Comparative Evaluation of the Immune Responses in Cattle Mammary Tissues Naturally Infected with Bovine Parainfluenza Virus Type 3 and Bovine Alphaherpesvirus-1

Abstract
:1. Introduction
2. Results
2.1. Detection of Beta-Actin, BPIV-3, and BoHV-1 in Mammary Tissues
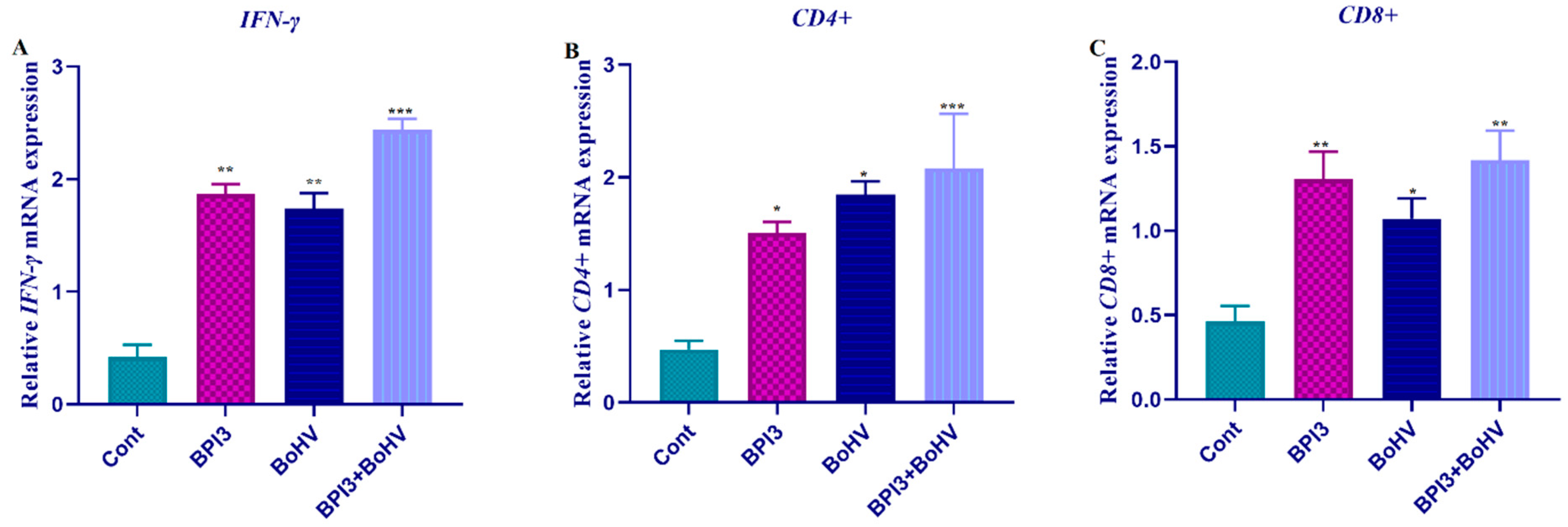
2.2. The Expression Profiles of IFN-γ, CD4, and CD8
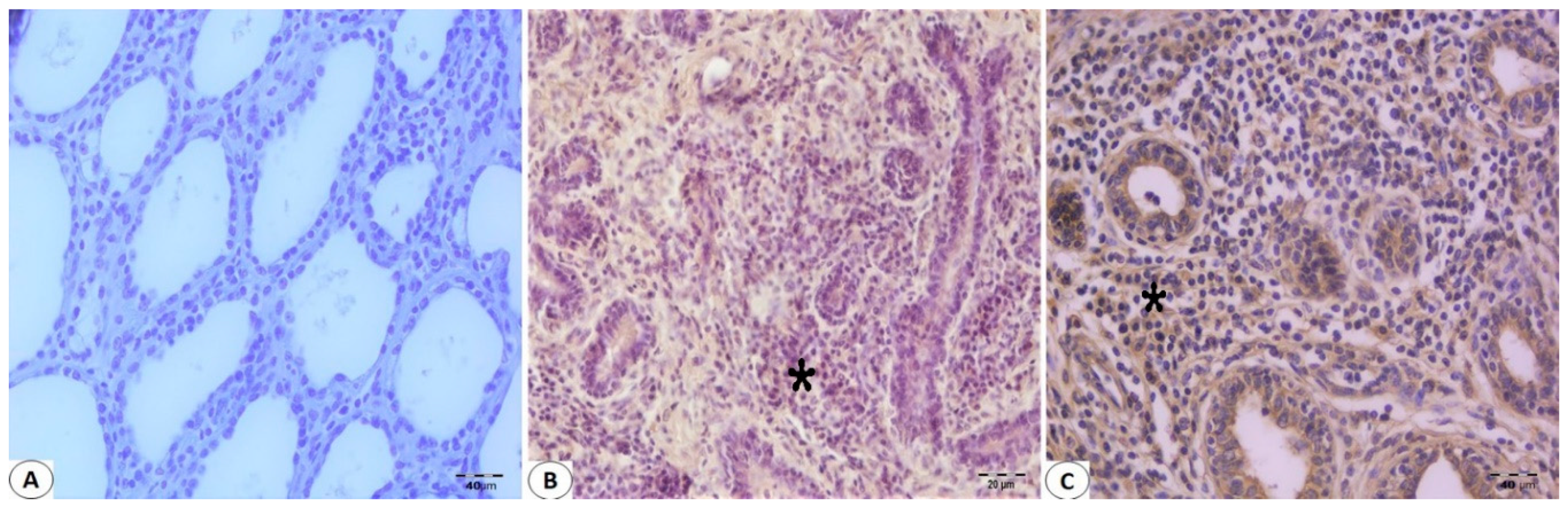
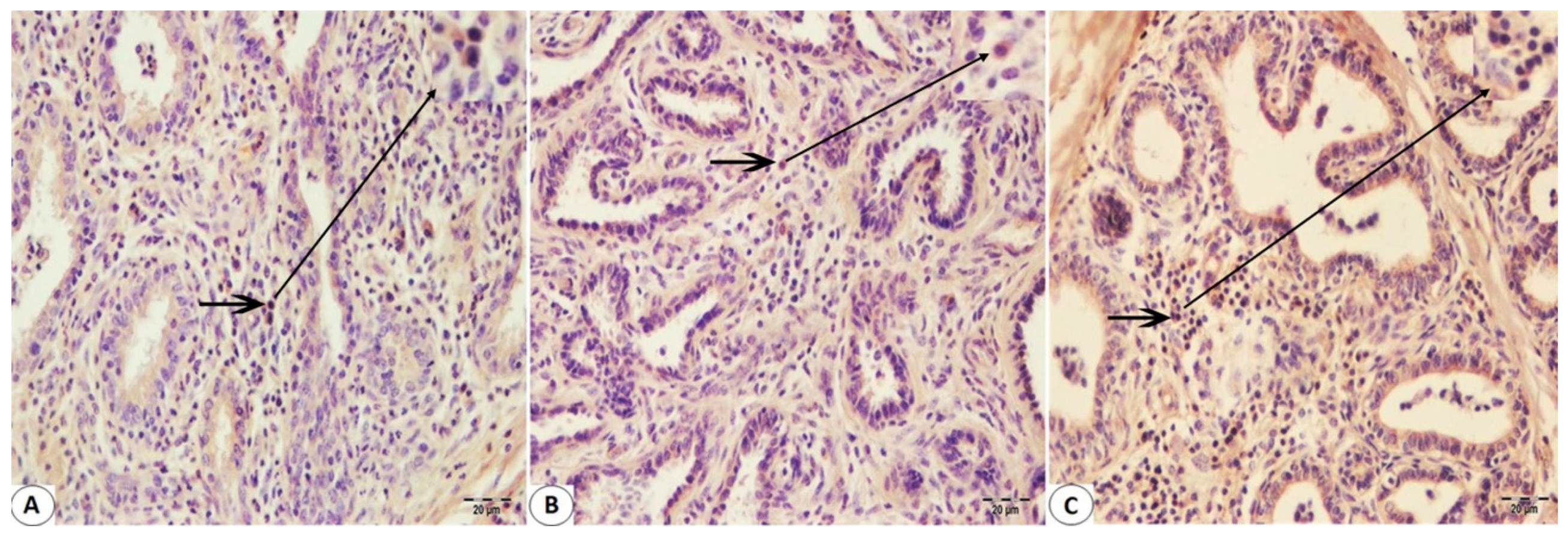
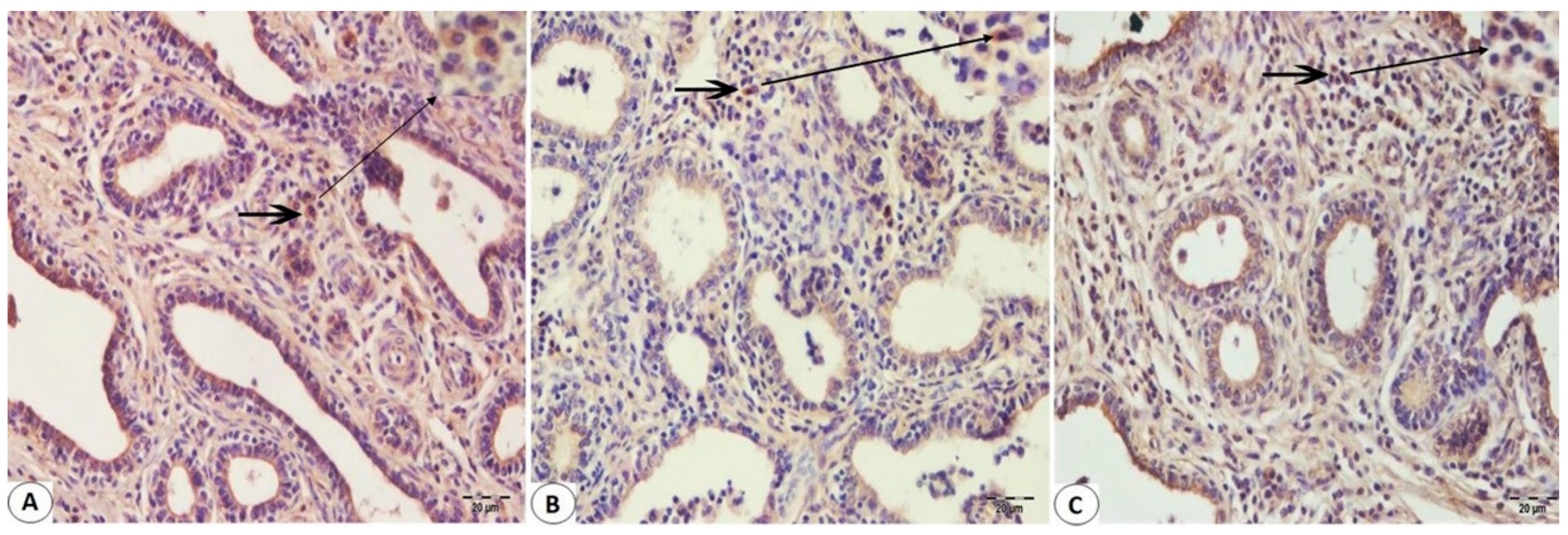
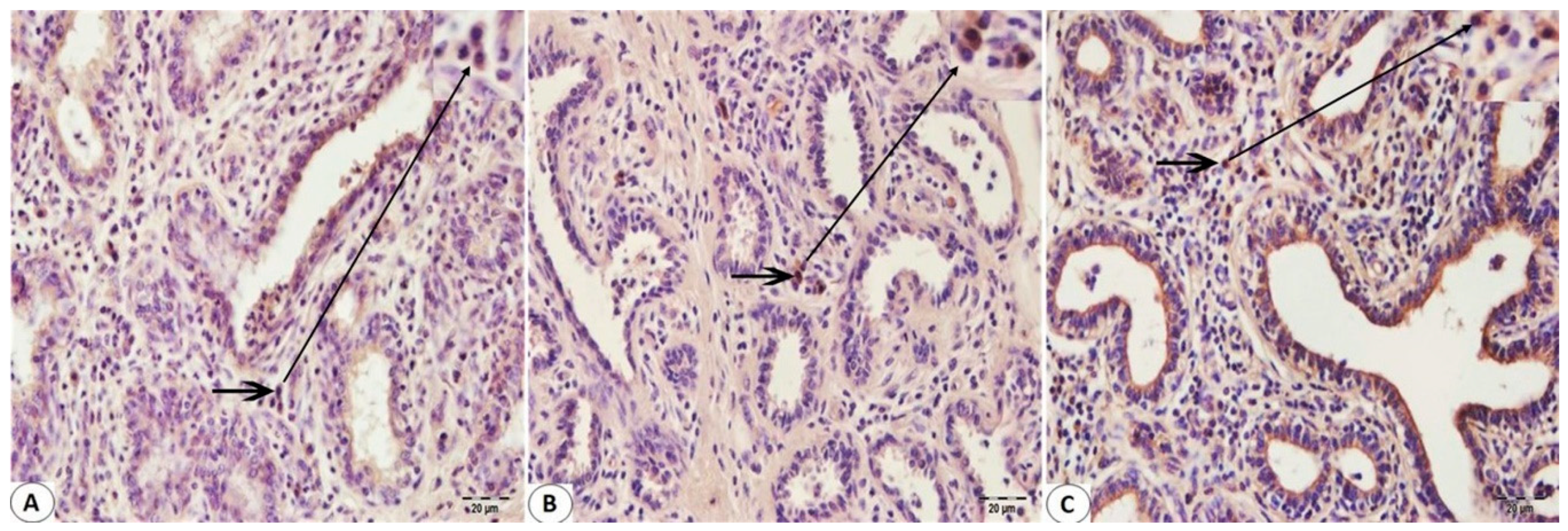
2.3. Immunohistochemistry Staining
2.4. Comparative Results
3. Discussion
4. Material and Method
4.1. Material
4.2. Total RNA Isolation and cDNA Sythesis
4.3. Real Time PCR
4.4. Immunohistochemistry Staining
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- International Dairy Federation. Bovine Mastitis. Definitions and Guidelines for Diagnosis; International Dairy Federation: Brussels, Belgium, 1987; Volume 211, pp. 3–8. [Google Scholar]
- Vaarst, M.; Enevoldsen, C. Patterns of clinical mastitis manifestations in Danish organic dairy herds. J. Dairy Res. 1997, 64, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Gourlay, R.E.J.; Espinasse, J.; Barle, C. Isolation of Mycoplasma agalactiae var bovis and infectious Bovine rhinotracheitis virus from an outbreak of mastitis in France. Vet. Rec. 1974, 95, 534–535. [Google Scholar] [CrossRef]
- Wellenberg, G.J.; van der Poel, W.H.; van der Vorst, T.J.; van Valkengoed, P.H.; Schukken, Y.H.; Wagenaar, F.; Van Oirschot, J.T. Bovine herpesvirus 4 in Bovine clinical mastitis. Vet. Rec. 2000, 147, 222–225. [Google Scholar] [CrossRef] [PubMed]
- Burrows, R.; Mann, J.A.; Greig, A.; Chapman, W.G.; Goodridge, D. The growth and persistence of foot-and-mouth disease virus in the Bovine mammary gland. J. Hyg. 1971, 69, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, Y.; Kaji, T.; Kume, T.; Omuro, M.; Hiramune, T.; Murase, N.; Matumoto, M. Infection of cattle with parainfluenza 3 virus with special reference to udder infection. I. Virus isolation from milk. Jpn. J. Microbiol. 1966, 10, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Wellenberg, G.J.; van der Poel, W.H.; Van Oirschot, J.T. Viral infections and Bovine mastitis: A review. Vet. Microbiol. 2002, 88, 27–45. [Google Scholar] [CrossRef]
- Hutchings, D.; Babiuk, L. Lymphocyte proliferative responses to separated Bovine herpesvirus 1 proteins in immune cattle. J. Virol. 1990, 64, 5114–5122. [Google Scholar] [PubMed]
- Choi, S.-H.; Splitter, G.A. Induction of MHC-unrestricted cytolytic CD4+ T cells against virally infected target cells by cross-linking CD4 molecules. J. Immunol. 1994, 153, 3874–3881. [Google Scholar] [PubMed]
- Denis, M.; Slaoui, M.; Keil, G.; Babiuk, L.; Ernst, E.; Pastoret, P.-P.; Thiry, E. Identification of different target glycoproteins for Bovine herpes virus type 1-specific cytotoxic T lymphocytes depending on the method of in vitro stimulation. Immunology 1993, 78, 7. [Google Scholar] [PubMed]
- Turin, L.; Russo, S.; Poli, G. BHV-1: New molecular approaches to control a common and widespread infection. Mol. Med. 1999, 5, 261. [Google Scholar] [CrossRef] [PubMed]
- Banos, G.; Wall, E.; Coffey, M.P.; Bagnall, A.; Gillespie, S.; Russell, G.C.; McNeilly, T.N. Identification of immune traits correlated with dairy cow health, reproduction and productivity. PLoS ONE 2013, 8, e65766. [Google Scholar] [CrossRef] [PubMed]
- Sordillo, L.; Shafer-Weaver, K.; DeRosa, D. Immunobiology of the mammary gland. J. Dairy Sci. 1997, 80, 1851–1865. [Google Scholar] [CrossRef]
- Park, Y.H.; Park, J.Y.; Kim, S.H.; Ahn, J.S.; Davies, C.J. Characterization of lymphocyte subpopulations and major histocompatibility complex haplotypes of mastitis-resistant and susceptible cows. J. Vet. Sci. 2004, 5, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Oviedo-Boyso, J.; Valdez-Alarcón, J.J.; Cajero-Juárez, M.; Ochoa-Zarzosa, A.; López-Meza, J.E.; Bravo-Patino, A.; Baizabal-Aguirre, V.M. Innate immune response of Bovine mammary gland to pathogenic bacteria responsible for mastitis. J. Infect. 2007, 54, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Bach, E.A.; Aguet, M.; Schreiber, R.D. The IFNγ receptor: A paradigm for cytokine receptor signaling. Annu. Rev. Immunol. 1997, 15, 563–591. [Google Scholar] [CrossRef] [PubMed]
- Jonasch, E.; Haluska, F.G. Interferon in oncological practice: Review of interferon biology, clinical applications, and toxicities. Oncologist 2001, 6, 34–55. [Google Scholar] [CrossRef] [PubMed]
- Young, H.A. Regulation of interferon-γ gene expression. J. Interf. Cytokine res. 1996, 16, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Carnaud, C.; Lee, D.; Donnars, O.; Park, S.-H.; Beavis, A.; Koezuka, Y.; Bendelac, A. Cutting edge: Cross-talk between cells of the innate immune system: NKT cells rapidly activate NK cells. J. Immunol. 1999, 163, 4647–4650. [Google Scholar] [PubMed]
- Frucht, D.M.; Fukao, T.; Bogdan, C.; Schindler, H.; O’Shea, J.J.; Koyasu, S. IFN-γ production by antigen-presenting cells: Mechanisms emerge. Trends Immunol. 2001, 22, 556–560. [Google Scholar] [CrossRef]
- Flaishon, L.; Hershkoviz, R.; Lantner, F.; Lider, O.; Alon, R.; Levo, Y.; Flavell, R.A.; Shachar, I. Autocrine secretion of interferon γ negatively regulates homing of immature B cells. J. Exp. Med. 2000, 192, 1381–1388. [Google Scholar] [CrossRef] [PubMed]
- Harris, D.P.; Haynes, L.; Sayles, P.C.; Duso, D.K.; Eaton, S.M.; Lepak, N.M.; Johnson, L.L.; Swain, S.L.; Lund, F.E. Reciprocal regulation of polarized cytokine production by effector B and T cells. Nat. Immunol. 2000, 1, 475. [Google Scholar] [CrossRef] [PubMed]
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-γ: An overview of signals, mechanisms and functions. J. Leukoc. Biol. 2004, 75, 163–189. [Google Scholar] [CrossRef] [PubMed]
- Shtrichman, R.; Samuel, C.E. The role of gamma interferon in antimicrobial immunity. Curr. Opin. Microbiol. 2001, 4, 251–259. [Google Scholar] [CrossRef]
- Nonnecke, B.; Kimura, K.; Goff, J.; Kehrli, M. Effects of the Mammary Gland on Functional Capacities of Blood Mononuclear Leukocyte Populations from Periparturient Cows1. J. Dairy Sci. 2003, 86, 2359–2368. [Google Scholar] [CrossRef]
- Sordillo, L.M.; Babiuk, L.A. Controlling acute Escherichia coli mastitis during the periparturient period with recombinant Bovine interferon gamma. Vet. Microbiol. 1991, 28, 189–198. [Google Scholar] [CrossRef]
- Sordillo, L.M.; Babiuk, L.A. Modulation of Bovine mammary neutrophil function during the periparturient period following in vitro exposure to recombinant Bovine interferon gamma. Vet. Immunol. Immunop. 1991, 27, 393–402. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Petrovski, K.R.; Trajcev, M.; Buneski, G. A review of the factors affecting the costs of Bovine mastitis. J. S. Afr. Vet. Assoc. 2006, 77, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Gomes, F.; Henriques, M. Control of Bovine Mastitis: Old and Recent Therapeutic Approaches. Curr. Microbiol. 2016, 72, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Koszinowski, U.H.; Reddehase, M.J.; Jonjic, S. The role of CD4 and CD8 T cells in viral infections. Curr. Opin. Immunol. 1991, 3, 471–475. [Google Scholar] [CrossRef]
- Fensterl, V.; Chattopadhyay, S.; Sen, G.C. No Love Lost Between Viruses and Interferons. Annu. Rev. Virol. 2015, 2, 549–572. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.K.; Gack, M.U. RIG-I-like receptor regulation in virus infection and immunity. Curr. Opin. Virol. 2015, 12, 7–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrar, M.A.; Schreiber, R.D. The molecular cell biology of interferon-gamma and its receptor. Annu. Rev. Immunol. 1993, 11, 571–611. [Google Scholar] [CrossRef] [PubMed]
- Ngai, P.; McCormick, S.; Small, C.; Zhang, X.; Zganiacz, A.; Aoki, N.; Xing, Z. Gamma interferon responses of CD4 and CD8 T-cell subsets are quantitatively different and independent of each other during pulmonary Mycobacterium bovis BCG infection. Infect. Immun. 2007, 75, 2244–2252. [Google Scholar] [CrossRef] [PubMed]
- Ayari-Fakhfakh, E.; Ghram, A.; Albina, E.; Cetre-Sossah, C. Expression of cytokines following vaccination of goats with a recombinant capripoxvirus vaccine expressing Rift Valley fever virus proteins. Vet. Immunol. Immunopathol. 2018, 197, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Reed, J.R.; Schwertfeger, K.L. Immune cell location and function during post-natal mammary gland development. J. Mammary Gland Biol. Neoplasia 2010, 15, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, T.; Bi, Z.; Reiss, C.S. Interferon-gamma induced type I nitric oxide synthase activity inhibits viral replication in neurons. J. Neuroimmunol. 1996, 68, 101–108. [Google Scholar] [CrossRef]
- Mikloska, Z.; Cunningham, A.L. Alpha and gamma interferons inhibit herpes simplex virus type 1 infection and spread in epidermal cells after axonal transmission. J. Virol. 2001, 75, 11821–11826. [Google Scholar] [CrossRef] [PubMed]
- Cantin, E.; Tanamachi, B.; Openshaw, H. Role for gamma interferon in control of herpes simplex virus type 1 reactivation. J. Virol. 1999, 73, 3418–3423. [Google Scholar] [PubMed]
- Geiger, K.D.; Gurushanthaiah, D.; Howes, E.L.; Lewandowski, G.A.; Reed, J.C.; Bloom, F.E.; Sarvetnick, N.E. Cytokine-mediated survival from lethal herpes simplex virus infection: Role of programmed neuronal death. Proc. Natl. Acad. Sci. USA 1995, 92, 3411–3415. [Google Scholar] [CrossRef] [PubMed]
- Sarawar, S.R.; Carding, S.R.; Allan, W.; McMickle, A.; Fujihashi, K.; Kiyono, H.; McGhee, J.R.; Doherty, P.C. Cytokine profiles of bronchoalveolar lavage cells from mice with influenza pneumonia: Consequences of CD4+ and CD8+ T cell depletion. Reg. Immunol. 1993, 5, 142–150. [Google Scholar] [PubMed]
- Graziosi, C.; Pantaleo, G.; Gantt, K.R.; Fortin, J.P.; Demarest, J.F.; Cohen, O.J.; Sekaly, R.P.; Fauci, A.S. Lack of evidence for the dichotomy of TH1 and TH2 predominance in HIV-infected individuals. Science 1994, 265, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Soghoian, D.Z.; Streeck, H. Cytolytic CD4(+) T cells in viral immunity. Expert Rev. Vaccines 2010, 9, 1453–1463. [Google Scholar] [CrossRef] [PubMed]
- Porichis, F.; Kaufmann, D.E. HIV-specific CD4 T cells and immune control of viral replication. Curr. Opin. HIV AIDS 2011, 6, 174–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theze, J.; Chakrabarti, L.A.; Vingert, B.; Porichis, F.; Kaufmann, D.E. HIV controllers: A multifactorial phenotype of spontaneous viral suppression. Clin. Immunol. 2011, 141, 15–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, D.M.; Lee, S.; Garcia-Hernandez Mde, L.; Swain, S.L. Multifunctional CD4 cells expressing gamma interferon and perforin mediate protection against lethal influenza virus infection. J. Virol. 2012, 86, 6792–6803. [Google Scholar] [CrossRef] [PubMed]
- Ranasinghe, S.; Flanders, M.; Cutler, S.; Soghoian, D.Z.; Ghebremichael, M.; Davis, I.; Lindqvist, M.; Pereyra, F.; Walker, B.D.; Heckerman, D.; et al. HIV-specific CD4 T cell responses to different viral proteins have discordant associations with viral load and clinical outcome. J. Virol. 2012, 86, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Soghoian, D.Z.; Jessen, H.; Flanders, M.; Sierra-Davidson, K.; Cutler, S.; Pertel, T.; Ranasinghe, S.; Lindqvist, M.; Davis, I.; Lane, K.; et al. HIV-specific cytolytic CD4 T cell responses during acute HIV infection predict disease outcome. Sci. Transl. Med. 2012, 4, 123–125. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, T.M.; Li, C.K.; Chui, C.S.; Huang, A.K.; Perkins, M.; Liebner, J.C.; Lambkin-Williams, R.; Gilbert, A.; Oxford, J.; Nicholas, B.; et al. Preexisting influenza-specific CD4+ T cells correlate with disease protection against influenza challenge in humans. Nat. Med. 2012, 18, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Callendret, B.; Xu, D.; Brasky, K.M.; Feng, Z.; Hensley, L.L.; Guedj, J.; Perelson, A.S.; Lemon, S.M.; Lanford, R.E.; et al. Dominance of the CD4(+) T helper cell response during acute resolving hepatitis A virus infection. J. Exp. Med. 2012, 209, 1481–1492. [Google Scholar] [CrossRef] [PubMed]
- Swain, S.L.; McKinstry, K.K.; Strutt, T.M. Expanding roles for CD4(+) T cells in immunity to viruses. Nat. Rev. Immunol. 2012, 12, 136–148. [Google Scholar] [CrossRef] [PubMed]
- Babiuk, L.A.; van Drunen Littel-van den Hurk, S.; Tikoo, S.K. Immunology of Bovine herpesvirus 1 infection. Vet. Microbiol. 1996, 53, 31–42. [Google Scholar] [CrossRef]
- Biswas, S.; Bandyopadhyay, S.; Dimri, U.; Patra, P.H. Bovine herpesvirus-1 (BHV-1)—A re-emerging concern in livestock: A revisit to its biology, epidemiology, diagnosis, and prophylaxis. Vet. Q. 2013, 33, 68–81. [Google Scholar] [CrossRef] [PubMed]
- Weynants, V.; Walravens, K.; Didembourg, C.; Flanagan, P.; Godfroid, J.; Letesson, J.J. Quantitative assessment by flow cytometry of T-lymphocytes producing antigen-specific gamma-interferon in Brucella immune cattle. Vet. Immunol. Immunopathol. 1998, 66, 309–320. [Google Scholar] [CrossRef]
- Hu, Z.; Molloy, M.J.; Usherwood, E.J. CD4(+) T-cell dependence of primary CD8(+) T-cell response against vaccinia virus depends upon route of infection and viral dose. Cell. Mol. Immunol. 2016, 13, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Winkler, M.T.; Doster, A.; Jones, C. Bovine herpesvirus 1 can infect CD4(+) T lymphocytes and induce programmed cell death during acute infection of cattle. J. Virol. 1999, 73, 8657–8668. [Google Scholar] [PubMed]
- Liu, T.; Khanna, K.M.; Chen, X.; Fink, D.J.; Hendricks, R.L. CD8(+) T cells can block herpes simplex virus type 1 (HSV-1) reactivation from latency in sensory neurons. J. Exp. Med. 2000, 191, 1459–1466. [Google Scholar] [CrossRef] [PubMed]
- Divito, S.; Cherpes, T.L.; Hendricks, R.L. A triple entente: Virus, neurons, and CD8+ T cells maintain HSV-1 latency. Immunol. Res. 2006, 36, 119–126. [Google Scholar] [CrossRef]
- Burrows, S.R.; Moss, D.J.; Khanna, R. Understanding human T-cell-mediated immunoregulation through herpesviruses. Immunol. Cell Biol. 2011, 89, 352–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verweij, M.C.; Horst, D.; Griffin, B.D.; Luteijn, R.D.; Davison, A.J.; Ressing, M.E.; Wiertz, E.J. Viral inhibition of the transporter associated with antigen processing (TAP): A striking example of functional convergent evolution. PLoS Pathog. 2015, 11, e1004743. [Google Scholar] [CrossRef] [PubMed]
- Hart, J.; MacHugh, N.D.; Sheldrake, T.; Nielsen, M.; Morrison, W.I. Identification of immediate early gene products of Bovine herpes virus 1 (BHV-1) as dominant antigens recognized by CD8 T cells in immune cattle. J. Gen. Virol. 2017, 98, 1843–1854. [Google Scholar] [CrossRef] [PubMed]
- Andrejeva, J.; Childs, K.S.; Young, D.F.; Carlos, T.S.; Stock, N.; Goodbourn, S.; Randall, R.E. The V proteins of paramyxoviruses bind the IFN-inducible RNA helicase, mda-5, and inhibit its activation of the IFN-beta promoter. Proc. Natl. Acad. Sci. USA 2004, 101, 17264–17269. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Kitagawa, Y.; Zhou, M.; Itoh, M.; Gotoh, B. An anti-interferon activity shared by paramyxovirus C proteins: Inhibition of Toll-like receptor 7/9-dependent alpha interferon induction. FEBS Lett. 2014, 588, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Kotenko, S.V.; Gallagher, G.; Baurin, V.V.; Lewis-Antes, A.; Shen, M.; Shah, N.K.; Langer, J.A.; Sheikh, F.; Dickensheets, H.; Donnelly, R.P. IFN-lambdas mediate antiviral protection through a distinct class II cytokine receptor complex. Nat. Immunol. 2003, 4, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Eberle, K.C.; McGill, J.L.; Reinhardt, T.A.; Sacco, R.E. Parainfluenza Virus 3 Blocks Antiviral Mediators Downstream of the Interferon Lambda Receptor by Modulating Stat1 Phosphorylation. J. Virol. 2015, 90, 2948–2958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguayo-Hiraldo, P.I.; Arasaratnam, R.J.; Tzannou, I.; Kuvalekar, M.; Lulla, P.; Naik, S.; Martinez, C.A.; Piedra, P.A.; Vera, J.F.; Leen, A.M. Characterizing the Cellular Immune Response to Parainfluenza Virus 3. J. Infect. Dis. 2017, 216, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Kubis, P.; Materniak, M.; Kuzmak, J. Comparison of nested PCR and qPCR for the detection and quantitation of BoHV6 DNA. J. Virol. Methods 2013, 194, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Thonur, L.; Maley, M.; Gilray, J.; Crook, T.; Laming, E.; Turnbull, D.; Nath, M.; Willoughby, K. One-step multiplex real time RT-PCR for the detection of Bovine respiratory syncytial virus, Bovine herpesvirus 1 and Bovine parainfluenza virus 3. BMC Vet. Res. 2012, 8, 37. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material | BPIV-3 | BoHV-1 | BPIV-3 and BoHV-1 |
|---|---|---|---|
| Macroscopic lesions | Absence | Absence | Absence |
| Mammary tissue | 26 | 16 | 5 |
| Percentage (%) | 21.66% | 13.3% | 4.16% |
| N = 5 | CD4 + T cells | CD8 + T cells | IFN-γ |
|---|---|---|---|
| BPIV-3 | 0.80 ± 0.44 a | 2.20 ± 0.44 b | 1.40 ± 0.89 c |
| BoHV-1 | 1.66 ± 0.51 aı | 1.00 ± 1.09 bı | 1.66 ± 1.03 cı |
| BPIV-3 + BoHV-1 | 2.75 ± 0.50 aıı | 2.75 ± 0.50 aıı | 2.75 ± 0.50 cıı |
| Gene Name | Primer Sequence | Amplicon Size (bp) | Accession No | Reaction Conditions |
| Bo-IFN-γ | F - AATTCCGGTGGATGATCTGC R - TCTCCGGCCTCGAAAGAGAT | 130 | NM_174086.1 | 94 °C 15 s/55 °C 30 s/72 °C 30 s (40 cycles) |
| Bo-CD4 | F - TGATGAAAGTGACTAAGTCCCCA R- TTCGGCTGATTTGAGCCCTT | 121 | NM_001103225.1 | 94 °C 15 s/58 °C 30 s/72 °C 30 s (40 cycles) |
| Bo-CD8 | F - CTGGACTTCGCCTGCAATATC R- CACGTCTTCGGTTCCGGC | 112 | NM_174015.1 | 94 °C 15 s/57 °C 30 s/72 °C 30 s (40 cycles) |
| Bo-Beta Actin | F - GTCGACACCGCAACCAGTTC R - TACGAGTCCTTCTGGCCCAT | 181 | AY141970.1 | 94 °C 15 s/57 °C 30 s/72 °C 30 s (40 cycles) |
| Gene Name | Primer Sequences | Reference | Reactions Conditions | |
| BPIV-3 | F: TGATTGGATGTTCGGGAGTGA R: AGAATCCTTTCCTCAATCCTGATATACT | [28] | 94 °C for 15 s, 58 °C for 30 s/72 °C for 30 s (40 cycle) | |
| BoHV-1 | F: TGTGGACCTAAACCTCACGGT R: GTAGTCGAGCAGACCCGTGTC | [28] | 94 °C for 15 s, 59 °C for 30 s/72 °C for 30 s (40 cycle) | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
ÇOMAKLI, S.; ÖZDEMİR, S. Comparative Evaluation of the Immune Responses in Cattle Mammary Tissues Naturally Infected with Bovine Parainfluenza Virus Type 3 and Bovine Alphaherpesvirus-1. Pathogens 2019, 8, 26. https://doi.org/10.3390/pathogens8010026
ÇOMAKLI S, ÖZDEMİR S. Comparative Evaluation of the Immune Responses in Cattle Mammary Tissues Naturally Infected with Bovine Parainfluenza Virus Type 3 and Bovine Alphaherpesvirus-1. Pathogens. 2019; 8(1):26. https://doi.org/10.3390/pathogens8010026
Chicago/Turabian StyleÇOMAKLI, Selim, and Selçuk ÖZDEMİR. 2019. "Comparative Evaluation of the Immune Responses in Cattle Mammary Tissues Naturally Infected with Bovine Parainfluenza Virus Type 3 and Bovine Alphaherpesvirus-1" Pathogens 8, no. 1: 26. https://doi.org/10.3390/pathogens8010026
APA StyleÇOMAKLI, S., & ÖZDEMİR, S. (2019). Comparative Evaluation of the Immune Responses in Cattle Mammary Tissues Naturally Infected with Bovine Parainfluenza Virus Type 3 and Bovine Alphaherpesvirus-1. Pathogens, 8(1), 26. https://doi.org/10.3390/pathogens8010026