among which hyphal growth is the most common cause of invasive fungal infections in humans. Morphological plasticity is its defining feature and is critical for its pathogenesis. Unlike other fungal pathogens that exist primarily in either yeast or hyphal forms,
C. albicans is able to switch reversibly between yeast and hyphal growth forms in response to environmental cues. Nutrient limitation is a known cue that can stimulate yeast-to-hypha morphogenesis at elevated temperatures [
29]. Our results indicate that propolis extracts efficiently inhibit hyphal growth at sublethal concentration in both nutrition limitation culture media. This observation is in agreement with the results of Haghdoots and coworkers who revealed high efficiency of Iranian propolis in reduction of germ tube formation by
C. albicans isolates [
22].
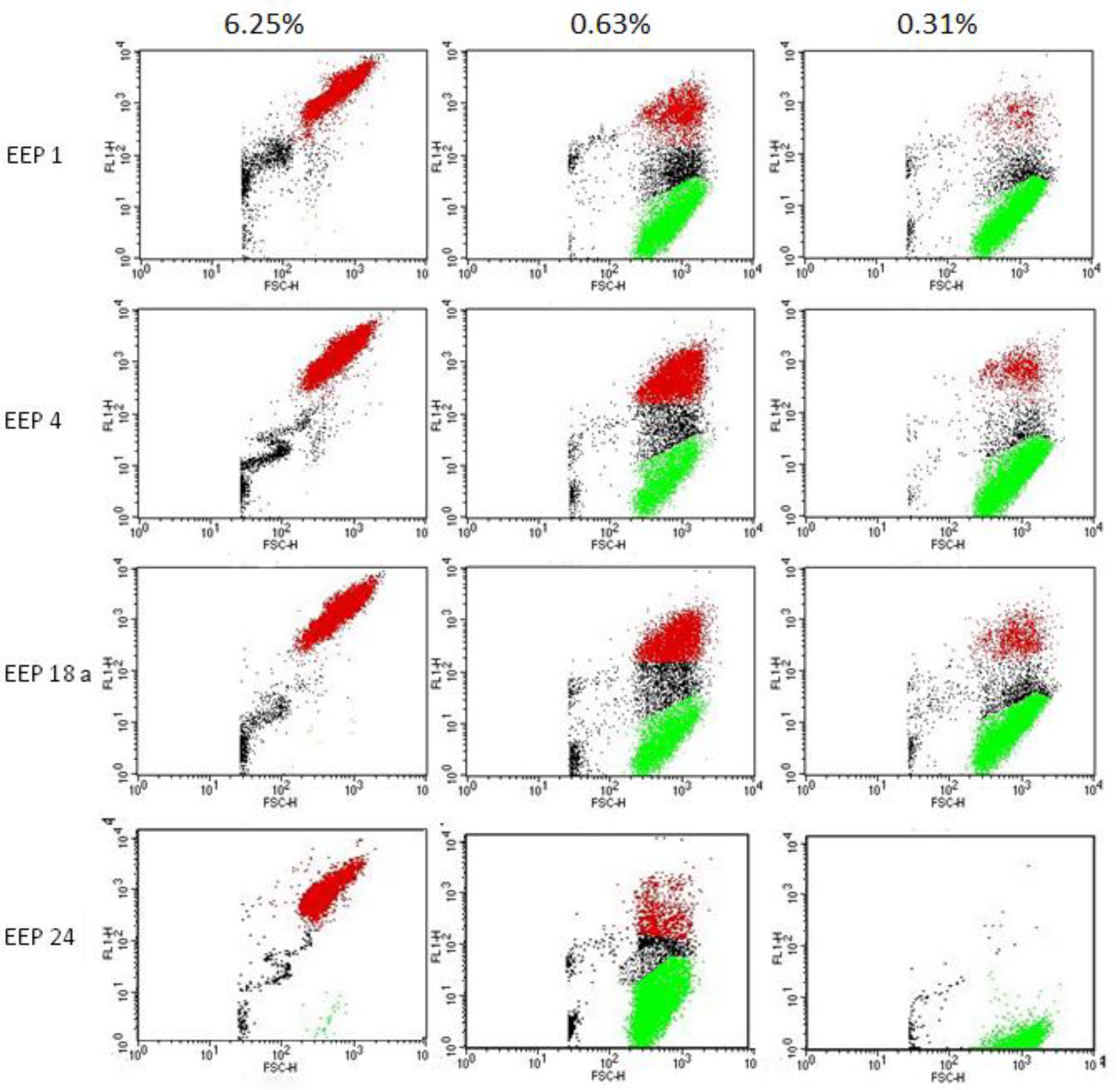
Our studies provided some novel information on possible mode of action of EEPs. Reversal of this action by ergosterol and results of experiment aiming in investigation of membrane depolarization suggested that the cell membrane may be a possible site of action of EEPs components. Experiment with sorbitol as osmoprotectant agent suggests that the cell wall is not a target for ingredients of this product or is a target of secondary importance. However, the detailed mechanism of antifungal activity of EEPs remains unclear and is a subject of our current research.

 ,
, 











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}