A Siderophore Analog of Fimsbactin from Acinetobacter Hinders Growth of the Phytopathogen Pseudomonas syringae and Induces Systemic Priming of Immunity in Arabidopsis thaliana



Abstract
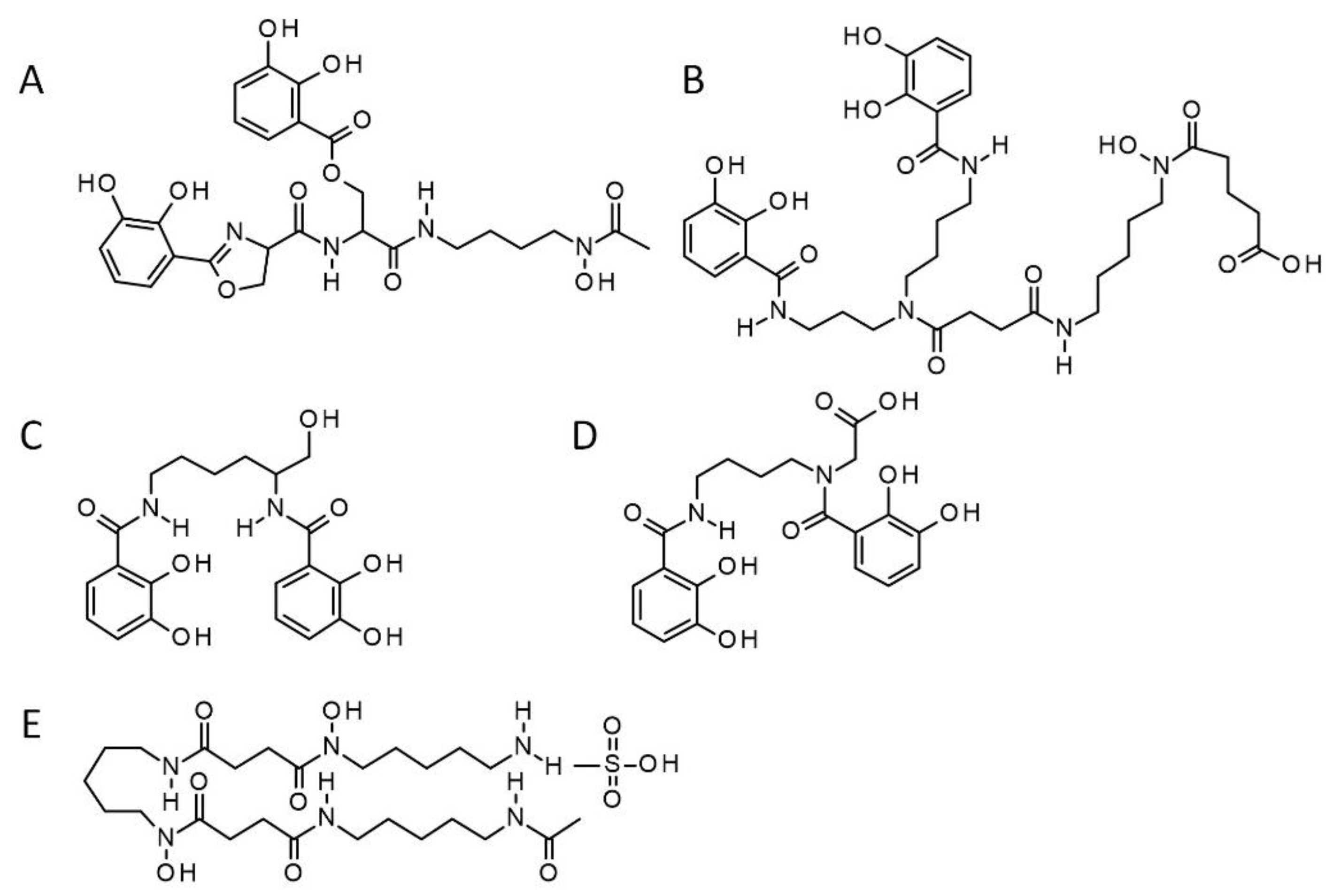
:1. Introduction
2. Results
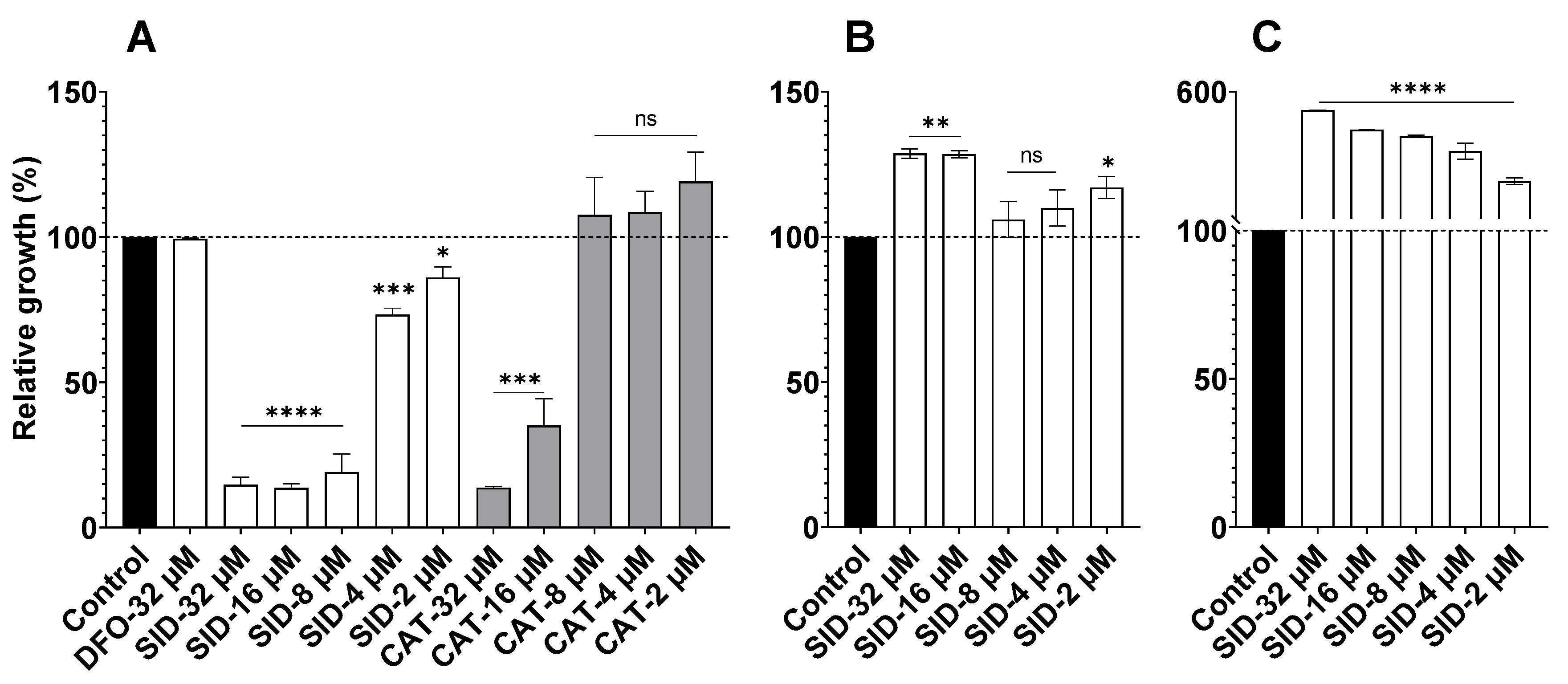
2.1. SID Inhibits Pst DC3000 but not B. subtilis or A. baumannii Growth In Vitro
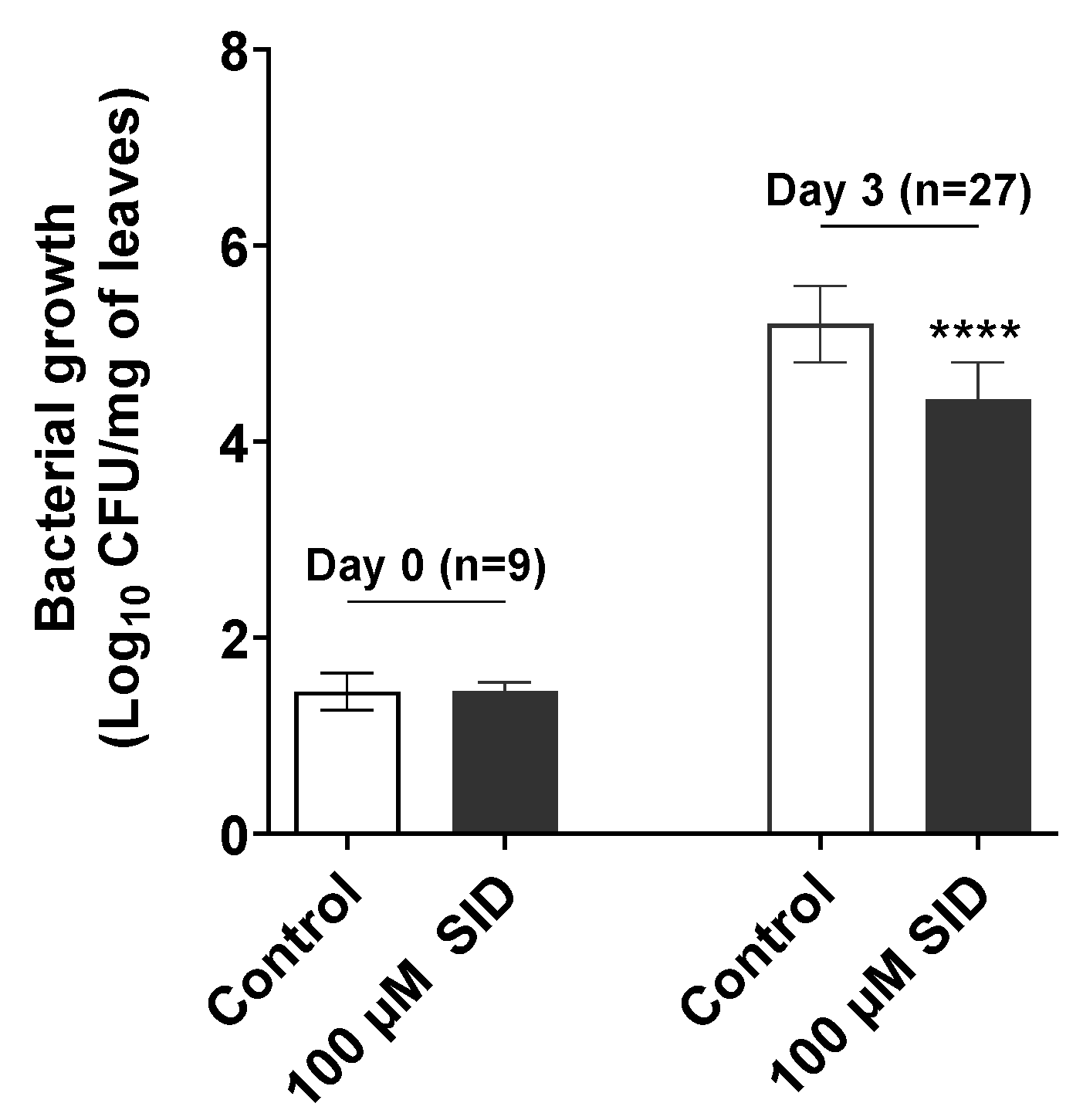
2.2. SID Promotes Priming of Arabidopsis Systemic Resistance against Pst DC3000
2.3. SA Pathway is Primed by SID
3. Discussion
4. Materials and Methods
4.1. Synthetic Siderophores
4.2. In Vitro Growth Inhibition of Pseudomonas Syringae Pv Tomato DC3000 (Pst DC3000) by Synthetic Siderophores
4.3. SID-induced Resistance of Arabidopsis against Pst DC3000
4.4. Characterization of Immunity Markers Induced by SID in Arabidopsis
4.5. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weinberg, E.D. The Lactobacillus Anomaly: Total Iron Abstinence. Perspect. Biol. Med. 1997, 40, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Troxell, B.; Xu, H.; Yang, X.F. Borrelia burgdorferi, a pathogen that lacks iron, encodes manganese-dependent superoxide dismutase essential for resistance to streptonigrin. J. Biol. Chem. 2012, 287, 19284–19293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomine, S.; Lanquar, V. Iron Transport and Signaling in Plants. Transporters and Pumps in Plant. Signaling, 1st ed.; Geisler, M., Venema, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Staiger, D. Chemical strategies for iron acquisition in plants. Angew. Chem. Int. Ed. Engl. 2002, 41, 2259–2264. [Google Scholar] [CrossRef]
- Morrissey, J.; Guerinot, M.L. Iron uptake and transport in plants: The good, the bad, and the ionome. Chem. Rev. 2009, 109, 4553–4567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aznar, A.; Dellagi, A. New insights into the role of siderophores as triggers of plant immunity: What can we learn from animals? J. Exp. Bot. 2015, 66, 3001–3010. [Google Scholar] [CrossRef] [Green Version]
- Franza, T.; Expert, D. Role of iron homeostasis in the virulence of phytopathogenic bacteria: An “à la carte” menu. Mol. Plant. Pathol. 2013, 14, 429–438. [Google Scholar] [CrossRef]
- Oide, S.; Moeder, W.; Krasnoff, S.; Gibson, D.; Haas, H.; Yoshioka, K.; Turgeon, B.G. NPS6, encoding a Nonribosomal Peptide Synthetase Involved in Siderophore-Mediated Iron Metabolism, Is a Conserved Virulence Determinant of Plant Pathogenic Ascomycetes. Plant Cell 2006, 18, 2836–2853. [Google Scholar] [CrossRef] [Green Version]
- Dellagi, A.; Rigault, M.; Segond, D.; Roux, C.; Kraepiel, Y.; Cellier, F.; Briat, J.-F.; Gaymard, F.; Expert, D. Siderophore-mediated upregulation of Arabidopsis ferritin expression in response to Erwinia chrysanthemi infection. Plant J. 2005, 43, 262–272. [Google Scholar] [CrossRef]
- Taguchi, F.; Suzuki, T.; Inagaki, Y.; Toyoda, K.; Shiraishi, T.; Ichinose, Y. The siderophore pyoverdine of Pseudomonas syringae pv. tabaci 6605 is an intrinsic virulence factor in host tobacco infection. J. Bacteriol. 2010, 192, 117–126. [Google Scholar]
- Flo, T.H.; Smith, K.D.; Sato, S.; Rodriguez, D.J.; Holmes, M.A.; Strong, R.K.; Akira, S.; Aderem, M. Lipocalin 2 mediates an innate immune response to bacterial infection by sequestrating iron. Nature 2004, 432, 917–921. [Google Scholar] [CrossRef]
- Charron, J.-B.F.; Ouellet, F.; Houde, M.; Sarhan, F. The plant Apolipoprotein D ortholog protects Arabidopsis against oxidative stress. BMC Plant. Biol. 2008, 8, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lugtenberg, B.; Kamilova, F. Plant-Growth-Promoting Rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieterse, C.M.J.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal modulation of plant immunity. Annu Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boller, T.; Felix, G. A Renaissance of Elicitors: Perception of Microbe-Associated Molecular Patterns and Danger Signals by Pattern-Recognition Receptors. Annu. Rev. Plant. Biol. 2009, 60, 379–406. [Google Scholar] [CrossRef] [PubMed]
- Meziane, H.; Van Der Sluis, I.; Van Loon, L.C.; Höfte, M.; Bakker, P.A. Determinants of Pseudomonas putida WCS358 involved in inducing systemic resistance in plants. Mol. Plant. Pathol. 2005, 6, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Dodds, P.N.; Rathjen, J.P. Plant immunity: Towards an integrated view of plant-pathogen interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef]
- Tsuda, K.; Somssich, I.E. Transcriptional networks in plant immunity. New Phytol. 2015, 206, 932–947. [Google Scholar] [CrossRef]
- Kuan, K.B.; Othman, R.; Abdul Rahim, K.; Shamsuddin, Z.H. Plant growth-promoting rhizobacteria inoculation to enhance vegetative growth, nitrogen fixation and nitrogen remobilisation of maize under greenhouse conditions. PLoS ONE 2016, 11, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Proschak, A.; Lubuta, P.; Grün, P.; Löhr, F.; Wilharm, G.; De Berardinis, V.; Bode, H.B. Structure and Biosynthesis of Fimsbactins A-F, Siderophores from Acinetobacter baumannii and Acinetobacter baylyi. ChemBioChem 2013, 14, 633–638. [Google Scholar] [CrossRef]
- Bohac, T.J.; Fang, L.; Giblin, D.E.; Wencewicz, T.A. Fimsbactin and Acinetobactin Compete for the Periplasmic Siderophore Binding Protein BauB in Pathogenic Acinetobacter baumannii. ACS Chem. Biol. 2019, 14, 674–687. [Google Scholar] [CrossRef]
- Wencewicz, T.A.; Miller, M.J. Biscatecholate-monohydroxamate mixed ligand siderophore-carbacephalosporin conjugates are selective sideromycin antibiotics that target Acinetobacter baumannii. J. Med. Chem. 2013, 56, 4044–4052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunze, B.; Bedorf, N.; Kohl, W.; Höfle, G.; Reichenbach, H. Myxochelin A, A New Iron-chelating Compound from Angiococcus disciformis (Myxobacterales). J. Antibiot. 1989, 42, 14–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, M.; Lin, Y.M.; Miller, P.A.; Möllmann, U.; Boggess, W.; Miller, M.J. Siderophore Conjugates of Daptomycin are Potent Inhibitors of Carbapenem Resistant Strains of Acinetobacter baumannii. ACS Infect. Dis. 2018, 4, 1529–1535. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S.C.; Robinson, A.K.; Rodríguez-Quiñones, F. Bacterial iron homeostasis. FEMS Microbiol. Rev. 2003, 27, 215–237. [Google Scholar] [CrossRef]
- Aznar, A.; Chen, N.W.G.; Rigault, M.; Riache, N.; Joseph, D.; Desmaële, D.; Mouille, G.; Boutet, S.; Soubigou-Taconnat, L.; Renou, J.-P.; et al. Scavenging Iron: A Novel Mechanism of Plant Immunity Activation by Microbial Siderophores. Plant. Physiol. 2014, 164, 2167–2183. [Google Scholar] [CrossRef] [Green Version]
- Dellagi, A.; Segond, D.; Rigault, M.; Fagard, M.; Simon, C.; Saindrenan, P.; Expert, D. Microbial Siderophores Exert a Subtle Role in Arabidopsis during Infection by Manipulating the Immune Response and the Iron Status. Plant. Physiol. 2009, 150, 1687–1696. [Google Scholar] [CrossRef] [Green Version]
- Vives-Peris, V.; de Ollas, C.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Root exudates: From plant to rhizosphere and beyond. Plant. Cell Rep. 2019. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced Systemic Resistance by Beneficial Microbes. Ann. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [Green Version]
- Damalas, C.A.; Eleftherohorinos, I.G. Pesticide exposure, safety issues, and risk assessment indicators. Int. J. Environ. Res. Public Health 2011, 8, 1402–1419. [Google Scholar] [CrossRef]
- Crowley, D.E. Microbial Siderophores in the Plant Rhizosphere. In Iron Nutrition in Plants and Rhizospheric Microorganisms, 1st ed.; Barton, L.L., Abadia, J., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 169–198. [Google Scholar] [CrossRef]
- Glick, B.R. The enhancement of plant growth by free-living bacteria. Can. J. Microbiol. 1995, 41, 109–117. [Google Scholar] [CrossRef]
- Grandchamp, G.M.; Caro, L.; Shank, E. Pirated Siderophores Promote Sporulation in Bacillus subtilis. Appl. Environ. Microbiol. 2017, 83, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Expert, D.; Franza, T.; Dellagi, A.; Walker, J.M. Iron in Plant-Pathogen Interactions. Molecular Aspects of Iron Metabolism in Pathogenic and Symbiotic Plant.-Microbes Associations, 1st ed.; Expert, D., O’Brian, M.R., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 7–39. Available online: https://link.springer.com/chapter/10.1007/978-94-007-5267-2_2 (accessed on 24 August 2012).
- Leeman, M.; Den Ouden, E.M.; Van Pelt, J.A.; Dirkx, F.P.M.; Steijl, H.; Bakker, P.A.H.M.; Schippers, B. Iron availability affects induction of systemic resistance to Fusarium wilt of radish by Pseudomonas fluorescens. Phytopathol. 1996, 86, 149–155. [Google Scholar] [CrossRef]
- Kieu, N.P.; Aznar, A.; Segond, D.; Rigault, M.; Simond-Côte, E.; Kunz, C.; Soulie, M.-C.; Expert, D.; Dellagi, A. Iron deficiency affects plant defence responses and confers resistance to Dickeya dadantii and Botrytis cinerea. Mol. Plant. Pathol. 2012, 13, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Yang, Y.; Liu, K.; Zhang, L.; Guo, H.; Sun, T.; Wang, H. Involvement of endogenous salicylic acid in iron-deficiency responses in Arabidopsis. J. Exp. Bot. 2016, 67, 4179–4193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Lamothe, R.; El Oirdi, M.; Brisson, N.; Bouarab, K. The conjugated auxin indole-3-acetic acid-aspartic acid promotes plant disease development. Plant Cell 2012, 24, 672–777. [Google Scholar] [CrossRef]
- Swift, M.L. GraphPad prism, data analysis, and scientific graphing. J. Chem. Inf. Comput. Sci. 1997, 37, 411–412. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | GenBank Accession | Pathway | Primer Sequence |
|---|---|---|---|
| AtPR1 | AT2G14610 | Salicylic acid | F: CTCATACACTCTGGTGGG |
| R: TTGGCACATCCGAGTC | |||
| AtPDF1.2 | AT5G44420 | Jasmonic acid | F: TCTTTGGTGCTAAATCGTGTGT |
| R: TGTAACAACAACGGGAAAATAAACA | |||
| AtEF1 | AT5G60390 | Elongation factor Tu | F: TCTCCGAGTACCCACCTTTGR: TCCTTCTTGTCCACGCTCTT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Betoudji, F.; Abd El Rahman, T.; Miller, M.J.; Ghosh, M.; Jacques, M.; Bouarab, K.; Malouin, F. A Siderophore Analog of Fimsbactin from Acinetobacter Hinders Growth of the Phytopathogen Pseudomonas syringae and Induces Systemic Priming of Immunity in Arabidopsis thaliana. Pathogens 2020, 9, 806. https://doi.org/10.3390/pathogens9100806
Betoudji F, Abd El Rahman T, Miller MJ, Ghosh M, Jacques M, Bouarab K, Malouin F. A Siderophore Analog of Fimsbactin from Acinetobacter Hinders Growth of the Phytopathogen Pseudomonas syringae and Induces Systemic Priming of Immunity in Arabidopsis thaliana. Pathogens. 2020; 9(10):806. https://doi.org/10.3390/pathogens9100806
Chicago/Turabian StyleBetoudji, Fabrice, Taha Abd El Rahman, Marvin J. Miller, Manuka Ghosh, Mario Jacques, Kamal Bouarab, and François Malouin. 2020. "A Siderophore Analog of Fimsbactin from Acinetobacter Hinders Growth of the Phytopathogen Pseudomonas syringae and Induces Systemic Priming of Immunity in Arabidopsis thaliana" Pathogens 9, no. 10: 806. https://doi.org/10.3390/pathogens9100806
APA StyleBetoudji, F., Abd El Rahman, T., Miller, M. J., Ghosh, M., Jacques, M., Bouarab, K., & Malouin, F. (2020). A Siderophore Analog of Fimsbactin from Acinetobacter Hinders Growth of the Phytopathogen Pseudomonas syringae and Induces Systemic Priming of Immunity in Arabidopsis thaliana. Pathogens, 9(10), 806. https://doi.org/10.3390/pathogens9100806