A Contemporary View of Respiratory Syncytial Virus (RSV) Biology and Strain-Specific Differences


Abstract
:1. An Overview of RSV Biology
1.1. RSV Disease and Vaccines
1.2. RSV Genome
1.3. RSV Antigenicity and Infectivity
1.4. RSV Genetic Diversity
2. Common Laboratory Strains of RSV
2.1. RSV Long
2.2. RSV A2
2.3. RSV Line 19 and A2-line19F
2.4. RSV CH-18537
2.5. RSV Memphis-37
3. Contemporary RSV Strains, Pathogenesis, and Viral Evolution
3.1. The Genetics and Evolution of RSV Clinical Isolates
3.2. Disease, Pathogenesis, and Cytopathology among Clinical Isolates
4. Summary and Future Implications for Vaccine Design
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Nair, H.; Simoes, E.A.; Rudan, I.; Gessner, B.D.; Azziz-Baumgartner, E.; Zhang, J.S.F.; Feikin, D.R.; A Mackenzie, G.; Moiïsi, J.C.; Roca, A.; et al. Global and regional burden of hospital admissions for severe acute lower respiratory infections in young children in 2010: A systematic analysis. Lancet 2013, 381, 1380–1390. [Google Scholar] [CrossRef]
- Mazur, N.; Higgins, D.; Nunes, M.C.; Melero, J.A.; Langedijk, A.C.; Horsley, N.; Buchholz, U.J.; Openshaw, P.J.; McLellan, J.S.; Englund, J.A.; et al. The respiratory syncytial virus vaccine landscape: Lessons from the graveyard and promising candidates. Lancet Infect Dis. 2018, 18, e295–e311. [Google Scholar] [CrossRef]
- Thompson, W.W.; Shay, D.K.; Weintraub, E.; Brammer, L.; Cox, N.; Anderson, L.J.; Fukuda, K. Mortality Associated with Influenza and Respiratory Syncytial Virus in the United States. JAMA 2003, 289, 179–186. [Google Scholar] [CrossRef]
- Dowell, S.F.; Anderson, L.J.; Gary, H.E.; Erdman, D.D.; Plouffe, J.F.; File, T.M.; Marston, B.J.; Breiman, R.F. Respiratory Syncytial Virus Is an Important Cause of Community-Acquired Lower Respiratory Infection among Hospitalized Adults. J. Infect. Dis. 1996, 174, 456–462. [Google Scholar] [CrossRef] [Green Version]
- Walsh, E.E.; Peterson, D.R.; Falsey, A.R. Is Clinical Recognition of Respiratory Syncytial Virus Infection in Hospitalized Elderly and High-Risk Adults Possible? J. Infect. Dis. 2007, 195, 1046–1051. [Google Scholar] [CrossRef] [PubMed]
- Falsey, A.R.; Hennessey, P.A.; Formica, M.A.; Cox, C.; Walsh, E.E. Respiratory Syncytial Virus Infection in Elderly and High-Risk Adults. N. Engl. J. Med. 2005, 352, 1749–1759. [Google Scholar] [CrossRef]
- Shi, T.; McAllister, D.A.; O’Brien, K.L.; Simões, E.A.F.; Madhi, S.A.; Gessner, B.D.; Polack, F.P.; Balsells, E.; Acacio, S.; Aguayo, C.; et al. Global, regional, and national disease burden estimates of acute lower respiratory infections due to respiratory syncytial virus in young children in 2015: A systematic review and modelling study. Lancet 2017, 390, 946–958. [Google Scholar] [CrossRef]
- Miyairi, I.; DeVincenzo, J.P. Human Genetic Factors and Respiratory Syncytial Virus Disease Severity. Clin. Microbiol. Rev. 2008, 21, 686–703. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, K.; Kurachek, S.C.; Cairns, L.M.; Burns, J.C.; Goodspeed, B. Treatment of respiratory viral infection in an immunodeficient infant with ribavirin aerosol. Am. J. Dis. Child. 1984, 138, 305–308. [Google Scholar] [CrossRef]
- Macdonald, N.E.; Hall, C.B.; Suffin, S.C.; Alexson, C.; Harris, P.J.; Manning, J.A. Respiratory Syncytial Viral Infection in Infants with Congenital Heart Disease. N. Engl. J. Med. 1982, 307, 397–400. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.K.; McMillan, J.A.; Gross, S.J. Rehospitalization for respiratory illness in infants of less than 32 weeks’ gestation. PEDIATRICS 1991, 88, 527–532. [Google Scholar] [PubMed]
- Berkovich, S. Acute respiratory illness in the premature nursery associated with respiratory syncytial virus infections. Pediatrics 1964, 34, 753–760. [Google Scholar] [PubMed]
- Collins, P.L.; Melero, J.A. Progress in understanding and controlling respiratory syncytial virus: Still crazy after all these years. Virus Res. 2011, 162, 80–99. [Google Scholar] [CrossRef] [Green Version]
- Ke, Z.; Dillard, R.S.; Chirkova, T.; Leon, F.; Stobart, C.C.; Hampton, C.M.; Strauss, J.D.; Rajan, D.; Rostad, C.A.; Taylor, J.V.; et al. The Morphology and Assembly of Respiratory Syncytial Virus Revealed by Cryo-Electron Tomography. Viruses 2018, 10, 446. [Google Scholar] [CrossRef] [PubMed]
- Hotard, A.L.; Shaikh, F.Y.; Lee, S.; Yan, D.; Teng, M.N.; Plemper, R.K.; Crowe, J.E.; Moore, M.L.; Lopez-Ona, A. A Stabilized Respiratory Syncytial Virus Reverse Genetics System Amenable to Recombination Mediated Mutagenesis. Virology 2012, 434, 129–136. [Google Scholar] [CrossRef]
- Collins, P.L.; Hill, M.G.; Camargo, E.; Grosfeld, H.; Chanock, R.M.; Murphy, B.R. Production of infectious human respiratory syncytial virus from cloned cDNA confirms an essential role for the transcription elongation factor from the 5’ proximal open reading frame of the M2 mRNA in gene expression and provides a capability for vaccine development. Proc. Natl. Acad. Sci. USA 1995, 92, 11563–11567. [Google Scholar]
- Bitko, V.; Shulyayeva, O.; Mazumder, B.; Musiyenko, A.; Ramaswamy, M.; Look, D.C.; Barik, S. Nonstructural proteins of respiratory syncytial virus suppress premature apoptosis by an NF-κB-dependent, interferon-independent mechanism and facilitate virus growth. J. Virol. 2007, 81, 1786–1795. [Google Scholar] [CrossRef]
- Swedan, S.; Musiyenko, A.; Barik, S. Respiratory Syncytial Virus Nonstructural Proteins Decrease Levels of Multiple Members of the Cellular Interferon Pathways. J. Virol. 2009, 83, 9682–9693. [Google Scholar] [CrossRef] [Green Version]
- Spann, K.M.; Tran, K.C.; Collins, P.L. Effects of Nonstructural Proteins NS1 and NS2 of Human Respiratory Syncytial Virus on Interferon Regulatory Factor 3, NF-κB, and Proinflammatory Cytokines. J. Virol. 2005, 79, 5353–5362. [Google Scholar] [CrossRef]
- Lo, M.S.; Brazas, R.M.; Holtzman, M.J. Respiratory Syncytial Virus Nonstructural Proteins NS1 and NS2 Mediate Inhibition of Stat2 Expression and α/β Interferon Responsiveness. J. Virol. 2005, 79, 9315–9319. [Google Scholar] [CrossRef] [PubMed]
- Teng, M.N.; Whitehead, S.S.; Collins, P.L. Contribution of the Respiratory Syncytial Virus G Glycoprotein and Its Secreted and Membrane-Bound Forms to Virus Replication In Vitro and In Vivo. Virology 2001, 289, 283–296. [Google Scholar] [CrossRef]
- Sastre, P.; Melero, J.A.; Garcia-Barreno, B.; Palomo, C. Comparison of affinity chromatography and adsorption to vaccinia virus recombinant infected cells for depletion of antibodies directed against respiratory syncytial virus glycoproteins present in a human immunoglobulin preparation. J. Med. Virol. 2005, 76, 248–255. [Google Scholar] [CrossRef]
- Cortjens, B.; Yasuda, E.; Yu, X.; Wagner, K.; Claassen, Y.B.; Bakker, A.Q.; Van Woensel, J.B.M.; Beaumont, T. Broadly Reactive Anti-Respiratory Syncytial Virus G Antibodies from Exposed Individuals Effectively Inhibit Infection of Primary Airway Epithelial Cells. J. Virol. 2017, 91, e02357-16. [Google Scholar] [CrossRef] [Green Version]
- Ngwuta, J.O.; Chen, M.; Modjarrad, K.; Joyce, M.G.; Kanekiyo, M.; Kumar, A.; Yassine, H.M.; Moin, S.M.; Killikelly, A.M.; Chuang, G.-Y.; et al. Prefusion F–specific antibodies determine the magnitude of RSV neutralizing activity in human sera. Sci. Transl. Med. 2015, 7, 309ra162. [Google Scholar] [CrossRef]
- Levine, S.; Klaiber-Franco, R.; Paradiso, P.R. Demonstration that Glycoprotein G Is the Attachment Protein of Respiratory Syncytial Virus. J. Virol. 1987, 68, 2521–2524. [Google Scholar] [CrossRef] [PubMed]
- Hendricks, D.A.; Baradaran, K.; McIntosh, K.; Patterson, J.L. Appearance of a Soluble Form of the G Protein of Respiratory Syncytial Virus in Fluids of Infected Cells. J. Virol. 1987, 68, 1705–1714. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.R.; Lichtenstein, D.; Ball, L.A.; Wertz, G.W. The membrane-associated and secreted forms of the respiratory syncytial virus attachment glycoprotein G are synthesized from alternative initiation codons. J. Virol. 1994, 68, 4538–4546. [Google Scholar] [PubMed]
- Kwilas, S.; Liesman, R.M.; Zhang, L.; Walsh, E.; Pickles, R.J.; Peeples, M.E. Respiratory Syncytial Virus Grown in Vero Cells Contains a Truncated Attachment Protein That Alters Its Infectivity and Dependence on Glycosaminoglycans. J. Virol. 2009, 83, 10710–10718. [Google Scholar] [CrossRef] [Green Version]
- Melero, J.A.; Mas, V.; McLellan, J.S. Structural, antigenic and immunogenic features of respiratory syncytial virus glycoproteins relevant for vaccine development. Vaccine 2017, 35, 461–468. [Google Scholar] [CrossRef]
- Johnson, P.R.; Spriggs, M.K.; Olmsted, R.A.; Collins, P.L. The G glycoprotein of human respiratory syncytial viruses of subgroups A and B: Extensive sequence divergence between antigenically related proteins. Proc. Natl. Acad. Sci. USA 1987, 84, 5625–5629. [Google Scholar] [CrossRef]
- Collins, P.L.; Mottet, G. Oligomerization and post-translational processing of glycoprotein G of human respiratory syncytial virus: Altered O-glycosylation in the presence of brefeldin A. J. Virol. 1992, 73, 849–863. [Google Scholar] [CrossRef] [PubMed]
- Satake, M.; Coligan, J.E.; Elango, N.; Norrby, E.; Venkatesan, S. Respiratory syncytial virus envelope glycoprotein (G) has a novel structure. Nucleic Acids Res. 1985, 13, 7795–7812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLellan, J.S.; Ray, W.C.; Peeples, M.E. Structure and function of respiratory syncytial virus surface glycoproteins. Curr. Top Microbiol. Immunol. 2013, 372, 83–104. [Google Scholar] [CrossRef] [PubMed]
- Tripp, R.A.; Jones, L.P.; Haynes, L.M.; Zheng, H.; Murphy, P.M.; Anderson, L.J. CX3C chemokine mimicry by respiratory syncytial virus G glycoprotein. Nat. Immunol. 2001, 2, 732–738. [Google Scholar] [CrossRef]
- Hickling, T.P.; Malhotra, R.; Bright, H.; McDowell, W.; Blair, E.D.; Sim, R.B. Lung Surfactant Protein A Provides a Route of Entry for Respiratory Syncytial Virus into Host Cells. Viral. Immunol. 2000, 13, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Barr, F.E.; Pedigo, H.; Johnson, T.R.; Shepherd, V.L. Surfactant Protein-A Enhances Uptake of Respiratory Syncytial Virus by Monocytes and U937 Macrophages. Am. J. Respir. Cell Mol. Boil. 2000, 23, 586–592. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, R.; Ward, M.; Bright, H.; Priest, R.; Foster, M.R.; Hurle, M.; Blair, E.; Bird, M. Isolation and characterisation of potential respiratory syncytial virus receptor(s) on epithelial cells. Microbes Infect. 2003, 5, 123–133. [Google Scholar] [CrossRef]
- Feldman, S.A.; Audet, S.; Beeler, J.A. The Fusion Glycoprotein of Human Respiratory Syncytial Virus Facilitates Virus Attachment and Infectivity via an Interaction with Cellular Heparan Sulfate. J. Virol. 2000, 74, 6442–6447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chirkova, T.; Stobart, C.C.; Hartert, T.V.; Lin, S.; Gaston, K.A.; Anderson, L.J.; Oomens, A.G.P.; Boyoglu-Barnum, S.; Moore, M.L.; Ziady, A.G.; et al. CX3CR1 is an important surface molecule for respiratory syncytial virus infection in human airway epithelial cells. J. Gen. Virol. 2015, 96, 2543–2556. [Google Scholar] [CrossRef]
- Meng, J.; Hotard, A.L.; Currier, M.G.; Lee, S.; Stobart, C.C.; Moore, M.L. Respiratory Syncytial Virus Attachment Glycoprotein Contribution to Infection Depends on the Specific Fusion Protein. J. Virol. 2016, 90, 245–253. [Google Scholar] [CrossRef] [Green Version]
- McLellan, J.S.; Chen, M.; Leung, S.; Graepel, K.W.; Du, X.; Yang, Y.; Zhou, T.; Baxa, U.; Yasuda, E.; Beaumont, T.; et al. Structure of RSV Fusion Glycoprotein Trimer Bound to a Prefusion-Specific Neutralizing Antibody. Science 2013, 340, 1113–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stobart, C.C.; Rostad, C.A.; Ke, Z.; Dillard, R.S.; Hampton, C.M.; Strauss, J.D.; Yi, H.; Hotard, A.L.; Meng, J.; Pickles, R.J.; et al. A live RSV vaccine with engineered thermostability is immunogenic in cotton rats despite high attenuation. Nat. Commun. 2016, 7, 13916. [Google Scholar] [CrossRef]
- Tayyari, F.; Marchant, D.; Moraes, T.J.; Duan, W.; Mastrangelo, P.; Hegele, R.G. Identification of nucleolin as a cellular receptor for human respiratory syncytial virus. Nat. Med. 2011, 17, 1132–1135. [Google Scholar] [CrossRef] [PubMed]
- Villenave, R.; Nguyen, M.T.; Hammonds, J.; Sakamoto, K.; Lee, S.; Meng, J.; Currier, M.G.; Stobart, C.C.; Hotard, A.L.; Pretto, C.D.; et al. EGFR Interacts with the Fusion Protein of Respiratory Syncytial Virus Strain 2-20 and Mediates Infection and Mucin Expression. PLOS Pathog. 2016, 12, e1005622. [Google Scholar]
- Battles, M.B.; McLellan, J.S. Respiratory syncytial virus entry and how to block it. Nat. Rev. Microbiol. 2019, 17, 233–245. [Google Scholar] [CrossRef] [PubMed]
- McLellan, J.S.; Chen, M.; Joyce, M.G.; Sastry, M.; Stewart-Jones, G.B.E.; Yang, Y.; Zhang, B.; Chen, L.; Srivatsan, S.; Zheng, A.; et al. Structure-Based Design of a Fusion Glycoprotein Vaccine for Respiratory Syncytial Virus. Science 2013, 342, 592–598. [Google Scholar] [CrossRef] [Green Version]
- Karron, R.A.; Buchholz, U.J.; Collins, P.L. Live-Attenuated Respiratory Syncytial Virus Vaccines; Springer Nature: Berlin, Germany, 2013; Volume 372, pp. 259–284. [Google Scholar]
- Mufson, M.A.; Orvell, C.; Rafnar, B.; Norrby, E. Two Distinct Subtypes of Human Respiratory Syncytial Virus. J. Virol. 1985, 66, 2111–2124. [Google Scholar] [CrossRef]
- Zlateva, K.T.; Lemey, P.; Vandamme, A.-M.; Van Ranst, M. Molecular Evolution and Circulation Patterns of Human Respiratory Syncytial Virus Subgroup A: Positively Selected Sites in the Attachment G Glycoprotein. J. Virol. 2004, 78, 4675–4683. [Google Scholar] [CrossRef] [Green Version]
- Zlateva, K.T.; Lemey, P.; Moës, E.; Vandamme, A.-M.; Van Ranst, M. Genetic Variability and Molecular Evolution of the Human Respiratory Syncytial Virus Subgroup B Attachment G Protein. J. Virol. 2005, 79, 9157–9167. [Google Scholar] [CrossRef] [Green Version]
- Sande, C.J.; Mutunga, M.N.; Medley, G.F.; Cane, P.A.; Nokes, D.J. Group- and genotype-specific neutralizing antibody responses against respiratory syncytial virus in infants and young children with severe pneumonia. J. Infect. Dis. 2013, 207, 489–492. [Google Scholar] [CrossRef] [PubMed]
- Waris, M. Pattern of Respiratory Syncytial Virus Epidemics in Finland: Two-Year Cycles with Alternating Prevalence of Groups A and B. J. Infect. Dis. 1991, 163, 464–469. [Google Scholar] [CrossRef] [PubMed]
- Trento, A.; Abrego, L.; Rodríguez-Fernández, R.; González-Sánchez, M.I.; González-Martínez, F.; Delfraro, A.; Pascale, J.M.; Arbiza, J.; Melero, J.A. Conservation of G-Protein Epitopes in Respiratory Syncytial Virus (Group A) Despite Broad Genetic Diversity: Is Antibody Selection Involved in Virus Evolution? J. Virol. 2015, 89, 7776–7785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peret, T.C.; Golub, J.A.; Anderson, L.J.; Hall, C.B.; Schnabel, K.C. Circulation patterns of genetically distinct group A and B strains of human respiratory syncytial virus in a community. J. Virol. 1998, 79, 2221–2229. [Google Scholar] [CrossRef] [PubMed]
- Agoti, C.N.; Otieno, J.R.; Gitahi, C.W.; Cane, P.A.; Nokes, D.J. Rapid Spread and Diversification of Respiratory Syncytial Virus Genotype ON1, Kenya. Emerg. Infect. Dis. 2014, 20, 950–959. [Google Scholar] [CrossRef] [PubMed]
- Eshaghi, A.; Duvvuri, V.R.; Lai, R.; Nadarajah, J.T.; Li, A.; Patel, S.N.; Low, D.E.; Gubbay, J.B. Genetic Variability of Human Respiratory Syncytial Virus A Strains Circulating in Ontario: A Novel Genotype with a 72 Nucleotide G Gene Duplication. PLoS ONE 2012, 7, e32807. [Google Scholar] [CrossRef] [PubMed]
- Trento, A.; Galiano, M.; Videla, C.; Carballal, G.; García-Barreno, B.; Melero, J.A.; Palomo, C. Major changes in the G protein of human respiratory syncytial virus isolates introduced by a duplication of 60 nucleotides. J. Virol. 2003, 84, 3115–3120. [Google Scholar] [CrossRef] [PubMed]
- Hotard, A.L.; Laikhter, E.; Brooks, K.; Hartert, T.V.; Moore, M.L.; Lopez-Ona, A. Functional Analysis of the 60-Nucleotide Duplication in the Respiratory Syncytial Virus Buenos Aires Strain Attachment Glycoprotein. J. Virol. 2015, 89, 8258–8266. [Google Scholar] [CrossRef] [Green Version]
- Botosso, V.F.; Zanotto, P.M.D.A.; Ueda, M.; Arruda, E.; Gilio, A.E.; Vieira, S.E.; Stewien, K.E.; Peret, T.C.T.; Jamal, L.F.; Pardini, M.I.D.M.C.; et al. Positive Selection Results in Frequent Reversible Amino Acid Replacements in the G Protein Gene of Human Respiratory Syncytial Virus. PLOS Pathog. 2009, 5, e1000254. [Google Scholar] [CrossRef]
- Chanock, R.; Finberg, L. Recovery from infants with respiratory illness of a virus related to chimpanzee coryza agent (CCA). II. Epidemiologic aspects of infection in infants and young children. Am. J. Hyg. 1957, 66, 291–300. [Google Scholar]
- Blount, R.E.; Morris, J.A.; Savage, R.E. Recovery of cytopathogenic agent from chimpanzees with coryza. Proc. Soc. Exp. Boil. Med. 1956, 92, 544–549. [Google Scholar]
- Chanock, R.; Finberg, L. Recovery from Infants with Respiratory Illness of A Virus Related To Chimpanzee Coryza Agent (Cca). Am. J. Epidemiol. 1957, 66, 281–290. [Google Scholar] [CrossRef]
- Sullender, W.M. Respiratory Syncytial Virus Genetic and Antigenic Diversity. Clin. Microbiol. Rev. 2000, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.M.; Chanock, R.M.; Rifkind, D.; Kravetz, H.M.; Knight, V. Respiratory syncytial virus. IV. Correlation of virus shedding, serologic response, and illness in adult volunteers. JAMA 1961, 176, 663–667. [Google Scholar]
- Coates, H.V.; Forsyth, B.R.; Chanock, R.M. Biophysical Studies of Respiratory Syncytial Virus I. Density of Respiratory Syncytial Virus and Associated Complement-Fixing Antigens in a Cesium Chloride Density Gradient. J. Bacteriol. 1966, 91, 1263–1269. [Google Scholar] [Green Version]
- Coates, H.V.; Kendrick, L.; Chanock, R.M. Antigenic Differences between Two Strains of Respiratory Syncytial Virus. Exp. Boil. Med. 1963, 112, 958–964. [Google Scholar] [CrossRef]
- Suto, T.; Yano, N.; Ikeda, M.; Miyamoto, M.; Takai, S.; Shigeta, S.; Hinuma, Y.; Ishida, N. Respiratory Syncytial Virus Infection and Its Serologic Epidemiology. Am. J. Epidemiol. 1965, 82, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Doggett, J.E.; Taylor-Robinson, D. Serological studies with respiratory syncytial virus. Arch. Virol. 1965, 15, 601–608. [Google Scholar] [CrossRef]
- Wulff, H.; Kidd, P.; Wenner, H.A. Respiratory Syncytial Virus: Observations on Antigenic Heterogeneity. Exp. Boil. Med. 1964, 115, 240–243. [Google Scholar] [CrossRef]
- Norrby, E.; Marusyk, H.; Örvell, C. Morphogenesis of Respiratory Syncytial Virus in a Green Monkey Kidney Cell Line (Vero). J. Virol. 1970, 6, 237–242. [Google Scholar] [Green Version]
- Prince, G.A.; Jenson, A.B.; Horswood, R.L.; Camargo, E.; Chanock, R.M. The pathogenesis of respiratory syncytial virus infection in cotton rats. Am. J. Pathol. 1978, 93, 771–791. [Google Scholar]
- Prince, G.A.; Porter, D.D. The pathogenesis of respiratory syncytial virus infection in infant ferrets. Am. J. Pathol. 1976, 82, 339–352. [Google Scholar] [PubMed]
- Cavallaro, J.J.; Maassab, H.F. Adaptation of Respiratory Syncytial (RS) Virus to Brain of Suckling Mice. Exp. Boil. Med. 1966, 121, 37–41. [Google Scholar] [CrossRef]
- Hu, B.; Jiang, J.; Zhan, J.; Li, G.; Jiang, Y.; Guan, X.; Chen, Y.; Fang, Z. Development of a reverse genetics system for respiratory syncytial virus long strain and an immunogenicity study of the recombinant virus. Virol. J. 2014, 11, 142. [Google Scholar] [CrossRef]
- Anderson, L.J.; Hierholzer, J.C.; Tsou, C.; Hendry, R.M.; Fernie, B.F.; Stone, Y.; McIntosh, K. Antigenic Characterization of Respiratory Syncytial Virus Strains with Monoclonal Antibodies. J. Infect. Dis. 1985, 151, 626–633. [Google Scholar] [CrossRef]
- Rossey, I.; Sedeyn, K.; Wrapp, D.; Kanekiyo, M.; Chen, M.; Mas, V.; Spitaels, J.; Schepens, B.; Saelens, X.; Gilman, M.S.A.; et al. Potent single-domain antibodies that arrest respiratory syncytial virus fusion protein in its prefusion state. Nat. Commun. 2017, 8, 14158. [Google Scholar] [CrossRef]
- Jones, H.G.; Ritschel, T.; Pascual, G.; Brakenhoff, J.P.J.; Keogh, E.; Furmanova-Hollenstein, P.; Lanckacker, E.; Wadia, J.S.; Gilman, M.S.A.; Williamson, R.A.; et al. Structural basis for recognition of the central conserved region of RSV G by neutralizing human antibodies. PLOS Pathog. 2018, 14, e1006935. [Google Scholar] [CrossRef]
- Lewis, F.A.; Rae, M.L.; Lehmann, N.I.; Ferris, A.A. A syncytial virus associated with epidemic disease of the lower respiratory tract in infants and young children. Med. J. Aust. 1961, 48, 932–933. [Google Scholar]
- Woolums, A.R.; Lee, S.; Moore, M.L. Animal Models of Respiratory Syncytial Virus Pathogenesis and Vaccine Development: Opportunities and Future Directions. RSV 2011. Available online: https://www.intechopen.com/download/pdf/24392 (accessed on 5 May 2019).
- Moore, M.L.; Stokes, K.L.; Hartert, T.V. The impact of viral genotype on pathogenesis and disease severity: respiratory syncytial virus and human rhinoviruses. Curr. Opin. Immunol. 2013, 25, 761–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stokes, K.L.; Chi, M.H.; Sakamoto, K.; Newcomb, D.C.; Currier, M.G.; Huckabee, M.M.; Lee, S.; Goleniewska, K.; Pretto, C.; Williams, J.V.; et al. Differential Pathogenesis of Respiratory Syncytial Virus Clinical Isolates in BALB/c Mice. J. Virol. 2011, 85, 5782–5793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villenave, R.; O’Donoghue, D.; Thavagnanam, S.; Touzelet, O.; Skibinski, G.; Heaney, L.G.; McKaigue, J.P.; Coyle, P.V.; Shields, M.D.; Power, U.F. Differential cytopathogenesis of respiratory syncytial virus prototypic and clinical isolates in primary pediatric bronchial epithelial cells. Virol. J. 2011, 8, 43. [Google Scholar] [CrossRef]
- Moore, M.L.; Chi, M.H.; Luongo, C.; Lukacs, N.W.; Polosukhin, V.V.; Huckabee, M.M.; Newcomb, D.C.; Buchholz, U.J.; Crowe, J.E.; Goleniewska, K.; et al. A Chimeric A2 Strain of Respiratory Syncytial Virus (RSV) with the Fusion Protein of RSV Strain Line 19 Exhibits Enhanced Viral Load, Mucus, and Airway Dysfunction. J. Virol. 2009, 83, 4185–4194. [Google Scholar] [CrossRef] [Green Version]
- Rostad, C.A.; Stobart, C.C.; Todd, S.O.; Molina, S.A.; Lee, S.; Blanco, J.C.G.; Moore, M.L. Enhancing the Thermostability and Immunogenicity of a Respiratory Syncytial Virus (RSV) Live-Attenuated Vaccine by Incorporating Unique RSV Line19F Protein Residues. J. Virol. 2018, 92, e01568-17. [Google Scholar] [CrossRef] [PubMed]
- Herlocher, M.; Ewasyshyn, M.; Sambhara, S.; Gharaee-Kermani, M.; Cho, D.; Lai, J.; Klein, M.; Maassab, H. Immunological properties of plaque purified strains of live attenuated respiratory syncytial virus (RSV) for human vaccine. Vaccine 1999, 17, 172–181. [Google Scholar] [CrossRef]
- Lukacs, N.W.; Moore, M.L.; Rudd, B.D.; Berlin, A.A.; Collins, R.D.; Olson, S.J.; Ho, S.B.; Peebles, R.S. Differential Immune Responses and Pulmonary Pathophysiology Are Induced by Two Different Strains of Respiratory Syncytial Virus. Am. J. Pathol. 2006, 169, 977–986. [Google Scholar] [CrossRef] [Green Version]
- Moore, M.L.; Peebles, R.S. Respiratory syncytial virus disease mechanisms implicated by human, animal model, and in vitro data facilitate vaccine strategies and new therapeutics. Pharmacol. Ther. 2006, 112, 405–424. [Google Scholar] [CrossRef] [PubMed]
- Belshe, R.B.; Anderson, E.L.; Walsh, E.E. Immunogenicity of Purified F Glycoprotein of Respiratory Syncytial Virus: Clinical and Immune Responses to Subsequent Natural Infection in Children. J. Infect. Dis. 1993, 168, 1024–1029. [Google Scholar] [CrossRef]
- Ghildyal, R.; Hogg, G.; Mills, J.; Meanger, J. Detection and subgrouping of respiratory syncytial virus directly from nasopharyngeal aspirates. Clin. Microbiol. Infect. 1997, 3, 120–123. [Google Scholar] [CrossRef] [Green Version]
- Storch, G.A.; Park, C.S. Monoclonal antibodies demonstrate heterogeneity in the G glycoprotein of prototype strains and clinical isolates of respiratory syncytial virus. J. Med Virol. 1987, 22, 345–356. [Google Scholar] [CrossRef]
- Lim, C.S.; Kumarasinghe, G.; Chow, V.T.K. Sequence and phylogenetic analysis of SH, G, and F genes and proteins of Human respiratory syncytial virus isolates from Singapore. Acta Virol. 2003, 47, 97–104. [Google Scholar]
- Walsh, E.E.; Brandriss, M.W.; Schlesinger, J.J. Immunological Differences between the Envelope Glycoproteins of Two Strains of Human Respiratory Syncytial Virus. J. Virol. 1987, 68, 2169–2176. [Google Scholar] [CrossRef]
- Kim, Y.-I.; DeVincenzo, J.P.; Jones, B.G.; Rudraraju, R.; Harrison, L.; Meyers, R.; Cehelsky, J.; Álvarez, R.; Hurwitz, J.L. Respiratory Syncytial Virus Human Experimental Infection Model: Provenance, Production, and Sequence of Low-Passaged Memphis-37 Challenge Virus. PLoS ONE 2014, 9, 113100. [Google Scholar] [CrossRef] [PubMed]
- DeVincenzo, J.P.; Wilkinson, T.; Vaishnaw, A.; Cehelsky, J.; Meyers, R.; Nochur, S.; Harrison, L.; Meeking, P.; Mann, A.; Moane, E.; et al. Viral Load Drives Disease in Humans Experimentally Infected with Respiratory Syncytial Virus. Am. J. Respir. Crit. Care Med. 2010, 182, 1305–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagga, B.; Harrison, L.; Roddam, P.; DeVincenzo, J. Unrecognized prolonged viral replication in the pathogenesis of human RSV infection. J. Clin. Virol. 2018, 106, 1–6. [Google Scholar] [CrossRef]
- DeVincenzo, J.; Lambkin-Williams, R.; Wilkinson, T.; Cehelsky, J.; Nochur, S.; Walsh, E.; Meyers, R.; Gollob, J.; Vaishnaw, A. A randomized, double-blind, placebo-controlled study of an RNAi-based therapy directed against respiratory syncytial virus. Proc. Natl. Acad. Sci. USA 2010, 107, 8800–8805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eyles, J.E.; Johnson, J.E.; Megati, S.; Roopchand, V.; Cockle, P.J.; Weeratna, R.; Makinen, S.; Brown, T.P.; Lang, S.; Witko, S.E.; et al. Nonreplicating Vaccines Can Protect African Green Monkeys from the Memphis 37 Strain of Respiratory Syncytial Virus. J. Infect. Dis. 2013, 208, 319–329. [Google Scholar] [CrossRef]
- Larios Mora, A.; Detalle, L.; Van Geelen, A.; Davis, M.S.; Stohr, T.; Gallup, J.M.; Ackermann, M.R. Kinetics of Respiratory Syncytial Virus (RSV) Memphis Strain 37 (M37) Infection in the Respiratory Tract of Newborn Lambs as an RSV Infection Model for Human Infants. PLoS ONE 2015, 10, e0143580. [Google Scholar] [CrossRef] [Green Version]
- Cui, G.; Zhu, R.; Qian, Y.; Deng, J.; Zhao, L.; Sun, Y.; Wang, F. Genetic Variation in Attachment Glycoprotein Genes of Human Respiratory Syncytial Virus Subgroups A and B in Children in Recent Five Consecutive Years. PLoS ONE 2013, 8, e75020. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, M.A.; Van Niekerk, S.; Tempia, S.; Moyes, J.; Cohen, C.; Madhi, S.A.; Venter, M. Replacement and Positive Evolution of Subtype A and B Respiratory Syncytial Virus G-Protein Genotypes From 1997–2012 in South Africa. J. Infect. Dis. 2013, 208, 227–237. [Google Scholar] [CrossRef]
- Mufson, M.A.; Belshe, R.B.; Örvell, C.; Norrby, E. Respiratory Syncytial Virus Epidemics: Variable Dominance of Subgroups A and B Strains Among Children, 1981–1986. J. Infect. Dis. 1988, 157, 143–148. [Google Scholar] [CrossRef]
- Arnott, A.; Vong, S.; Mardy, S.; Chu, S.; Naughtin, M.; Sovann, L.; Buecher, C.; Beauté, J.; Rith, S.; Borand, L.; et al. A Study of the Genetic Variability of Human Respiratory Syncytial Virus (HRSV) in Cambodia Reveals the Existence of a New HRSV Group B Genotype. J. Clin. Microbiol. 2011, 49, 3504–3513. [Google Scholar] [CrossRef] [Green Version]
- Dapat, I.C.; Shobugawa, Y.; Sano, Y.; Saito, R.; Sasaki, A.; Suzuki, Y.; Kumaki, A.; Zaraket, H.; Dapat, C.; Oguma, T.; et al. New Genotypes within Respiratory Syncytial Virus Group B Genotype BA in Niigata, Japan. J. Clin. Microbiol. 2010, 48, 3423–3427. [Google Scholar] [CrossRef] [Green Version]
- Trento, A.; Viegas, M.; Galiano, M.; Videla, C.; Carballal, G.; Mistchenko, A.S.; Melero, J.A. Natural history of human respiratory syncytial virus inferred from phylogenetic analysis of the attachment (G) glycoprotein with a 60-nucleotide duplication. J. Virol. 2006, 80, 975–984. [Google Scholar] [CrossRef] [PubMed]
- Shobugawa, Y.; Saito, R.; Sano, Y.; Zaraket, H.; Suzuki, Y.; Kumaki, A.; Dapat, I.; Oguma, T.; Yamaguchi, M.; Suzuki, H. Emerging Genotypes of Human Respiratory Syncytial Virus Subgroup A among Patients in Japan. J. Clin. Microbiol. 2009, 47, 2475–2482. [Google Scholar] [CrossRef] [Green Version]
- Tabatabai, J.; Prifert, C.; Pfeil, J.; Grulich-Henn, J.; Schnitzler, P. Novel Respiratory Syncytial Virus (RSV) Genotype ON1 Predominates in Germany during Winter Season 2012–13. PLoS ONE 2014, 9, 109191. [Google Scholar] [CrossRef]
- Auksornkitti, V.; Kamprasert, N.; Thongkomplew, S.; Suwannakarn, K.; Theamboonlers, A.; Samransamruajkij, R.; Poovorawan, Y. Molecular characterization of human respiratory syncytial virus, 2010-2011: Identification of genotype ON1 and a new subgroup B genotype in Thailand. Arch. Virol. 2014, 159, 499–507. [Google Scholar] [CrossRef]
- Khor, C.-S.; Sam, I.-C.; Hooi, P.-S.; Chan, Y.-F. Displacement of predominant respiratory syncytial virus genotypes in Malaysia between 1989 and 2011. Infect. Genet. Evol. 2013, 14, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Liu, H.; Tabor, D.E.; Tovchigrechko, A.; Qi, Y.; Ruzin, A.; Esser, M.T.; Jin, H. Emergence of new antigenic epitopes in the glycoproteins of human respiratory syncytial virus collected from a US surveillance study, 2015-17. Sci. Rep. 2019, 9, 3898. [Google Scholar] [CrossRef]
- Tabatabai, J.; Thielen, A.; Lehners, N.; Daeumer, M.; Schnitzler, P. Respiratory syncytial virus A in haematological patients with prolonged shedding: Premature stop codons and deletion of the genotype ON1 72-nucleotide-duplication in the attachment G gene. J. Clin. Virol. 2018, 98, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Otieno, J.R.; Kamau, E.M.; Oketch, J.W.; Ngoi, J.M.; Gichuki, A.M.; Binter, Š.; Otieno, G.P.; Ngama, M.; Agoti, C.N.; Cane, P.A.; Kellam, P. Whole genome analysis of local Kenyan and global sequences unravels the epidemiological and molecular evolutionary dynamics of RSV genotype ON1 strains. Virus Evol. 2018, 4. [Google Scholar] [CrossRef] [PubMed]
- Abou-El-Hassan, H.; Massaad, E.; Soudani, N.; Assaf-Casals, A.; Shaker, R.; Khoury, M.L.; Ghanem, S.; Karam, M.; Andary, R.; Saito, R.; et al. Detection of ON1 and novel genotypes of human respiratory syncytial virus and emergence of palivizumab resistance in Lebanon. PLoS ONE 2019, 14, e0212687. [Google Scholar] [CrossRef] [PubMed]
- Haider, M.S.H.; Khan, W.H.; Deeba, F.; Ali, S.; Ahmed, A.; Naqvi, I.H.; Dohare, R.; Alsenaidy, H.A.; Alsenaidy, A.M.; Broor, S.; et al. BA9 lineage of respiratory syncytial virus from across the globe and its evolutionary dynamics. PLoS ONE 2018, 13, e0193525. [Google Scholar] [CrossRef]
- Di Giallonardo, F.; Kok, J.; Fernandez, M.; Carter, I.; Geoghegan, J.; Dwyer, D.; Holmes, E.; Eden, J.S. Evolution of Human Respiratory Syncytial Virus (RSV) over Multiple Seasons in New South Wales, Australia. Viruses 2018, 10, 476. [Google Scholar] [CrossRef]
- Fodha, I.; Vabret, A.; Ghedira, L.; Seboui, H.; Chouchane, S.; Dewar, J.; Gueddiche, N.; Trabelsi, A.; Boujaafar, N.; Freymuth, F. Respiratory syncytial virus infections in hospitalized infants: Association between viral load, virus subgroup, and disease severity. J. Med Virol. 2007, 79, 1951–1958. [Google Scholar] [CrossRef]
- Otieno, J.R.; Kamau, E.M.; Agoti, C.N.; Lewa, C.; Otieno, G.; Bett, A.; Ngama, M.; Cane, P.A.; Nokes, D.J. Spread and Evolution of Respiratory Syncytial Virus A Genotype ON1, Coastal Kenya, 2010–2015. Emerg. Infect. Dis. 2017, 23, 264–271. [Google Scholar] [CrossRef]
- Esposito, S.; Piralla, A.; Zampiero, A.; Bianchini, S.; Di Pietro, G.; Scala, A.; Pinzani, R.; Fossali, E.; Baldanti, F.; Principi, N. Characteristics and Their Clinical Relevance of Respiratory Syncytial Virus Types and Genotypes Circulating in Northern Italy in Five Consecutive Winter Seasons. PLoS ONE 2015, 10, 0129369. [Google Scholar] [CrossRef]
- White, L.J.; Waris, M.; Cane, P.A.; Nokes, D.J.; Medley, G.F.; Medley, G. The transmission dynamics of groups A and B human respiratory syncytial virus (hRSV) in England & Wales and Finland: seasonality and cross-protection. Epidemiol. Infect. 2005, 133, 279–289. [Google Scholar]
- Vandini, S.; Biagi, C.; Lanari, M. Respiratory Syncytial Virus: The Influence of Serotype and Genotype Variability on Clinical Course of Infection. Int. J. Mol. Sci. 2017, 18, 1717. [Google Scholar] [CrossRef]
- Hall, C.B.; Walsh, E.E.; Schnabel, K.C.; Long, C.E.; McConnochie, K.M.; Hildreth, S.W.; Anderson, L.J. Occurrence of Groups A and B of Respiratory Syncytial Virus over 15 Years: Associated Epidemiologic and Clinical Characteristics in Hospitalized and Ambulatory Children. J. Infect. Dis. 1990, 162, 1283–1290. [Google Scholar] [CrossRef]
- Agoti, C.N.; Mwihuri, A.G.; Sande, C.J.; Onyango, C.O.; Medley, G.F.; Cane, P.A.; Nokes, D.J. Genetic Relatedness of Infecting and Reinfecting Respiratory Syncytial Virus Strains Identified in a Birth Cohort from Rural Kenya. J. Infect. Dis. 2012, 206, 1532–1541. [Google Scholar] [CrossRef]
- Rodriguez-Fernandez, R.; Tapia, L.I.; Yang, C.-F.; Torres, J.P.; Chavez-Bueno, S.; Garcia, C.; Jaramillo, L.M.; Moore-Clingenpeel, M.; Jafri, H.S.; Peeples, M.E.; et al. Respiratory Syncytial Virus Genotypes, Host Immune Profiles, and Disease Severity in Young Children Hospitalized with Bronchiolitis. J. Infect. Dis. 2017, 217, 24–34. [Google Scholar] [CrossRef]
- Yoshihara, K.; Le, M.N.; Okamoto, M.; Wadagni, A.C.A.; Nguyen, H.A.; Toizumi, M.; Pham, E.; Suzuki, M.; Nguyen, A.T.T.; Oshitani, H.; et al. Association of RSV-A ON1 genotype with Increased Pediatric Acute Lower Respiratory Tract Infection in Vietnam. Sci. Rep. 2016, 6, 27856. [Google Scholar] [CrossRef] [Green Version]
- Laham, F.R.; Mansbach, J.M.; Piedra, P.A.; Hasegawa, K.; Sullivan, A.F.; Espinola, J.A.; Camargo, C.A. Clinical Profiles of Respiratory Syncytial Virus Subtypes A and B among Children Hospitalized with Bronchiolitis. Pediatr. Infect. J. 2017, 36, 808–810. [Google Scholar] [CrossRef]
- Martinello, R.A.; Chen, M.D.; Weibel, C.; Kahn, J.S. Correlation between Respiratory Syncytial Virus Genotype and Severity of Illness. J. Infect. Dis. 2002, 186, 839–842. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Peeples, M.E.; Boucher, R.C.; Collins, P.L.; Pickles, R.J. Respiratory Syncytial Virus Infection of Human Airway Epithelial Cells Is Polarized, Specific to Ciliated Cells, and without Obvious Cytopathology. J. Virol. 2002, 76, 5654–5666. [Google Scholar] [CrossRef] [Green Version]
- Groves, H.E.; Guo-Parke, H.; Broadbent, L.; Shields, M.D.; Power, U.F. Characterisation of morphological differences in well-differentiated nasal epithelial cell cultures from preterm and term infants at birth and one-year. PLoS ONE 2018, 13, e0201328. [Google Scholar] [CrossRef] [PubMed]
- Villenave, R.; Thavagnanam, S.; Sarlang, S.; Parker, J.; Douglas, I.; Skibinski, G.; Heaney, L.G.; McKaigue, J.P.; Coyle, P.V.; Shields, M.D.; et al. In vitro modeling of respiratory syncytial virus infection of pediatric bronchial epithelium, the primary target of infection in vivo. Proc. Natl. Acad. Sci. USA 2012, 109, 5040–5045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, T.E.; Guzman, K.; Davis, C.W.; Abdullah, L.H.; Nettesheim, P. Mucociliary differentiation of serially passaged normal human tracheobronchial epithelial cells. Am. J. Respir. Cell Mol. Boil. 1996, 14, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Drajac, C.; Laubreton, D.; Riffault, S.; Descamps, D. Pulmonary Susceptibility of Neonates to Respiratory Syncytial Virus Infection: A Problem of Innate Immunity? J. Immunol. 2017, 2017, 1–13. [Google Scholar] [CrossRef]



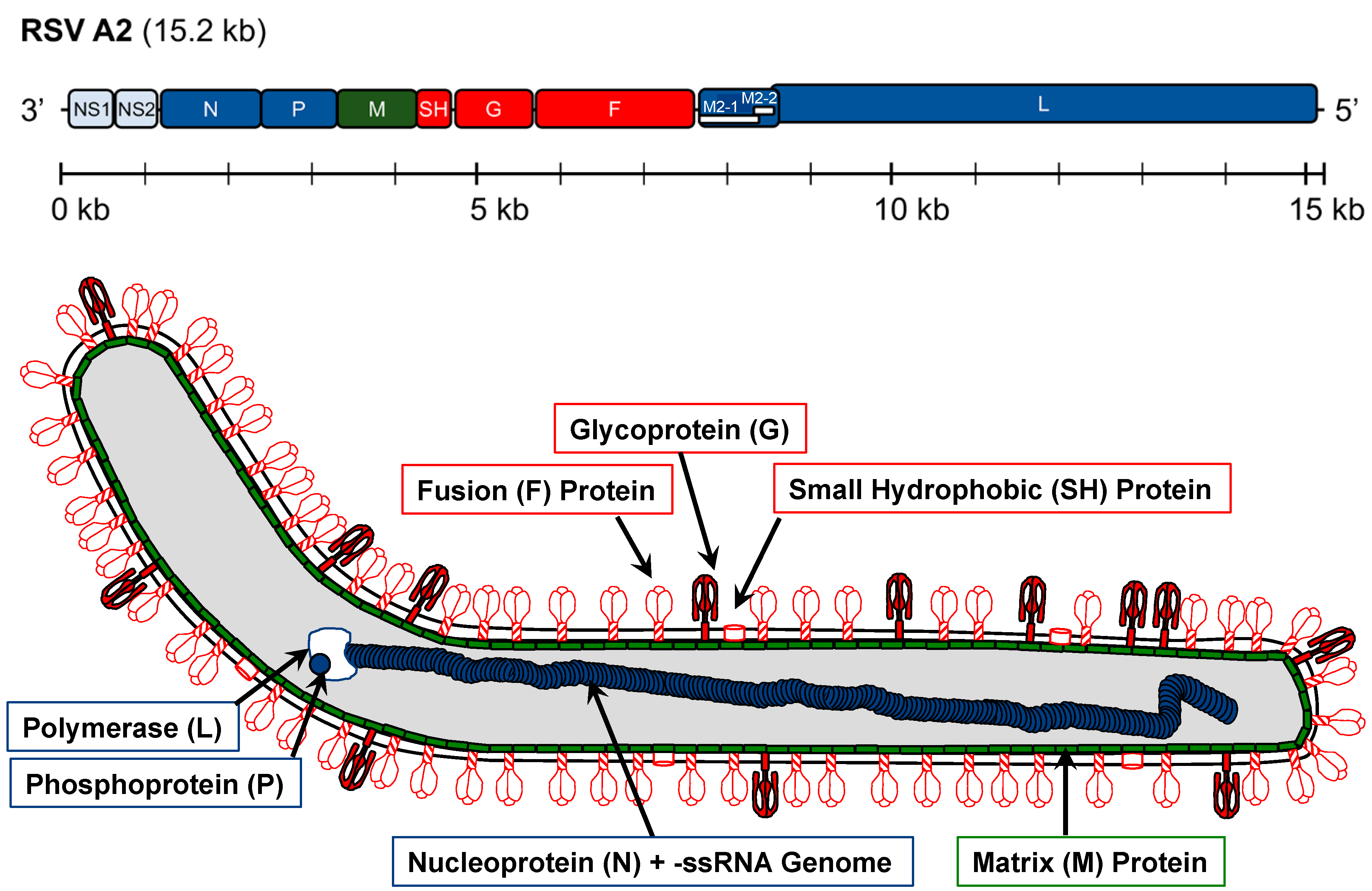{kind=link}
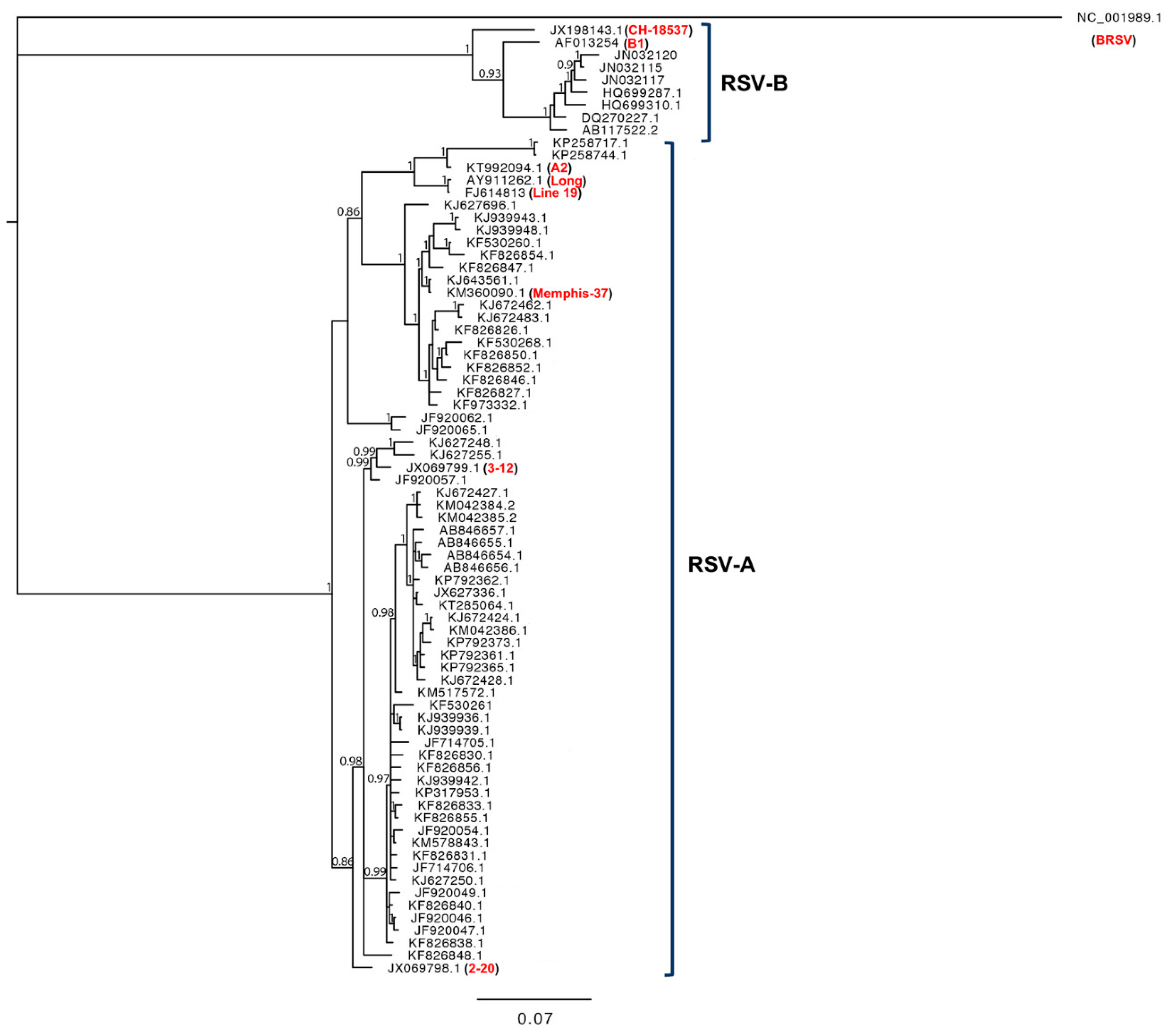{kind=link}
| Virus Strain | Type and Designation | Characteristics and Applications |
|---|---|---|
| Long | RSV-A (Laboratory Strain) |
|
| A2 | RSV-A (Laboratory Strain) |
|
| Line 19/A2-line19F | RSV-A (Laboratory Strain) |
|
| CH 18537 | RSV-B (Laboratory Strain) |
|
| Memphis-37 | RSV-A (Laboratory Strain) |
|
| Clinical Isolates | RSV-A and RSV-B Isolates |
|
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pandya, M.C.; Callahan, S.M.; Savchenko, K.G.; Stobart, C.C. A Contemporary View of Respiratory Syncytial Virus (RSV) Biology and Strain-Specific Differences. Pathogens 2019, 8, 67. https://doi.org/10.3390/pathogens8020067
Pandya MC, Callahan SM, Savchenko KG, Stobart CC. A Contemporary View of Respiratory Syncytial Virus (RSV) Biology and Strain-Specific Differences. Pathogens. 2019; 8(2):67. https://doi.org/10.3390/pathogens8020067
Chicago/Turabian StylePandya, Mansi C., Sean M. Callahan, Kyryll G. Savchenko, and Christopher C. Stobart. 2019. "A Contemporary View of Respiratory Syncytial Virus (RSV) Biology and Strain-Specific Differences" Pathogens 8, no. 2: 67. https://doi.org/10.3390/pathogens8020067
APA StylePandya, M. C., Callahan, S. M., Savchenko, K. G., & Stobart, C. C. (2019). A Contemporary View of Respiratory Syncytial Virus (RSV) Biology and Strain-Specific Differences. Pathogens, 8(2), 67. https://doi.org/10.3390/pathogens8020067