Two New Sturgeon Species are Susceptible to Acipenser Iridovirus European (AcIV-E) Infection

 ,
,  ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Sampling: Starry Sturgeon
2.2. Fish Sampling: Sterlet
2.3. Laboratory Exams
2.4. DNA Extraction and Real-Time PCR
2.5. MPC Gene Sequencing and Phylogenetic Analysis
3. Results
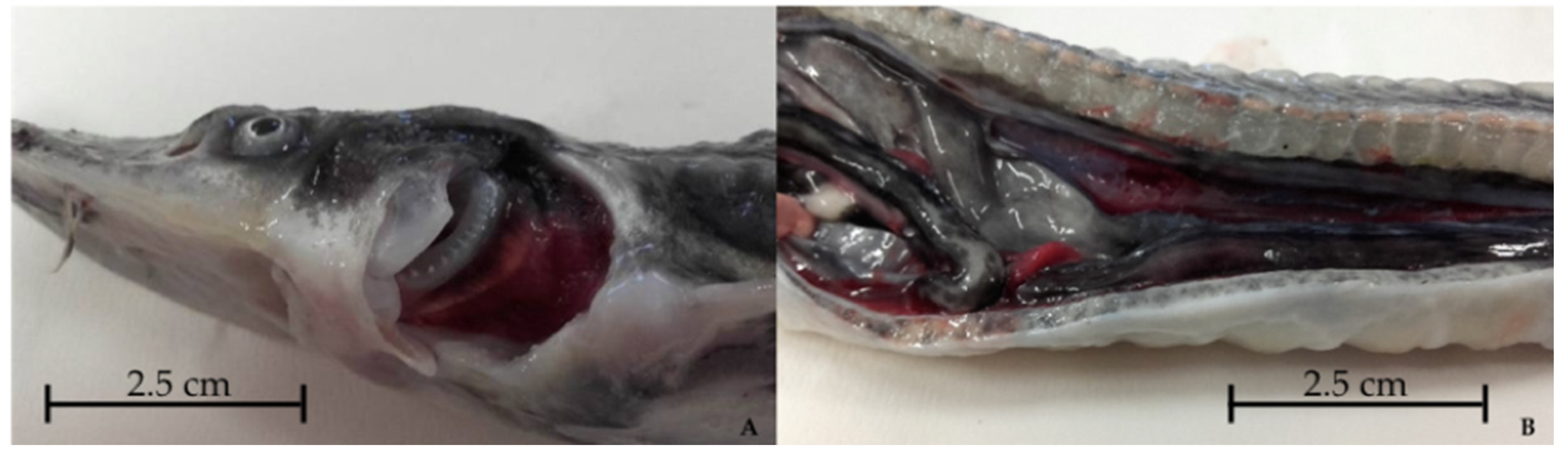
3.1. Starry Sturgeon
3.2. Sterlet
3.3. Phylogenetic Analysis
4. Discussion and Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ciulli, S.; Volpe, E.; Sirri, R.; Passalacqua, P.L.; Cesa Bianchi, F.; Serratore, P.; Mandrioli, L. Outbreak of mortality in Russian (Acipenser gueldenstaedtii) and Siberian (Acipenser baerii) sturgeons associated with sturgeon nucleo-cytoplasmatic large DNA virus. Vet. Microbiol. 2016, 191, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Bigarré, L.; Lesne, M.; Lautraite, A.; Chesneau, V.; Leroux, A.; Jamin, M.; Boitard, P.M.; Toffan, A.; Prearo, M.; Labrut, S.; et al. Molecular identification of iridoviruses infecting various sturgeon species in Europe. J. Fish Dis. 2017, 40, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Hick, P.; Becker, J.; Whittington, R. Iridoviruses of fish. In Aquaculture Virology; Kibenge, F.S.B., Godoy, M.G., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 127–152. [Google Scholar]
- Clouthier, S.; Anderson, E.; Kurath, G.; Breyta, R. Molecular systematics of sturgeon nucleocytoplasmic large DNA viruses. Mol. Phylogenet. Evol. 2018, 128, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Pallandre, L.; Lesne, M.; de Boisséson, C.; Briand, F.X.; Charrier, A.; Waltzek, T.; Daniel, P.; Tragnan, A.; Debeuf, B.; Chesneau, V.; et al. Acipenser iridovirus-European encodes a replication factor C (RFC) sub-unit. Arch. Virol. 2018, 163, 2985–2995. [Google Scholar] [CrossRef] [PubMed]
- Pallandre, L.; Lesne, M.; de Boisséson, C.; Charrier, A.; Daniel, P.; Tragnan, A.; Debeuf, B.; Chesneau, V.; Bigarré, L. Genetic identification of two Acipenser iridovirus-European variants using high-resolution melting analysis. J. Virol. Methods. 2019, 265, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Axén, C.; Vendramin, N.; Toffan, A. Outbreak of mortality associated with Acipenser Iridovirus European (AcIV-E) detection in Siberian Sturgeon (Acipenser baerii) farmed in Sweden. Fishes 2018, 3, 42. [Google Scholar] [CrossRef] [Green Version]
- Prearo, M.; Pedron, C.; Caruso, C.; Pastorino, P.; Masoero, L.; Burioli, E.; Bigarrè, L.; Toffan, A. Acipenser Iridovirus-European (AcIV-E) in Russian Sturgeon farmed in Italy: cases description. In Proceedings of the 18th International Conference for the European Association of Fish Pathologists, Belfast, Northern Ireland, 4–7 September 2017; Abstract 068-P. p. 286. [Google Scholar]
- Caruso, C.; Pedron, C.; Prato, R.; Toffan, A.; Bigarré, L.; Pastorino, P.; Burioli, E.A.V.; Righetti, M.; Foglini, C.; Masoero, L.; et al. Sturgeon Iridovirus: dal focolaio alla diagnosi di laboratorio. In Proceedings of the XXII Convegno Nazionale S.I.P.I. (Società Italiana Patologia Ittica), Torino, Italy, 8–9 september 2016; p. 16. [Google Scholar]
- Sievers, F.; Higgins, D.G. Clustal Omega, Accurate Alignment of Very Large Numbers of Sequences. In Multiple Sequence Alignment Methods; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2013; Volume 1079, pp. 105–116. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Ciulli, S.; Pedron, C.; Toffan, A. Malattie virali: sintomatologia e diagnosi di laboratorio. In Proceedings of the XXIV Convegno Nazionale S.I.P.I. (Società Italiana Patologia Ittica), San Michele all’Adige, Italy, 11–13 october 2018; p. 78. [Google Scholar]
- Soto, E.; Richey, C.; Stevens, B.; Yun, S.; Kenelty, K.; Reichley, S.; Griffin, M.; Kurobe, T.; Camus, A. Co-infection of Acipenserid herpesvirus 2 (AciHV-2) and Streptococcus iniae in cultured white sturgeon Acipenser transmontanus. Dis. Aquat. Org. 2017, 124, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Low, C.F.; Syarul Nataqain, B.; Chee, H.Y.; Rozaini, M.Z.H.; Najiah, M. Betanodavirus: Dissection of the viral life cycle. J. Fish. Dis. 2017, 40, 1489–1496. [Google Scholar] [CrossRef] [PubMed]
- Santi, M.; Pastorino, P.; Foglini, C.; Righetti, M.; Pedron, C.; Prearo, M. A survey of bacterial infections in sturgeon farming in Italy. J. Appl. Ichthyol. 2018, 35, 275–282. [Google Scholar] [CrossRef] [Green Version]
- Bowman, J.P. Genus XIII. Shewanella MacDonell and Colwell 1986. In Bergey’s Manual of Systematics Bacteriology 2nd Edition, Volume Two The Proteobacteria, Part B The Gammaproteobacteria; Garrity, G., Brenner, D.J., Krieg, N.R., Staley, J.T., Eds.; Springer: New York, NY, USA, 2005; pp. 480–490. ISBN 978-0-387-24144-9. [Google Scholar]
- Rusev, V.; Rusenova, N.; Simeonov, R.; Stratev, D. Staphylococcus warneri and Shewanella putrefaciens Co-infection in Siberian Sturgeon (Acipenser baerii) and Hybrid Sturgeon (Huso huso x Acipenser baerii). J. Microbiol. Exp. 2016, 3, 00078. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specimen | Gross Pathology | Parasitological Analysis | Bacteriological Analysis | qPCR (Ct) | End Point PCR |
|---|---|---|---|---|---|
| 37849/1 | abdominal swelling, increased gill volume, fatty liver, atrophy of internal organs | - | - | + (35.49) | + |
| 37849/2 | abdominal swelling, increased gill volume, fatty liver | - | - | + (33.06) | + |
| 37849/3 | abdominal swelling, increased gill volume, fatty liver | -- | - | + (36.92) | + |
| 37849/4 | abdominal swelling, increased gill volume, fatty liver, splenomegaly | - | - | + (32.85) | + |
| 37849/5 | abdominal swelling, increased gill volume, fatty liver, splenomegaly | - | - | + (32.59) | + |
| C + | // | // | // | + (18.32) | // |
| C – | // | // | // | - (no Ct) | // |
| Specimen | Gross Pathology | Parasitological Analysis | Bacteriological Analysis | qPCR (Ct) | End Point PCR |
|---|---|---|---|---|---|
| 101262/1 | thinness, congested gills, hepatic hypoplasia | - | - | + (25.44) | + |
| 101262/2 | thinness, congested gills | - | - | + (25.73) | + |
| 101262/3 | thinness, congested gills, hepatic hypoplasia | -- | - | + (22.76) | + |
| 101262/4 | thinness, congested gills | - | - | + (25.65) | + |
| 101262/5 | thinness, congested gills | - | Shewanella putrefaciens | + (23.42) | + |
| C + | // | // | // | + (16.77) | // |
| C - | // | // | // | - (no Ct) | // |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mugetti, D.; Pastorino, P.; Menconi, V.; Messina, M.; Masoero, L.; Ceresa, L.; Pedron, C.; Prearo, M. Two New Sturgeon Species are Susceptible to Acipenser Iridovirus European (AcIV-E) Infection. Pathogens 2020, 9, 156. https://doi.org/10.3390/pathogens9030156
Mugetti D, Pastorino P, Menconi V, Messina M, Masoero L, Ceresa L, Pedron C, Prearo M. Two New Sturgeon Species are Susceptible to Acipenser Iridovirus European (AcIV-E) Infection. Pathogens. 2020; 9(3):156. https://doi.org/10.3390/pathogens9030156
Chicago/Turabian StyleMugetti, Davide, Paolo Pastorino, Vasco Menconi, Margherita Messina, Loretta Masoero, Luisa Ceresa, Claudio Pedron, and Marino Prearo. 2020. "Two New Sturgeon Species are Susceptible to Acipenser Iridovirus European (AcIV-E) Infection" Pathogens 9, no. 3: 156. https://doi.org/10.3390/pathogens9030156
APA StyleMugetti, D., Pastorino, P., Menconi, V., Messina, M., Masoero, L., Ceresa, L., Pedron, C., & Prearo, M. (2020). Two New Sturgeon Species are Susceptible to Acipenser Iridovirus European (AcIV-E) Infection. Pathogens, 9(3), 156. https://doi.org/10.3390/pathogens9030156