Relation between Biofilm and Virulence in Vibrio tapetis: A Transcriptomic Study

 , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
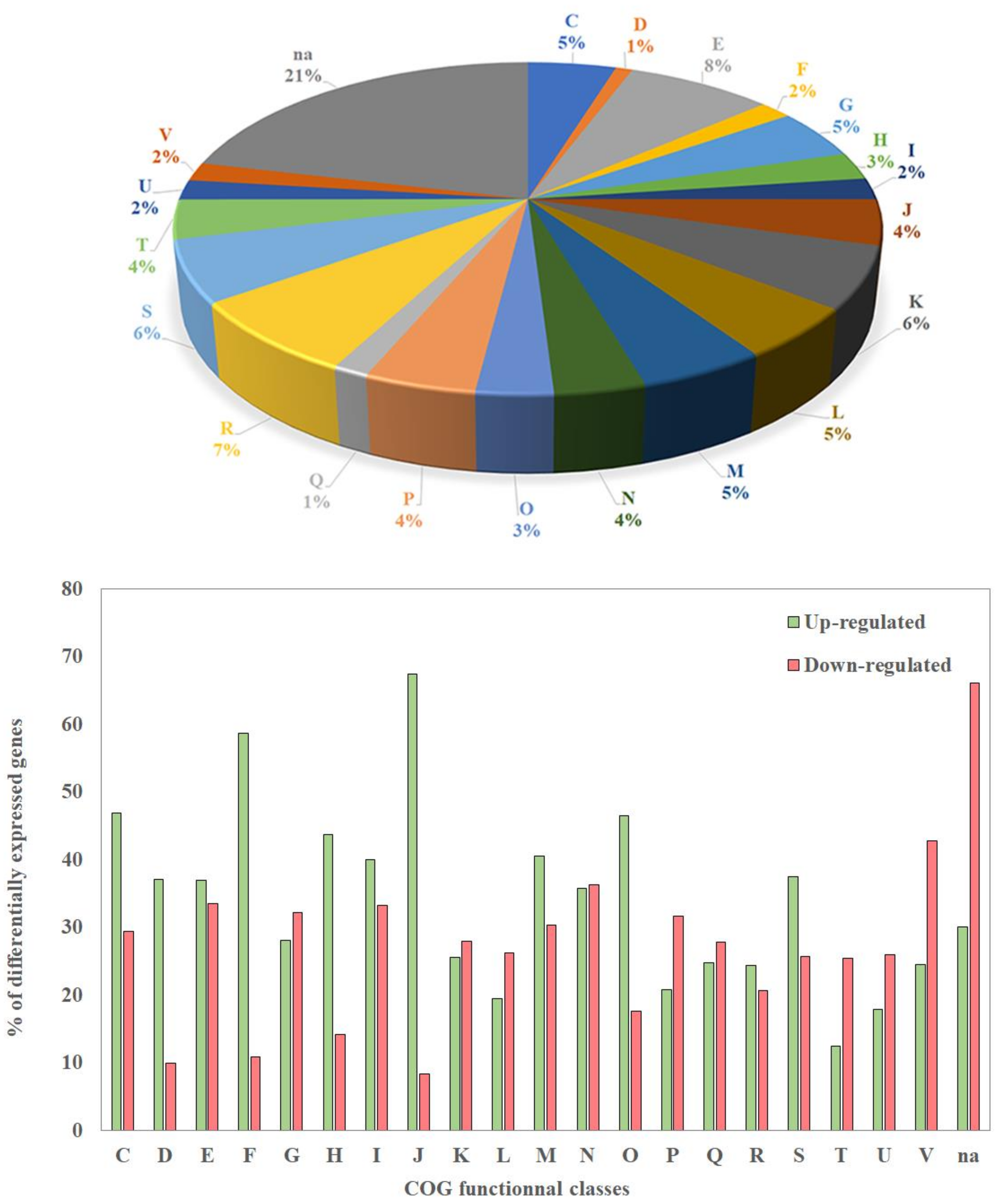
2.1. Transcriptional Activity in Biofilm Versus Planktonic Cells: Global Overview
2.2. Attachment and Motility Involving TFP
2.3. Matrix Production-Associated Genes
2.4. QS Associated Genes
2.5. Virulence-Associated Genes
3. Materials and Methods
3.1. Bacterial Strains
3.2. Planktonic Growth
3.3. Biofilm Culture
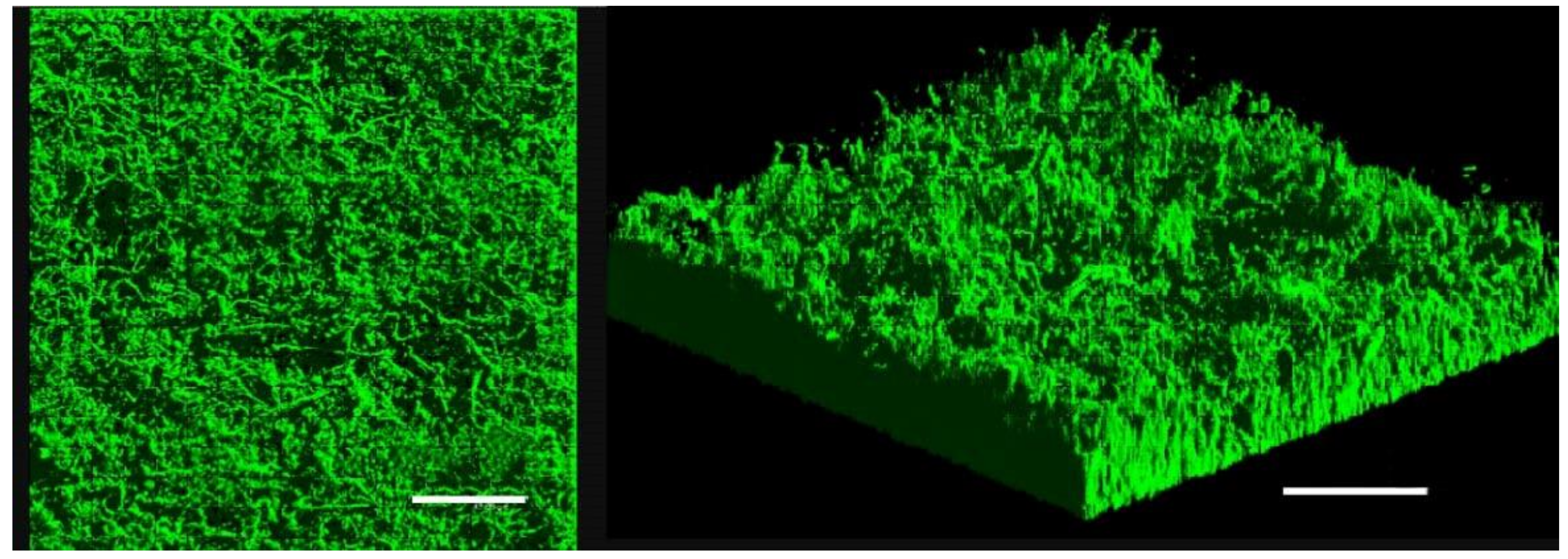
3.4. Confocal Laser Scanning Microscopy (CLSM)
3.5. RNA Extraction
3.6. RNA-Sequencing and Data Analyis
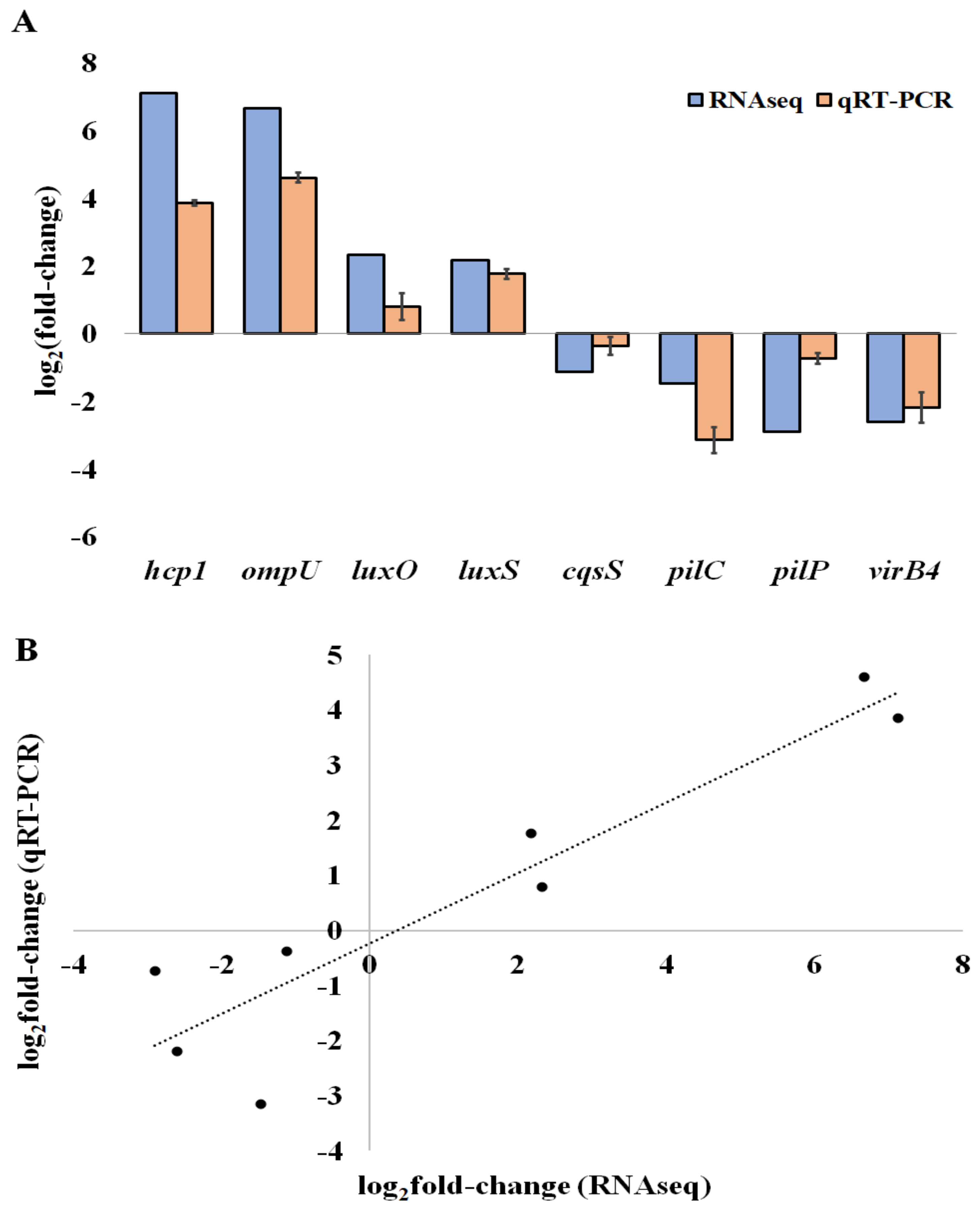
3.7. mRNA Quantification by Real-Time Reverse Transcription PCR
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Costerton, J.W.; Lewandowski, Z.; Caldwell, D.E.; Korber, D.R.; Lappin-Scott, H.M. Microbial biofilms. Annu. Rev. Microbiol. 1995, 49, 711–745. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [PubMed]
- Whiteley, M.; Bangera, M.G.; Bumgamer, R.E.; Parsek, M.R.; Teitzel, G.M.; Lory, S.; Greenberg, E.P. Gene expression in Pseudomonas aeruginosa biofilms. Nature 2001, 413, 860–864. [Google Scholar] [CrossRef] [PubMed]
- Schembri, M.A.; Kjaergaard, K.; Klemm, P. Global gene expression in Escherichia coli biofilms. Mol. Microbiol. 2003, 48, 253–267. [Google Scholar] [CrossRef] [PubMed]
- Beenken, K.E.; Dunman, P.M.; McAleese, F.; Macapagal, D.; Murphy, E.; Projan, S.J.; Blevins, J.S.; Smeltzer, M.S. Global gene expression in Staphylococcus aureus biofilms. J. Bacteriol. 2004, 186, 4665–4684. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W. Cystic fibrosis pathogenesis and the role of biofilms in persistent infection. Trends Microbiol. 2001, 9, 50–52. [Google Scholar] [CrossRef]
- Tamayo, R.; Patimalla, B.; Camilli, A. Growth in a biofilm induces a hyperinfectious phenotype in Vibrio cholerae. Infect. Immun. 2010, 78, 3560–3569. [Google Scholar] [CrossRef] [PubMed]
- Hoiby, N.; Ciofu, O.; Bjarnsholt, T. Pseudomonas aeruginosa biofilms in cystic fibrosis. Future Microbiol. 2011, 5, 1663–1674. [Google Scholar] [CrossRef] [PubMed]
- Teschler, J.K.; Zamorano-Sanchez, D.; Utada, A.S.; Warner, C.J.; Wong, G.C.; Linington, R.G.; Yildiz, F.H. Living in the matrix: Assembly and control of Vibrio cholerae biofilms. Nat. Rev. Microbiol. 2015, 13, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, S.; Paillard, C.; Le Pennec, G.; Dufour, A.; Bazire, A. Vibrio tapetis, the causative agent of Brown Ring Disease, forms biofilms with spherical components. Front. Microbiol. 2015, 6, 1384. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, F.; Fong, J.; Sadovskaya, I.; Grard, T.; Vinogradov, E. Structural characterization of the extracellular polysaccharide from Vibrio cholerae O1 El-Tor. PLoS ONE 2014, 9, e86751. [Google Scholar] [CrossRef] [PubMed]
- Guvener, Z.T.; McCarter, L.L. Multiple regulators control capsular polysaccharide production in Vibrio parahaemolyticus. J. Bacteriol. 2003, 185, 5431–5441. [Google Scholar] [CrossRef] [PubMed]
- Yip, E.S.; Grublesky, B.T.; Hussa, E.A.; Visick, K.L. A novel, conserved cluster of genes promotes symbiotic colonization and sigma-dependent biofilm formation by Vibrio fischeri. Mol. Microbiol. 2005, 57, 1485–1498. [Google Scholar] [CrossRef] [PubMed]
- Fuqua, C.; Parsek, M.R.; Greenberg, E.P. Regulation of gene expression by cell-to-cell communication: acyl-homoserine lactone quorum sensing. Annu. Rev. Genet. 2001, 35, 439–468. [Google Scholar] [CrossRef] [PubMed]
- Milton, D.L. Quorum sensing in vibrios: Complexity for diversification. Int. J. Med. Microbiol. 2006, 296, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Aljaro, C.; Melado-Rovira, S.; Milton, D.L.; Blanch, A.R. Quorum-sensing regulates biofilm formation in Vibrio scophthalmi. BMC Microbiol. 2012, 12, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, T.C.; Blackman, E.R.; Morrison, S.S.; Gibas, C.J.; Oliver, J.D. Transcriptome sequencing reveals the virulence and environmental genetic programs of Vibrio vulnificus exposed to host and estuarine conditions. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Mandlik, A.; Livny, J.; Robins, W.P.; Ritchie, J.M.; Mekalanos, J.J; Waldor, M.K. RNA-Seq-based monitoring of infection-linked changes in Vibrio cholerae gene expression. Cell Host Microbe 2011, 10, 165–174. [Google Scholar] [CrossRef] [Green Version]
- Higgins, D.A.; Pomianek, M.E.; Kraml, C.M.; Taylor, R.K.; Semmelhack, M.F.; Bassler, B.L. The major Vibrio cholerae autoinducer and its role in virulence factor production. Nature 2007, 450, 883–886. [Google Scholar] [CrossRef] [PubMed]
- Dias, G.M.; Bidault, A.; Le Chevalier, P.; Choquet, G.; Der Sarkissian, C.; Orlando, L.; Medigue, C.; Barbe, V.; Mangenot, S.; Thompson, C.C.; et al. Vibrio tapetis Displays an Original Type IV Secretion System in Strains Pathogenic for Bivalve Molluscs. Front. Microbiol. 2018, 9, 227. [Google Scholar] [CrossRef] [PubMed]
- Paillard, C. A short review of brown ring disease, a vibriosis affecting clams, Ruditapes phillipinarum and Ruditapes decussatus. Aquat. Living Resour. 2004, 17, 467–475. [Google Scholar] [CrossRef]
- Paillard, C.; Maes, P. The brown ring disease in the manila clam, Ruditapes philippinarum. II Microscopic study of the brown ring disease. J. Invertebr. Pathol. 1995, 65, 101–110. [Google Scholar] [CrossRef]
- Lakhal, F.; Bury-Mone, S.; Nomane, Y.; Le Goic, N.; Paillard, C.; Jacq, A. DjlA, a membrane-anchored DnaJ-like protein, is required for cytotoxicity of clam pathogen Vibrio tapetis to hemocytes. Appl. Environ. Microbiol. 2008, 74, 5750–5758. [Google Scholar] [CrossRef] [PubMed]
- Sternberg, C.; Bjarnsholt, T.; Shirtliff, M. Methods for dynamic investigations of surface-attached in vitro bacterial and fungal biofilms. Methods Mol. Biol. 2014, 1147, 3–22. [Google Scholar] [PubMed]
- Franca, A.; Melo, L.D.; Cerca, N. Comparison of RNA extraction methods from biofilm samples of Staphylococcus epidermidis. BMC Res. Notes 2011, 4, 572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dotsch, A.; Eckweiler, D.; Schniederjans, M.; Zimmermann, A.; Jensen, V.; Scharfe, M.; Geffers, R.; Haussler, S. The Pseudomonas aeruginosa transcriptome in planktonic cultures and static biofilms using RNA sequencing. PLoS ONE 2012, 7, e31092. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.; Franca, A.; Bradwell, K.R.; Serrano, M.G.; Jefferson, K.K.; Cerca, N. Comparative transcriptomic analysis of Gardnerella vaginalis biofilms vs. planktonic cultures using RNA-seq. NPJ Biofilms Microbiomes 2017, 3, 3. [Google Scholar] [CrossRef] [PubMed]
- Philips, J.; Rabaey, K.; Lovley, D.R.; Vargas, M. Biofilm Formation by Clostridium ljungdahlii Is Induced by Sodium Chloride Stress: Experimental Evaluation and Transcriptome Analysis. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Cury, J.A.; Koo, H. Extraction and purification of total RNA from Streptococcus mutans biofilms. Anal. Biochem. 2007, 365, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Rumbo-Feal, S.; Gomez, M.J.; Gayoso, C.; Alvarez-Fraga, L.; Cabral, M.P.; Aransay, A.M.; Rodriguez-Ezpeleta, N.; Fullaondo, A.; Valle, J.; Tomas, M.; et al. Whole transcriptome analysis of Acinetobacter baumannii assessed by RNA-sequencing reveals different mRNA expression profiles in biofilm compared to planktonic cells. PLoS ONE 2013, 8, e72968. [Google Scholar] [CrossRef]
- Folsom, J.P.; Richards, L.; Pitts, B.; Roe, F.; Ehrlich, G.D.; Parker, A.; Mazurie, A.; Stewart, P.S. Physiology of Pseudomonas aeruginosa in biofilms as revealed by transcriptome analysis. BMC Microbiol. 2010, 10, 294. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, Y.D.; Deslandes, V.; Jacques, M. Actinobacillus pleuropneumoniae genes expression in biofilms cultured under static conditions and in a drip-flow apparatus. BMC Genom. 2013, 14, 364. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2016, 11. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. B 1995, 57, 289–300. [Google Scholar]
- Shime-Hattori, A.; Iida, T.; Arita, M.; Park, K.S.; Kodama, T.; Honda, T. Two type IV pili of Vibrio parahaemolyticus play different roles in biofilm formation. FEMS Microbiol. Lett. 2006, 264, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Utada, A.S.; Bennett, R.R.; Fong, J.C.N.; Gibiansky, M.L.; Yildiz, F.H.; Golestanian, R.; Wong, G.C.L. Vibrio cholerae use pili and flagella synergistically to effect motility switching and conditional surface attachment. Nat Commun. 2014, 5, 4913. [Google Scholar] [CrossRef] [PubMed]
- Morris, A.R; Visick, K.L. Inhibition of SypG-induced biofilms and host colonization by the negative regulator SypE in Vibrio fischeri. PLoS ONE 2012, 8. [Google Scholar] [CrossRef] [PubMed]
- Ng, W.L.; Bassler, B.L. Bacterial quorum-sensing network architectures. Annu. Rev. Genet. 2009, 43, 197–222. [Google Scholar] [CrossRef] [PubMed]
- Neiditch, M.B.; Federle, M.J.; Miller, S.T.; Bassler, B.L.; Hughson, F.M. Regulation of LuxPQ receptor activity by the quorum-sensing signal autoinducer-2. Mol. Cell 2005, 18, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.J.; Benitez, J.A. Vibrio cholerae Biofilms and Cholera Pathogenesis. PLoS Negl. Trop. Dis. 2016, 10. [Google Scholar] [CrossRef] [PubMed]
- Goo, S.Y.; Lee, H.J.; Kim, W.H.; Han, K.L.; Park, D.K.; Kim, S.M.; Kim, K.S.; Lee, K.H.; Park, S.J. Identification of OmpU of Vibrio vulnificus as a fibronectin-binding protein and its role in bacterial pathogenesis. Infect. Immun. 2006, 74, 5586–5594. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Gao, H.; Xiao, N.; Liu, Y.; Li, J.; Li, L. Outer membrane protein U (OmpU) mediates adhesion of Vibrio mimicus to host cells via two novel N-terminal motifs. PLoS ONE 2015, 10, e0119026. [Google Scholar] [CrossRef] [PubMed]
- Madec, S.; Pichereau, V.; Jacq, A.; Boisset, C.; Guerard, F.; Paillard, C.; Nicolas, J.-L. Characterization of the secretomes of two vibrios pathogenic to mollusks. Plos ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, S.; Cheng, H.; Pang, H.; Lu, Y.; Jian, J. Role of the toxR Gene from Fish Pathogen Vibiro alginolyticus in the Physiology and Virulence. Indian J. Microbiol. 2017, 57, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Y.; Lauritz, J.; Jass, J.; Milton, D.L. Role for the major outer-membrane protein from Vibrio anguillarum in bile resistance and biofilm formation. Microbiology 2003, 149, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Filloux, A.; Hachani, A.; Bleves, S. The bacterial type VI secretion machine: Yet another player for protein transport across membranes. Microbiology 2008, 154, 1570–1583. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Chaudhuri, K. Identification of a unique IAHP (IcmF associated homologous proteins) cluster in Vibrio cholerae and other proteobacteria through in silico analysis. In Silico Biol. 2003, 3, 287–300. [Google Scholar] [PubMed]
- Pukatzki, S.; Ma, A.T.; Sturtevant, D.; Krastins, B.; Sarracino, D.; Nelson, W.C.; Heidelberg, J.F; Mekalanos, J.J. Identification of a conserved bacterial protein secretion system in Vibrio cholerae using the Dictyostelium host model system. Proc. Natl. Acad. Sci. USA 2006, 103, 1528–1533. [Google Scholar] [CrossRef] [PubMed]
- Linares, D.; Jean, N.; Van Overtvelt, P.; Ouidir, T.; Hardouin, J.; Blache, Y.; Molmeret, M. The marine bacteria Shewanella frigidimarina NCIMB400 upregulates the type VI secretion system during early biofilm formation. Environ. Microbiol. Rep. 2016, 8, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Tolker-Nielsen, T.; Sternberg, C. Growing and analyzing biofilms in flow chambers. Curr. Protoc. Microbiol. 2001. [Google Scholar] [CrossRef] [PubMed]
- Heydorn, A.; Nielsen, A.T.; Hentzer, M.; Sternberg, C.; Givskov, M.; Ersboll, B.K.; Molin, S. Quantification of biofilm structures by the novel computer program COMSTAT. Microbiology 2000, 146, 2395–2407. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. Fast QC, A Quality Control tool for High Throughput Sequence Data. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 20 September 2018).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq--a Python framework to work with high-throughput sequencing data. Bioinformatics 2014, 31, 166–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varet, H.; Brillet-Gueguen, L.; Coppee, J.Y.; Dillies, M.A. SARTools: A DESeq2- and EdgeR-Based R Pipeline for Comprehensive Differential Analysis of RNA-Seq Data. PLoS ONE 2016, 11, e0157022. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic. Acids. Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazire, A.; Dufour, A. The Pseudomonas aeruginosa rhlG and rhlAB genes are inversely regulated and RhlG is not required for rhamnolipid synthesis. BMC Microbiol. 2014, 14, 160. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | FC | Log2FC | Product |
|---|---|---|---|---|
| a2706 | mshA | 1.9 | 1 | Mannose-sensitive haemagglutinin A |
| a2707 | 2.1 | 1.1 | MSHA pilin protein MshA | |
| a2708 | mshA | 1.9 | 0.9 | Mannose-sensitive haemagglutinin A |
| a0251 | pilC | 0.3 | −1.5 | Type IV pilin assembly protein PilC |
| a2965 | pilQ | 0.2 | −2.2 | Fimbrial assembly protein PilQ (fragment) |
| a2966 | pilP | 0.1 | −2.9 | Putative pilus assembly protein PilP |
| a2967 | pilO | 0.3 | −1.6 | Putative pilus assembly protein PilO |
| a2968 | pilN | 0.2 | −2.3 | Putative TFP pilus assembly protein PilN |
| a2969 | pilM | 0.4 | −1.4 | Putative Type IV assembly protein PilM |
| Gene ID | KO | Gene Name | FC | Log2FC | Product |
|---|---|---|---|---|---|
| b1181 | K03606 | wcaJ | 0.1 | −2.8 | Putative UDP-sugar lipid carrier transferase |
| b1180 | K20920 | 0.3 | −1.7 | Polysaccharides biosynthesis/export protein VpsM | |
| b1179 | K20988 | 0.4 | −1.2 | Polysaccharides biosynthesis/export protein VpsN | |
| b1178 | K16554 | 0.2 | −1.9 | Polysaccharide biosynthesis transport protein | |
| b1177 | na | 0.2 | −1.9 | Putative polysaccharide biosynthesis protein | |
| a3110 | K02461 | epsL | 1.9 | 0.9 | Type II Secretion System protein L |
| a3109 | K02462 | epsM | 1.5 | 0.5 | Type II Secretion System protein M |
| a3108 | K02463 | epsN | 1.4 | 0.4 | Type II Secretion System protein N |
| a3119 | K02452 | epsC | 1.6 | 0.7 | Type II Secretion System protein C |
| a3118 | K02453 | gspD | 2.1 | 1 | General secretion pathway protein D |
| a3117 | K02454 | gspE | 2.2 | 1.2 | General secretion pathway protein E |
| a3116 | K02455 | gspF | 2 | 1 | General secretion pathway protein F |
| a3115 | K02456 | gspG | 2.7 | 1.4 | General secretion pathway protein G |
| a2724 | K00963 | galU | 2.7 | 1.4 | UTP-glucose-1-phosphate uridylyltransferase |
| a2985 | na | galE | 1.6 | 0.7 | UDP-glucose 4-epimerase |
| Gene ID | Gene Name | FC | Log2FC | Product |
|---|---|---|---|---|
| b1775 | cqsS | 0.4 | −1.1 | CAI-1 autoinducer sensor kinase/phosphatase CqsS |
| b1774 | cqsA | Not DEG | CAI-1 autoinducer synthase | |
| a0117 | luxS | 4.5 | 2.2 | S-ribosylhomocysteinase |
| b0125 | luxQ | 0.7 | −0.5 | Autoinducer 2 sensor kinase/phosphatase LuxQ |
| b0126 | luxP | Not DEG | Autoinducer 2-binding periplasmic protein LuxP | |
| a1225 | luxO | 4.9 | 2.3 | Regulatory protein LuxO |
| Gene ID | Gene Name | FC | Log2FC | Product |
|---|---|---|---|---|
| b0573 | 138.9 | 7.1 | T6SS component Hcp | |
| a2618 | yfiD | 116.3 | 6.8 | Pyruvate formate lyase subunit |
| a1598 | ompU | 101.1 | 6.6 | Outer membrane protein U |
| b0572 | 90 | 6.5 | T6SS component TssB (ImpB/VipA) | |
| b0571 | 62.6 | 5.9 | T6SS component TssC (ImpC/VipB) | |
| a3240 | 55.9 | 5.8 | Protein of unknown function |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, S.; Paillard, C.; Van Dillen, S.; Tahrioui, A.; Berjeaud, J.-M.; Dufour, A.; Bazire, A. Relation between Biofilm and Virulence in Vibrio tapetis: A Transcriptomic Study. Pathogens 2018, 7, 92. https://doi.org/10.3390/pathogens7040092
Rodrigues S, Paillard C, Van Dillen S, Tahrioui A, Berjeaud J-M, Dufour A, Bazire A. Relation between Biofilm and Virulence in Vibrio tapetis: A Transcriptomic Study. Pathogens. 2018; 7(4):92. https://doi.org/10.3390/pathogens7040092
Chicago/Turabian StyleRodrigues, Sophie, Christine Paillard, Sabine Van Dillen, Ali Tahrioui, Jean-Marc Berjeaud, Alain Dufour, and Alexis Bazire. 2018. "Relation between Biofilm and Virulence in Vibrio tapetis: A Transcriptomic Study" Pathogens 7, no. 4: 92. https://doi.org/10.3390/pathogens7040092
APA StyleRodrigues, S., Paillard, C., Van Dillen, S., Tahrioui, A., Berjeaud, J. -M., Dufour, A., & Bazire, A. (2018). Relation between Biofilm and Virulence in Vibrio tapetis: A Transcriptomic Study. Pathogens, 7(4), 92. https://doi.org/10.3390/pathogens7040092