
Genomic Analyses of Weissella cibaria W25, a Potential Bacteriocin-Producing Strain Isolated from Pasture in Campos das Vertentes, Minas Gerais, Brazil

 , ,
, ,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain
2.2. Genome Sequencing and Assembly
2.3. Genome Annotation and Analysis
2.4. Phylogenetic Analyses
2.5. Comparative Genomic
2.6. Availability of Nucleotide Sequence Data
3. Results and Discussion
3.1. Genome Sequencing, Annotation and Analysis
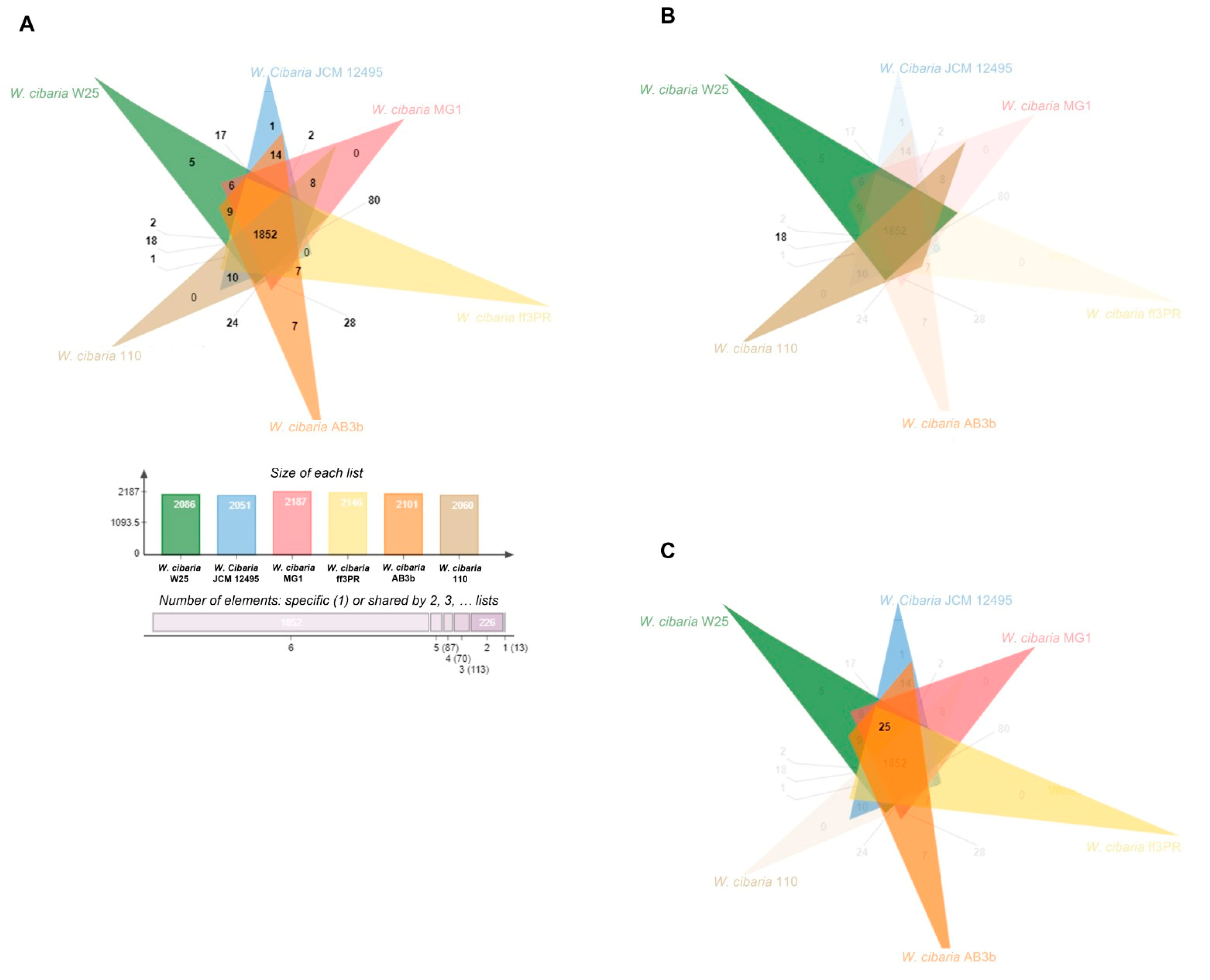
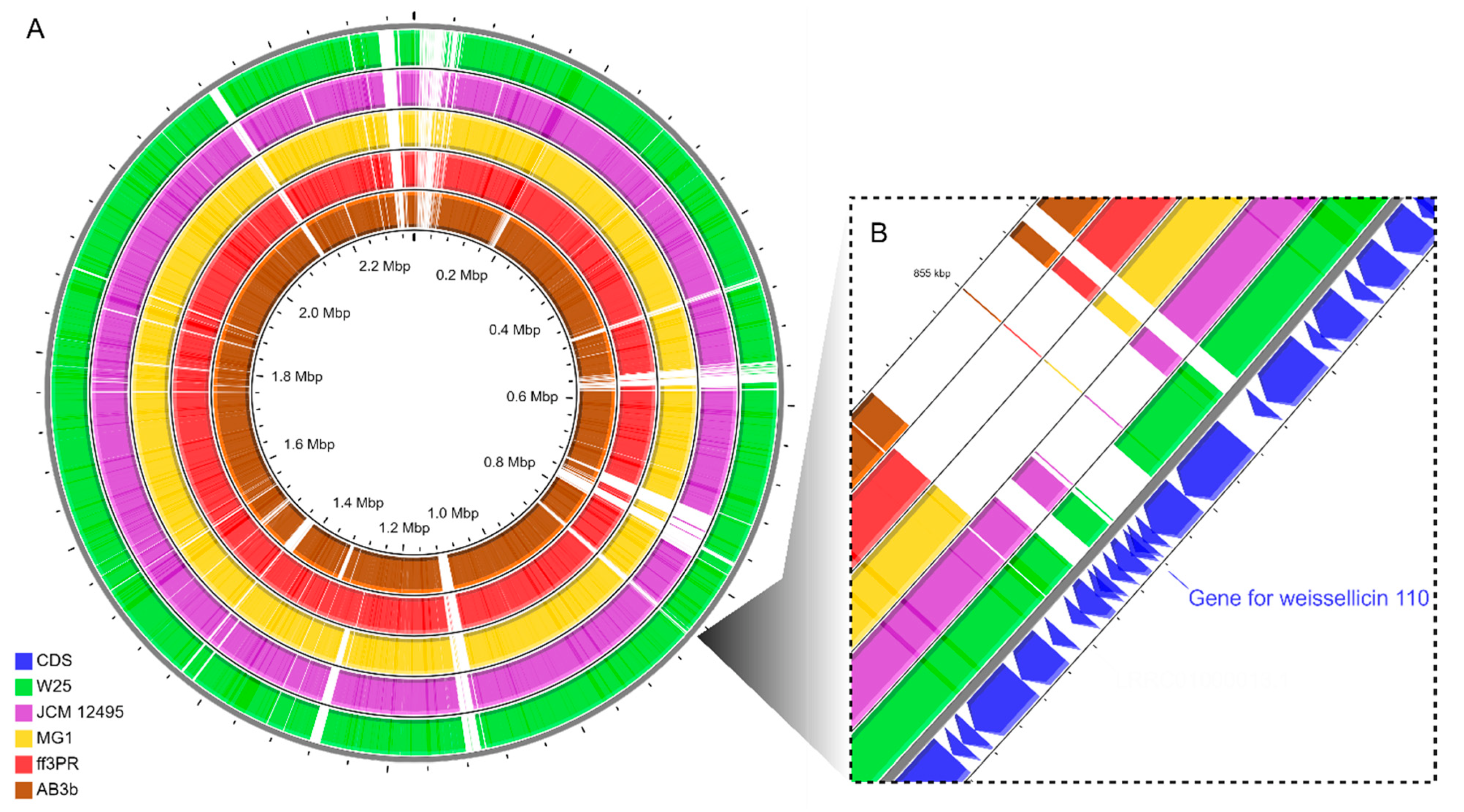
3.2. Phylogenetic Analyses and Comparative Genomic
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Finnegan, W.; Yan, M.; Holden, N.M.; Goggins, J. A review of environmental life cycle assessment studies examining cheese production. Int. J. Life Cycle Assess. 2018, 23, 1773–1787. [Google Scholar] [CrossRef]
- e Silva, E.O.O.; Nespolo, C.R.; Sehn, C.P.; Pinheiro, F.C.; Stefani, L.M. Lactic acid bacteria with antimicrobial, proteolytic and lipolytic activities isolated from ovine dairy products. Food Sci. Technol. 2020, 40, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Pyz-Łukasik, R.; Knysz, P.; Gondek, M. Hygiene Quality and Consumer Safety of Traditional Short- and Long-Ripened Cheeses from Poland. J. Food Qual. 2018, 2018, 8732412. [Google Scholar] [CrossRef] [Green Version]
- Tapia, J.A.A.; Escobar-Ramírez, C.M.; Tamplin, M.L.; Hernández-Iturriaga, M. Characterization of Bacterial Communities in Mexican Artisanal Raw Milk “Bola de Ocosingo” Cheese by High-Throughput Sequencing. Front. Microbiol. 2018, 9, 2598. [Google Scholar] [CrossRef]
- McAuliffe, O. Symposium review: Lactococcus lactis from nondairy sources: Their genetic and metabolic diversity and potential applications in cheese. J. Dairy Sci. 2018, 101, 3597–3610. [Google Scholar] [CrossRef] [Green Version]
- Collins, M.; Samelis, J.; Metaxopoulos, J.; Wallbanks, S. Taxonomic studies on some leuconostoc-like organisms from fermented sausages: Description of a new genus Weissella for the Leuconostoc paramesenteroides group of species. J. Appl. Bacteriol. 1993, 75, 595–603. [Google Scholar] [CrossRef]
- Teixeira, C.G.; Fusieger, A.; Martins, E.; de Freitas, R.; Vakarelova, M.; Nero, L.A.; de Carvalho, A.F. Biodiversity and technological features of Weissella isolates obtained from Brazilian artisanal cheese-producing regions. LWT 2021, 147, 111474. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 24 November 2021).
- Chevreux, B.; Wetter, T.; Suhai, S. Genome Sequence Assembly Using Trace Signals and Additional Sequence Information. Ger. Conf. Bioinform. 1999, 99, 45–46. [Google Scholar]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Lee, I.; Chalita, M.; Ha, S.-M.; Na, S.-I.; Yoon, S.-H.; Chun, J. ContEst16S: An algorithm that identifies contaminated prokaryotic genomes using 16S RNA gene sequences. Int. J. Syst. Evol. Microbiol. 2017, 67, 2053–2057. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.Y.; Glass, E.M.; Kubal, M.; et al. The RAST server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Van Heel, A.J.; De Jong, A.; Song, C.; Viel, J.; Kok, J.; Kuipers, O.P. BAGEL4: A user-friendly web server to thoroughly mine RiPPs and bacteriocins. Nucleic Acids Res. 2018, 46, W278–W281. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.V.; Lund, O.; Villa, L.; Møller Aarestrup, F.M.; Hasman, H. In Silico Detection and Typing of Plasmids using PlasmidFinder and Plasmid Multilocus Sequence Typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [Green Version]
- Cosentino, S.; Voldby Larsen, M.; Møller Aarestrup, F.; Lund, O. PathogenFinder—Distinguishing Friend from Foe Using Bacterial Whole Genome Sequence Data. PLoS ONE 2013, 8, e77302. [Google Scholar] [CrossRef]
- Hasman, H.; Saputra, D.; Sicheritz-Ponten, T.; Lund, O.; Svendsen, C.A.; Frimodt-Møller, N.; Aarestrup, F.M. Rapid Whole-Genome Sequencing for Detection and Characterization of Microorganisms Directly from Clinical Samples. J. Clin. Microbiol. 2014, 52, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.V.; Cosentino, S.; Lukjancenko, O.; Saputra, D.; Rasmussen, S.; Hasman, H.; Sicheritz-Pontén, T.; Aarestrup, F.M.; Ussery, D.W.; Lund, O. Benchmarking of Methods for Genomic Taxonomy. J. Clin. Microbiol. 2014, 52, 1529–1539. [Google Scholar] [CrossRef] [Green Version]
- Meier-Kolthoff, J.P.; Göker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Carbasse, J.S.; Peinado-Olarte, R.L.; Göker, M. TYGS and LPSN: A database tandem for fast and reliable genome-based classification and nomenclature of prokaryotes. Nucleic Acids Res. 2021, 50, D801–D807. [Google Scholar] [CrossRef]
- Xu, L.; Dong, Z.; Fang, L.; Luo, Y.; Wei, Z.; Guo, H.; Zhang, G.; Gu, Y.Q.; Coleman-Derr, D.; Xia, Q.; et al. OrthoVenn2: A web server for whole-genome comparison and annotation of orthologous clusters across multiple species. Nucleic Acids Res. 2019, 47, W52–W58. [Google Scholar] [CrossRef] [Green Version]
- Stothard, P.; Wishart, D.S. Circular genome visualization and exploration using CGView. Bioinformatics 2005, 21, 537–539. [Google Scholar] [CrossRef] [Green Version]
- Li, S.-W.; Chen, Y.-S.; Lee, Y.-S.; Yang, C.-H.; Srionnual, S.; Wu, H.-C.; Chang, C.-H. Comparative genomic analysis of bacteriocin-producing Weissella cibaria 110. Appl. Microbiol. Biotechnol. 2017, 101, 1227–1237. [Google Scholar] [CrossRef]
- Lynch, K.; Arendt, E.K.; Lucid, A.; Lucey, B.; Sleator, R.; Coffey, A. Genomics of Weissella cibaria with an examination of its metabolic traits. Microbiology 2015, 161, 914–930. [Google Scholar] [CrossRef]
- Bjorkroth, J.; Schillinger, U.; Geisen, R.; Weiss, N.; Hoste, B.; Holzapfel, W.H.; Korkeala, H.; Vandamme, P. Taxonomic study of Weissella confusa and description of Weissella cibaria sp. nov., detected in food and clinical samples. Int. J. Syst. Evol. Microbiol. 2002, 52, 141–148. [Google Scholar] [CrossRef]
- Lefort, V.; Desper, R.; Gascuel, O. FastME 2.0: A Comprehensive, Accurate, and Fast Distance-Based Phylogeny Inference Program: Table 1. Mol. Biol. Evol. 2015, 32, 2798–2800. [Google Scholar] [CrossRef] [Green Version]
- Farris, J.S. Estimating Phylogenetic Trees from Distancec Matrices. Am. Nat. 1972, 106, 645–668. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genome Feature | W25 | 110 | AB3b | ff3PR | JCM 12495T | MG1 |
|---|---|---|---|---|---|---|
| Accession a | JAFNKE000000000 | LRRC00000000 | JWHT00000000 | JWHV00000000 | BJEF00000000 | JWHU00000000 |
| Reference | This study | [26] | [27] | [27] | [28] | [27] |
| Contigs | 41 | 18 | 88 | 60 | 25 | 44 |
| Size (pb) | 2,412,435 | 2,347,049 | 2,465,158 | 2,357,128 | 2,323,953 | 2,436,232 |
| GC content (%) b | 45.04 | 44.9 | 44.7 | 44.9 | 45.1 | 44.7 |
| CDS | 2190 | 2209 | 2348 | 2228 | 2124 | 2284 |
| tRNA | 84 | 76 | 67 | 70 | 77 | 62 |
| rRNA | 15 | 5 | 7 | 5 | 9 | 4 |
| tmRNA | 1 | 1 | 1 | 1 | 1 | 1 |
| Repeat region | 1 | 1 | 0 | 0 | 0 | 0 |
| Strains | DDH | Prob. DDH ≥ 70% | G + C Difference |
|---|---|---|---|
| W. cibaria 110 | 63.8 | 63.95 | 0.18 |
| W. cibaria AB3b | 72.6 | 82.63 | 0.36 |
| W. cibaria ff3PR | 71.7 | 81.21 | 0.17 |
| W. cibaria JCM12495 | 87.1 | 94.69 | 0.11 |
| W. cibaria MG1 | 71.6 | 81.02 | 0.29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teixeira, C.G.; Rodrigues, R.d.S.; Yamatogi, R.S.; Lucau-Danila, A.; Drider, D.; Nero, L.A.; de Carvalho, A.F. Genomic Analyses of Weissella cibaria W25, a Potential Bacteriocin-Producing Strain Isolated from Pasture in Campos das Vertentes, Minas Gerais, Brazil. Microorganisms 2022, 10, 314. https://doi.org/10.3390/microorganisms10020314
Teixeira CG, Rodrigues RdS, Yamatogi RS, Lucau-Danila A, Drider D, Nero LA, de Carvalho AF. Genomic Analyses of Weissella cibaria W25, a Potential Bacteriocin-Producing Strain Isolated from Pasture in Campos das Vertentes, Minas Gerais, Brazil. Microorganisms. 2022; 10(2):314. https://doi.org/10.3390/microorganisms10020314
Chicago/Turabian StyleTeixeira, Camila Gonçalves, Rafaela da Silva Rodrigues, Ricardo Seiti Yamatogi, Anca Lucau-Danila, Djamel Drider, Luís Augusto Nero, and Antônio Fernandes de Carvalho. 2022. "Genomic Analyses of Weissella cibaria W25, a Potential Bacteriocin-Producing Strain Isolated from Pasture in Campos das Vertentes, Minas Gerais, Brazil" Microorganisms 10, no. 2: 314. https://doi.org/10.3390/microorganisms10020314
APA StyleTeixeira, C. G., Rodrigues, R. d. S., Yamatogi, R. S., Lucau-Danila, A., Drider, D., Nero, L. A., & de Carvalho, A. F. (2022). Genomic Analyses of Weissella cibaria W25, a Potential Bacteriocin-Producing Strain Isolated from Pasture in Campos das Vertentes, Minas Gerais, Brazil. Microorganisms, 10(2), 314. https://doi.org/10.3390/microorganisms10020314