
Lactic Acid Bacteria Isolated from Fresh Vegetable Products: Potential Probiotic and Postbiotic Characteristics Including Immunomodulatory Effects

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Isolation of Lactic Acid Bacteria
2.3. Tolerance to Stimulated Digestion Conditions Using INFOGEST2.0
2.4. Bile Tolerance
2.5. Bile Salt Hydrolase (BSH) Activity
2.6. Cholesterol Removal
2.7. Auto-Aggregation
2.8. Hydrophobicity
2.9. Adhesion to HT-29 Cells
2.10. Co-Aggregation
2.11. Antimicrobial Production
2.12. Lysozyme Activity
2.13. Antibiotic Susceptibility Test
2.14. Exopolysaccharide (EPS) Production
2.15. Identification of the LAB Isolates
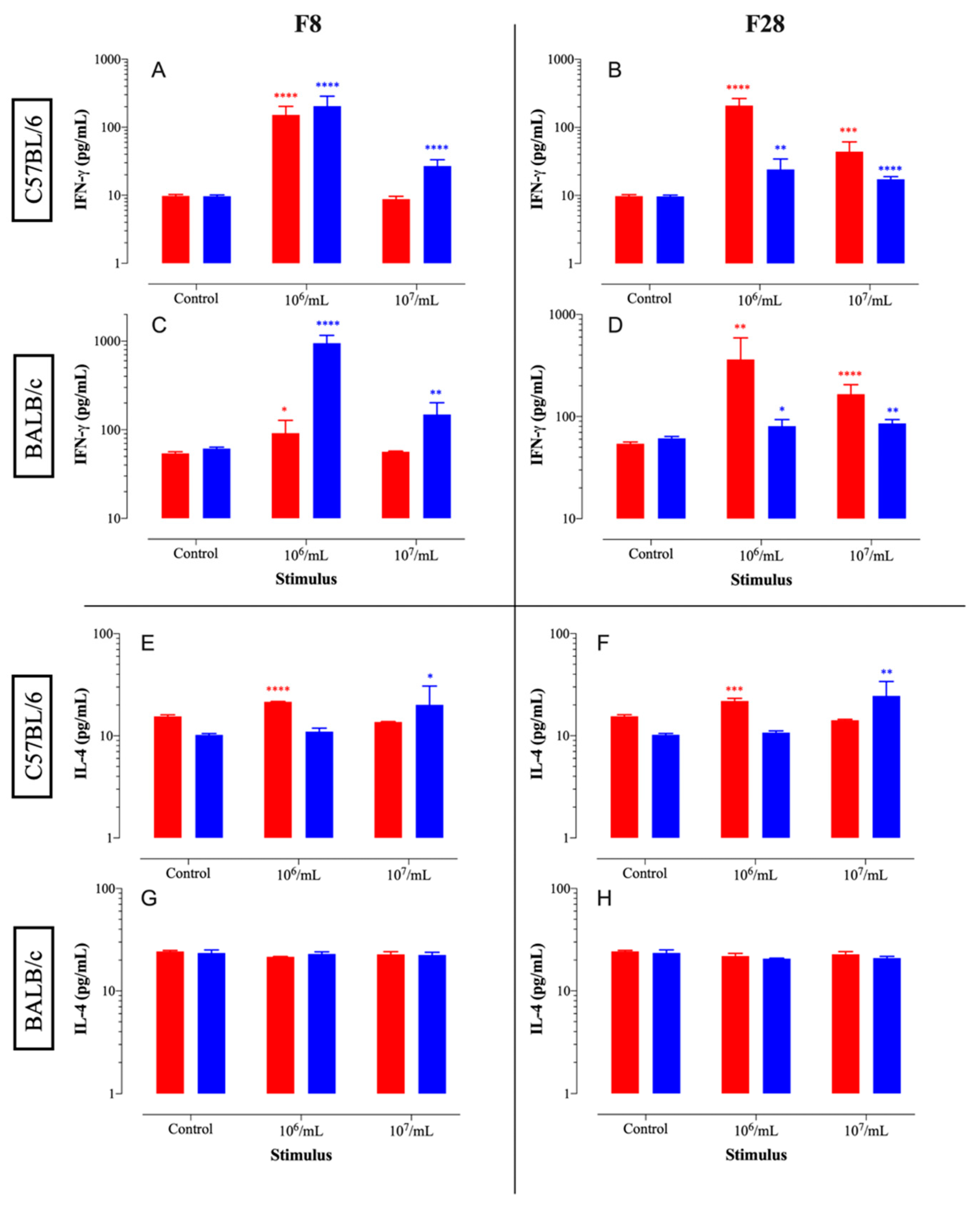
2.16. Immunomodulatory Effects
2.17. Statistical Analysis
3. Results and Discussion
3.1. Tolerance to In Vitro Digestive Condition
3.2. Bile Salts Tolerance
3.3. Bile Salt Hydrolase (BSH) Activities and Cholesterol Removal
3.4. Auto-Aggregation, Hydrophobicity, and Adherence to HT-29 Cells
3.5. Co-Aggregation and Antimicrobial Activities
3.6. Lysozyme Tolerance, Antibiotic Resistance, and EPS Production
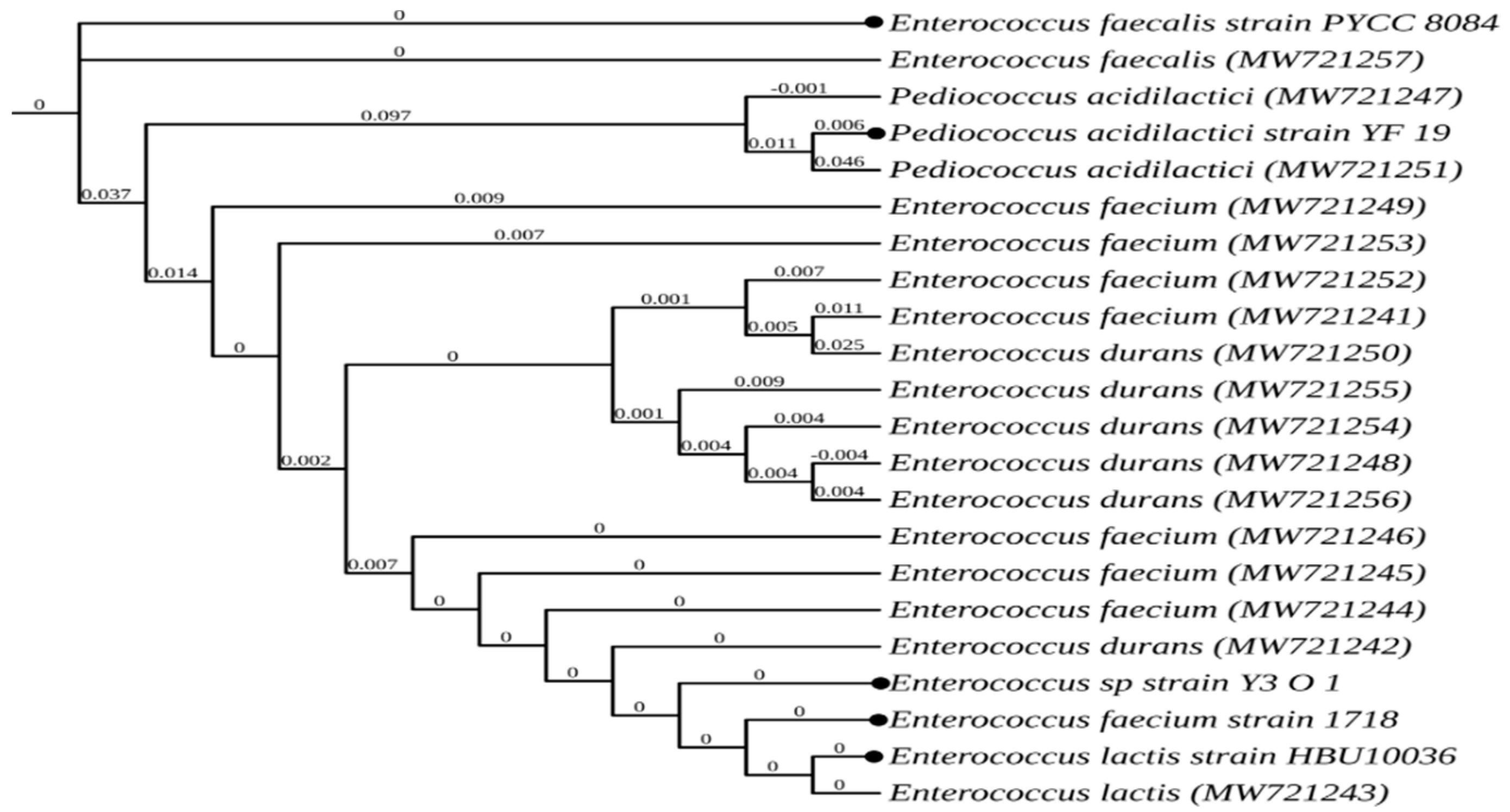
3.7. Identification by 16S rRNA
3.8. Immunomodulatory Effect
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ayyash, M.M.; Abdalla, A.K.; AlKalbani, N.S.; Baig, M.A.; Turner, M.S.; Liu, S.-Q.; Shah, N.P. Invited review: Characterization of new probiotics from dairy and nondairy products—Insights into acid tolerance, bile metabolism and tolerance, and adhesion capability. J. Dairy Sci. 2021, 104, 8363–8379. [Google Scholar] [CrossRef] [PubMed]
- Olaimat, A.N.; Aolymat, I.; Al-Holy, M.; Ayyash, M.; Abu Ghoush, M.; Al-Nabulsi, A.A.; Osaili, T.; Apostolopoulos, V.; Liu, S.-Q.; Shah, N.P. The potential application of probiotics and prebiotics for the prevention and treatment of COVID-19. Npj Sci. Food 2020, 4, 17. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mack, D.R. Probiotics in Inflammatory Bowel Diseases and Associated Conditions. Nutrients 2011, 3, 245–264. [Google Scholar] [CrossRef] [PubMed]
- Miremadi, F.; Ayyash, M.; Sherkat, F.; Stojanovska, L. Cholesterol reduction mechanisms and fatty acid composition of cellular membranes of probiotic Lactobacilli and Bifidobacteria. J. Funct. Foods 2014, 9, 295–305. [Google Scholar] [CrossRef]
- Liong, M.; Shah, N. Acid and Bile Tolerance and Cholesterol Removal Ability of Lactobacilli Strains. J. Dairy Sci. 2005, 88, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Markowiak, P.; Śliżewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef]
- Slavin, J.L.; Lloyd, B. Health Benefits of Fruits and Vegetables. Adv. Nutr. Int. Rev. J. 2012, 3, 506–516. [Google Scholar] [CrossRef] [Green Version]
- Ayyash, M.; Abushelaibi, A.; Al-Mahadin, S.; Enan, M.; El-Tarabily, K.; Shah, N. In-vitro investigation into probiotic characterisation of Streptococcus and Enterococcus isolated from camel milk. LWT-Food Sci. Technol. 2017, 87, 478–487. [Google Scholar] [CrossRef]
- Shivangi, S.; Devi, P.B.; Ragul, K.; Shetty, P.H. Probiotic Potential of Bacillus Strains Isolated from an Acidic Fermented Food Idli. Probiotics Antimicrob. Proteins 2020, 12, 1502–1513. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Sui, L.; Mu, G.; Qian, F.; Zhu, X. Screening of potential probiotics with anti-Helicobacter pylori activity from infant feces through principal component analysis. Food Biosci. 2021, 42, 101045. [Google Scholar] [CrossRef]
- Abushelaibi, A.; Al-Mahadin, S.; El-Tarabily, K.; Shah, N.P.; Ayyash, M. Characterization of potential probiotic lactic acid bacteria isolated from camel milk. LWT-Food Sci. Technol. 2017, 79, 316–325. [Google Scholar] [CrossRef]
- Oh, Y.J.; Jung, D.S. Evaluation of probiotic properties of Lactobacillus and Pediococcus strains isolated from Omegisool, a traditionally fermented millet alcoholic beverage in Korea. LWT-Food Sci. Technol. 2015, 63, 437–444. [Google Scholar] [CrossRef]
- Charteris, W.P.; Kelly, P.M.; Morelli, L.; Collins, J.K. Antibiotic Susceptibility of Potentially Probiotic Lactobacillus Species. J. Food Prot. 1998, 61, 1636–1643. [Google Scholar] [CrossRef] [PubMed]
- Alkalbani, N.; Turner, M.; Ayyash, M.M. Isolation, identification, and potential probiotic characterization of isolated lactic acid bacteria and in vitro investigation of the cytotoxicity, antioxidant, and antidiabetic activities in fermented sausage. Microb. Cell Factories 2019, 18, 188. [Google Scholar] [CrossRef] [Green Version]
- Lemoine, F.; Correia, D.; Lefort, V.; Doppelt-Azeroual, O.; Mareuil, F.; Cohen-Boulakia, S.; Gascuel, O. Ngphylogeny.fr: New generation phylogenetic services for non-specialists. Nucleic Acids Res. 2019, 47, W260–W265. [Google Scholar] [CrossRef] [Green Version]
- Al-Ramadi, B.K.; Adeghate, E.; Mustafa, N.; Ponery, A.S.; Fernandez-Cabezudo, M.J. Cytokine expression by attenuated intracellular bacteria regulates the immune response to infection: The Salmonella model. Mol. Immunol. 2002, 38, 931–940. [Google Scholar] [CrossRef]
- Fernandez-Cabezudo, M.J.; Ali, S.A.; Ullah, A.; Hasan, M.Y.; Kosanovic, M.; Fahim, M.A.; Adem, A.; Al-Ramadi, B.K. Pronounced susceptibility to infection by Salmonella enterica serovar Typhimurium in mice chronically exposed to lead correlates with a shift to Th2-type immune responses. Toxicol. Appl. Pharmacol. 2007, 218, 215–226. [Google Scholar] [CrossRef]
- Reuben, R.; Roy, P.; Sarkar, S.; Alam, A.R.U.; Jahid, I. Characterization and evaluation of lactic acid bacteria from indigenous raw milk for potential probiotic properties. J. Dairy Sci. 2020, 103, 1223–1237. [Google Scholar] [CrossRef]
- Yasmin, I.; Saeed, M.; Khan, W.A.; Khaliq, A.; Chughtai, M.F.J.; Iqbal, R.; Tehseen, S.; Naz, S.; Liaqat, A.; Mehmood, T.; et al. In Vitro Probiotic Potential and Safety Evaluation (Hemolytic, Cytotoxic Activity) of Bifidobacterium Strains Isolated from Raw Camel Milk. Microorganisms 2020, 8, 354. [Google Scholar] [CrossRef] [Green Version]
- Stasiak-Różańska, L.; Berthold-Pluta, A.; Pluta, A.; Dasiewicz, K.; Garbowska, M. Effect of Simulated Gastrointestinal Tract Conditions on Survivability of Probiotic Bacteria Present in Commercial Preparations. Int. J. Environ. Res. Public Health 2021, 18, 1108. [Google Scholar] [CrossRef]
- Yerlikaya, O.; Akbulut, N. In vitro characterisation of probiotic properties of Enterococcus faecium and Enterococcus durans strains isolated from raw milk and traditional dairy products. Int. J. Dairy Technol. 2020, 73, 98–107. [Google Scholar] [CrossRef]
- Melchior, S.; Marino, M.; Innocente, N.; Calligaris, S.; Nicoli, M.C. Effect of different biopolymer-based structured systems on the survival of probiotic strains during storage and in vitro digestion. J. Sci. Food Agric. 2020, 100, 3902–3909. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.-B.; Lew, L.-C.; Yeo, S.-K.; Parvathy, S.N.; Liong, M.-T. Probiotics and the BSH-related cholesterol lowering mechanism: A Jekyll and Hyde scenario. Crit. Rev. Biotechnol. 2013, 35, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Jawan, R.; Abbasiliasi, S.; Mustafa, S.; Kapri, M.R.; Halim, M.; Ariff, A.B. In vitro Evaluation of Potential Probiotic Strain Lactococcus lactis Gh1 and Its Bacteriocin-Like Inhibitory Substances for Potential Use in the Food Industry. Probiotics Antimicrob. Proteins 2021, 13, 422–440. [Google Scholar] [CrossRef]
- Ruiz, L.; Sánchez, B.; Margolles, A. Determination of bile salt hydrolase activity in bifidobacteria. In Bifidobacteria: Methods and Protocols; Van Sinderen, D., Ventura, M., Eds.; Springer: New York, NY, USA, 2021; pp. 149–155. [Google Scholar] [CrossRef]
- Sui, Y.; Liu, J.; Liu, Y.; Wang, Y.; Xiao, Y.; Gao, B.; Zhu, D. In vitro probiotic characterization of Lactobacillus strains from fermented tangerine vinegar and their cholesterol degradation activity. Food Biosci. 2021, 39, 100843. [Google Scholar] [CrossRef]
- Lye, H.S.; Balakrishnan, K.; Thiagarajah, K.; Ismail, N.I.M.; Ooi, S.Y. Beneficial Properties of Probiotics. Trop. Life Sci. Res. 2016, 27, 73–90. [Google Scholar] [CrossRef]
- Ladha, G.; Jeevaratnam, K. Probiotic Potential of Pediococcus pentosaceus LJR1, a Bacteriocinogenic Strain Isolated from Rumen Liquor of Goat (Capra aegagrus hircus). Food Biotechnol. 2018, 32, 60–77. [Google Scholar] [CrossRef]
- Vasiee, A.; Falah, F.; Behbahani, B.A.; Tabatabaee-Yazdi, F. Probiotic characterization of Pediococcus strains isolated from Iranian cereal-dairy fermented product: Interaction with pathogenic bacteria and the enteric cell line Caco-2. J. Biosci. Bioeng. 2020, 130, 471–479. [Google Scholar] [CrossRef]
- Domingos-Lopes, M.; Stanton, C.; Ross, R.; Silva, C. Histamine and cholesterol lowering abilities of lactic acid bacteria isolated from artisanal Pico cheese. J. Appl. Microbiol. 2020, 129, 1428. [Google Scholar] [CrossRef]
- Vila, T.; Rizk, A.M.; Sultan, A.S.; Jabra-Rizk, M.A. The power of saliva: Antimicrobial and beyond. PLoS Pathog. 2019, 15, e1008058. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Liu, X.; Zhi, A.; Li, J.; Wu, Y.; Zhang, Y. Characterization and Selection of Lactobacillus plantarum and Lactobacillus paracasei for prevention of oral bacterial infections from Chinese pickle. AMB Express 2021, 11, 84. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, A.K.; Ayyash, M.M.; Olaimat, A.N.; Osaili, T.M.; Al-Nabulsi, A.A.; Shah, N.P.; Holley, R. Exopolysaccharides as Antimicrobial Agents: Mechanism and Spectrum of Activity. Front. Microbiol. 2021, 12, 664395. [Google Scholar] [CrossRef] [PubMed]
- Cuffia, F.; George, G.; Godoy, L.; Vinderola, G.; Reinheimer, J.; Burns, P. In vivo study of the immunomodulatory capacity and the impact of probiotic strains on physicochemical and sensory characteristics: Case of pasta filata soft cheeses. Food Res. Int. 2019, 125, 108606. [Google Scholar] [CrossRef] [PubMed]
- Ai, C.; Ma, N.; Zhang, Q.; Wang, G.; Liu, X.; Tian, F.; Chen, P.; Chen, W. Immunomodulatory Effects of Different Lactic Acid Bacteria on Allergic Response and Its Relationship with in vitro Properties. PLoS ONE 2016, 11, e0164697. [Google Scholar] [CrossRef]
- Vinderola, C.G.; Duarte, J.; Thangavel, D.; Perdigón, G.; Farnworth, E.; Matar, C. Immunomodulating capacity of kefir. J. Dairy Res. 2005, 72, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Mills, C.D.; Kincaid, K.; Alt, J.M.; Heilman, M.J.; Hill, A.M. M-1/M-2 Macrophages and the Th1/Th2 Paradigm. J. Immunol. 2000, 164, 6166–6173. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Isolates | Starting | After Gastric | After Intestine |
|---|---|---|---|
| F1 | 8.8 ± 0.42 | 8.3 ± 0.67 | 8.2 ± 0.59 |
| F2 | 8.6 ± 0.26 | 8.5 ± 0.76 | 8.2 ± 0.61 |
| F3 | 8.6 ± 0.28 | 8.4 ± 0.77 | 8.1 ± 0.50 |
| F4 | 8.8 ± 0.52 | 8.5 ± 0.85 | 8.3 ± 0.70 |
| F5 | 8.9 ± 0.45 | 8.5 ± 0.85 | 8.1 ± 0.70 |
| F6 | 8.8 ± 0.37 | 8.4 ± 0.72 | 8.2 ± 0.53 |
| F7 | 8.8 ± 0.49 | 8.4 ± 0.84 | 8.3 ± 0.73 |
| F8 | 8.9 ± 0.44 | 8.4 ± 0.79 | 8.3 ± 0.65 |
| F9 | 8.9 ± 0.54 | 8.2 ± 0.90 | 8.3 ± 0.75 |
| F10 | 8.7 ± 0.44 | 8.4 ± 0.78 | 8.4 ± 0.73 |
| F11 | 8.8 ± 0.57 | 8.1 ± 0.87 | 8.3 ± 0.67 |
| F12 | 8.7 ± 0.53 | 8.5 ± 0.86 | 8.4 ± 0.69 |
| F13 | 8.7 ± 0.41 | 8.4 ± 0.63 | 8.3 ± 0.74 |
| F14 | 8.7 ± 0.43 | 8.4 ± 0.77 | 8.2 ± 0.78 |
| F15 | 8.8 ± 0.38 | 8.4 ± 0.72 | 8.5 ± 0.75 |
| F16 | 8.8 ± 0.31 | 8.4 ± 0.77 | 8.4 ± 0.68 |
| F17 | 8.8 ± 0.30 | 8.4 ± 0.69 | 8.3 ± 0.73 |
| F18 | 8.9 ± 0.46 | 8.4 ± 0.71 | 8.4 ± 0.75 |
| F19 | 8.3 ± 0.09 | 8.1 ± 0.62 | 7.9 ± 0.58 |
| F20 | 8.4 ± 0.18 | 8.3 ± 0.68 | 7.6 ± 0.57 |
| F21 | 8.9 ± 0.49 | 8.6 ± 0.83 | 8.4 ± 0.66 |
| F22 | 8.4 ± 0.32 | 8.3 ± 0.61 | 8.3 ± 0.64 |
| F23 | 8.6 ± 0.31 | 8.5 ± 0.88 | 8.2 ± 0.61 |
| F24 | 8.8 ± 0.46 | 8.4 ± 0.77 | 7.9 ± 0.35 |
| F25 | 8.7 ± 0.39 | 8.3 ± 0.78 | 8.2 ± 0.59 |
| F26 | 8.8 ± 0.53 | 8.5 ± 0.87 | 8.1 ± 0.47 |
| F27 | 8.4 ± 0.68 | 8.4 ± 0.85 | 8.1 ± 0.50 |
| F28 | 9.0 ± 0.57 | 8.5 ± 0.93 | 8.4 ± 0.81 |
| F29 | 8.8 ± 0.48 | 8.4 ± 0.90 | 7.6 ± 0.38 |
| F30 | 8.8 ± 0.51 | 8.6 ± 1.00 | 8.1 ± 0.51 |
| F31 | 8.7 ± 0.54 | 8.4 ± 0.78 | 8.2 ± 0.50 |
| F32 | 8.9 ± 0.61 | 8.2 ± 0.63 | 7.9 ± 0.44 |
| F33 | 8.6 ± 0.57 | 8.4 ± 0.74 | 7.7 ± 0.46 |
| F34 | 8.3 ± 0.48 | 8.0 ± 0.69 | 8.0 ± 0.40 |
| F35 | 8.6 ± 0.50 | 8.2 ± 0.59 | 7.9 ± 0.37 |
| F36 | 8.8 ± 0.43 | 8.4 ± 0.68 | 7.7 ± 0.30 |
| F37 | 8.9 ± 0.52 | 8.4 ± 0.84 | 8.2 ± 0.50 |
| F38 | 8.7 ± 0.40 | 8.3 ± 0.66 | 7.4 ± 0.20 |
| F39 | 8.8 ± 0.54 | 8.2 ± 0.71 | 8.1 ± 0.56 |
| F40 | 8.8 ± 0.58 | 8.5 ± 0.81 | 8.2 ± 0.52 |
| F41 | 8.9 ± 0.57 | 8.4 ± 0.72 | 8.5 ± 0.85 |
| F42 | 8.8 ± 0.51 | 8.5 ± 0.93 | 8.1 ± 0.53 |
| F43 | 8.9 ± 0.78 | 8.5 ± 0.79 | 8.5 ± 0.74 |
| F44 | 8.8 ± 0.52 | 8.5 ± 0.87 | 8.1 ± 0.53 |
| F45 | 9.0 ± 0.52 | 8.4 ± 0.72 | 8.4 ± 0.62 |
| F46 | 8.6 ± 0.38 | 8.4 ± 0.71 | 8.2 ± 0.49 |
| Isolates | 3 h | 6 h | ||||
|---|---|---|---|---|---|---|
| CA | OX | TA | CA | OX | TA | |
| F1 | 43.9 | 42.7 | 56.1 | 39.3 | 39.9 | 58.8 |
| F5 | 29.1 | 38.9 | 48.7 | 27.6 | 52.4 | 63.0 |
| F8 | 53.7 | 40.2 | 60.9 | 51.1 | 39.0 | 60.7 |
| F13 | 46.2 | 37.4 | 59.5 | 45.3 | 33.6 | 60.0 |
| F15 | 45.0 | 39.9 | 59.0 | 41.5 | 35.2 | 57.0 |
| F18 | 41.4 | 40.7 | 60.0 | 36.4 | 38.8 | 58.1 |
| F21 | 22.5 | 39.7 | 49.8 | 20.5 | 51.1 | 63.8 |
| F23 | 26.7 | 33.5 | 44.9 | 23.9 | 49.9 | 59.6 |
| F25 | 32.3 | 40.7 | 55.3 | 35.4 | 55.7 | 66.9 |
| F26 | 33.0 | 42.5 | 41.8 | 29.7 | 60.6 | 56.0 |
| F28 | 26.3 | 42.1 | 51.2 | 17.8 | 56.0 | 63.5 |
| F31 | 28.0 | 35.6 | 47.5 | 25.2 | 52.4 | 64.0 |
| F37 | 33.0 | 32.5 | 45.4 | 37.3 | 55.8 | 62.7 |
| F40 | 36.1 | 50.1 | 48.3 | 34.5 | 62.2 | 62.2 |
| F41 | 35.3 | 40.7 | 45.5 | 29.3 | 42.9 | 63.6 |
| F43 | 32.3 | 45.1 | 56.3 | 37.8 | 50.4 | 72.5 |
| F46 | 37.6 | 51.5 | 54.8 | 44.4 | 63.9 | 68.6 |
| Isolates | BSH (U/mg) | CR (%) |
|---|---|---|
| F1 | 0.83 ± 0.03 f | 35.0 ± 1.41 a |
| F5 | 0.95 ± 0.05 b | 27.5 ± 2.12 e |
| F8 | 1.03 ± 0.07 a | 34.5 ± 0.71 a |
| F13 | 1.00 ± 0.08 a | 17.5 ± 0.71 g |
| F15 | 0.93 ± 0.05 b | 17.0 ± 1.41 g |
| F18 | 0.83 ± 0.03 f | 29.5 ± 0.71 d |
| F21 | 0.91 ± 0.06 c | 30.0 ± 0.23 c |
| F23 | 0.87 ± 0.07 d | 33.0 ± 1.41 b |
| F25 | 0.80 ± 0.02 g | 30.5 ± 3.54 c |
| F26 | 0.84 ± 0.04 e | 24.5 ± 2.12 f |
| F28 | 0.84 ± 0.04 e | 29.0 ± 0.99 d |
| F31 | 0.84 ± 0.05 e | 30.0 ± 2.83 c |
| F37 | 0.82 ± 0.04 f | 35.0 ± 0.98 a |
| F40 | 0.80 ± 0.03 g | 30.0 ± 2.83 c |
| F41 | 0.81 ± 0.06 g | 27.5 ± 2.12 e |
| F43 | 0.97 ± 0.04 a,b | 24.5 ± 2.12 f |
| F46 | 0.91 ± 0.07 c | 30.0 ± 2.83 c |
| Isolates | Auto-Aggregation (%) | Hydrophobicity (%) | Attachment to HT-29 Cells | |||
|---|---|---|---|---|---|---|
| 4 h | 24 h | Xylene | Octane | Hexadecane | Log10 CFU | |
| F1 | 26.1 ± 0.87 a | 66.5 ± 2.83 c | 61.4 ± 3.07 b | 68.7 ± 2.06 c | 76.1 ± 3.05 b | 8.0 ± 0.08 c |
| F5 | 9.8 ± 2.27 d | 63.6 ± 2.57 d | 35.9 ± 1.79 c,d | 40.7 ± 1.22 d,e | 47.3 ± 1.89 c | 8.0 ± 0.00 c |
| F8 | 15.6 ± 1.90 c | 45.0 ± 2.19 f | 77.1 ± 3.86 a | 79.0 ± 2.37 b | 84.3 ± 3.37 a | 8.1 ± 0.02 b |
| F13 | 26.2 ± 0.02 a | 56.7 ± 0.13 e | 71.0 ± 3.55 a | 86.7 ± 2.60 a | 82.0 ± 3.28 a,b | 8.0 ± 0.10 c |
| F15 | 18.9 ± 0.31 b | 47.2 ± 0.60 f | 66.0 ± 3.30 b | 79.0 ± 2.37 b | 80.7 ± 3.23 b | 8.1 ± 0.07 b |
| F18 | 17.4 ± 0.58 b | 42.9 ± 0.29 g | 56.9 ± 2.84 b,c | 69.4 ± 2.08 c | 73.3 ± 2.93 b,c | 8.1 ± 0.04 b |
| F21 | 2.0 ± 0.16 h | 70.3 ± 0.84 b | 13.3 ± 0.66 f | 36.6 ± 1.10 d,e | 39.6 ± 1.58 d | 8.1 ± 0.04 b |
| F23 | 3.8 ± 1.15 f | 73.2 ± 2.90 a | 32.7 ± 1.64 d | 38.3 ± 1.15 d,e | 45.7 ± 1.83 c | 8.1 ± 0.03 b |
| F25 | 2.3 ± 1.06 g | 63.6 ± 1.44 d | 13.7 ± 0.69 f | 28.6 ± 0.86 e | 32.1 ± 1.29 e | 8.1 ± 0.04 b |
| F26 | 3.6 ± 0.30 f | 70.6 ± 2.61 b | 13.0 ± 0.65 f | 17.3 ± 0.52 g | 29.3 ± 1.17 f | 7.9 ± 0.03 d |
| F28 | 1.8 ± 0.13 h | 70.5 ± 1.99 b | 11.4 ± 0.57 g | 29.9 ± 0.90 e | 34.9 ± 1.39 d,e | 8.0 ± 0.07 c |
| F31 | 2.4 ± 0.16 g | 72.0 ± 1.17 a | 18.9 ± 0.94 e | 23.7 ± 0.71 e,f | 42.3 ± 1.69 c | 8.1 ± 0.06 b |
| F37 | 8.2 ± 0.34 d | 70.5 ± 2.73 b | 33.4 ± 1.67 c,d | 21.7 ± 0.65 f | 34.1 ± 1.37 d,e | 8.0 ± 0.05 c |
| F40 | 8.0 ± 0.53 e | 63.4 ± 1.02 d | 6.9 ± 0.34 h | 30.3 ± 0.91 e | 35.3 ± 1.41 d,e | 7.5 ± 0.01 d |
| F41 | 11.1 ± 1.04 c | 35.2 ± 0.35 h | 34.0 ± 1.70 c,d | 44.6 ± 1.34 d | 30.7 ± 1.23 e | 8.0 ± 0.05 c |
| F43 | 9.4 ± 1.91 d | 42.4 ± 1.61 g | 40.1 ± 2.01 c | 56.1 ± 1.68 c,d | 46.9 ± 1.87 c | 8.1 ± 0.02 b |
| F46 | - | - | 31.0 ± 1.55 d | 45.7 ± 1.37 d | 39.9 ± 1.59 d | 8.3 ± 0.08 a |
| Isolates | 2 h | 24 h | ||||||
|---|---|---|---|---|---|---|---|---|
| E. coli | S. typhi | S. aureus | L. monocytogenes | E. coli | S. typhi | S. aureus | L. monocytogenes | |
| F1 | 10.1 ± 0.50 a | 10.6 ± 0.63 a | 10.0 ± 0.70 a | 10.2 ± 0.71 a | 25.5 ± 1.78 d | 32.7 ± 1.31 b | 34.8 ± 0.70 d | 32.6 ± 1.63 b |
| F5 | 7.6 ± 0.38 c | 8.4 ± 0.51 b,c | 9.7 ± 0.68 b | 8.0 ± 0.56 b | 23.8 ± 1.67 d | 24.4 ± 0.98 c | 27.1 ± 0.54 e | 25.0 ± 1.25 d,e |
| F8 | 9.4 ± 0.47 a,b | 6.7 ± 0.40 d | 10.0 ± 0.70 a | 6.8 ± 0.47 d | 21.7 ± 1.52 e | 23.8 ± 0.95 c,d | 27.4 ± 0.55 e | 23.9 ± 1.19 d,e |
| F13 | 6.8 ± 0.34 e,d | 9.6 ± 0.58 b | 9.3 ± 0.65 b | 10.3 ± 0.72a | 22.1 ± 1.55 e | 28.0 ± 1.12 b,c | 31.3 ± 0.63 d | 28.6 ± 1.43 c |
| F15 | 8.4 ± 0.42 b | 9.4 ± 0.57 b | 10.8 ± 0.76 a | 8.7 ± 0.61 b | 22.4 ± 1.57 e | 26.7 ± 1.07 c | 30.7 ± 0.61 d | 26.2 ± 1.31 d |
| F18 | 7.8 ± 0.39 c | 5.9 ± 0.35 e | 8.2 ± 0.58 c | 6.7 ± 0.47 d | 20.5 ± 1.44 e | 22.5 ± 0.90 d | 29.7 ± 0.59 e | 24.8 ± 1.24d e |
| F21 | 5.5 ± 0.27 d | 3.8 ± 0.23 g | 4.4 ± 0.31 d | 3.9 ± 0.27 f | 38.6 ± 2.70 b | 40.0 ± 1.60 | 38.6 ± 0.77 b | 35.6 ± 1.78 b |
| F23 | 4.0 ± 0.20 e | 6.7 ± 0.40 d | 6.3 ± 0.44 de | 6.1 ± 0.43 d | 43.8 ± 3.07 a | 40.8 ± 1.63 b | 39.2 ± 0.78 b | 28.0 ± 1.40 c |
| F25 | 4.1 ± 0.21 e | 5.5 ± 0.33 e | 5.1 ± 0.36 e | 3.8 ± 0.27 f | 33.7 ± 2.36 c | 42.3 ± 1.69 a | 44.0 ± 0.88 a | 36.6 ± 1.83 a,b |
| F26 | 4.9 ± 0.25 e | 7.7 ± 0.46 c | 4.9 ± 0.34 f | 6.2 ± 0.43 d | 43.5 ± 3.05 a | 39.1 ± 1.56 a,b | 39.4 ± 0.79 b | 38.7 ± 1.94 a |
| F28 | 3.2 ± 0.16 f | 4.4 ± 0.26 f | 3.2 ± 0.23 g | 5.1 ± 0.36 e | 42.4 ± 2.97 a | 33.8 ± 1.35 b | 37.4 ± 0.75 c | 31.8 ± 1.59 b |
| F31 | 2.9 ± 0.15 g | 5.5 ± 0.33 e | 4.7 ± 0.33 g | 3.9 ± 0.27 f | 45.5 ± 3.18 a | 40.7 ± 1.63 a,b | 40.7 ± 0.81 b | 36.7 ± 1.83 a,b |
| F37 | 3.4 ± 0.17 f | 6.1 ± 0.37 d | 9.1 ± 0.64 b | 5.7 ± 0.40 e | 30.5 ± 2.14 c | 24.2 ± 0.97 c | 27.4 ± 0.55 | 26.4 ± 1.32 d |
| F40 | 5.7 ± 0.28 d | 7.7 ± 0.46 c | 7.1 ± 0.50 c,d | 6.7 ± 0.47 d | 26.6 ± 1.86 d | 25.7 ± 1.03 c | 28.5 ± 0.57 e | 26.5 ± 1.32 d |
| F41 | 5.7 ± 0.28 d | 6.4 ± 0.38 d | 7.5 ± 0.53 c,d | 7.1 ± 0.49 c | 18.2 ± 1.28 f | 21.0 ± 0.84 d | 25.2 ± 0.50 f | 21.2 ± 1.06 e |
| F43 | 5.1 ± 0.26 de | 5.2 ± 0.31 e | 7.0 ± 0.49 d | 6.1 ± 0.43 d | 21.5 ± 1.50 e | 22.7 ± 0.91 d | 27.8 ± 0.56 e | 21.1 ± 1.05 e |
| F46 |
| Isolate | Probiotic a | Postbiotic b | ||||||
|---|---|---|---|---|---|---|---|---|
| E. coli | S. aureus | S. typhi | L. monocytogenes | E. coli | S. aureus | S. typhi | L. monocytogenes | |
| F1 | + | ++ | +++ | +++ | +++ | +++ | +++ | +++ |
| F5 | + | ++ | ++ | +++ | ++ | ++ | ++ | +++ |
| F8 | + | ++ | ++ | ++ | ++ | ++ | ++ | ++ |
| F13 | + | +++ | +++ | +++ | ++ | ++ | ++ | ++ |
| F15 | + | +++ | +++ | +++ | + | ++ | ++ | +++ |
| F18 | + | ++ | +++ | +++ | ++ | ++ | ++ | ++ |
| F21 | + | +++ | +++ | +++ | ++ | ++ | ++ | ++ |
| F23 | + | +++ | +++ | ++ | ++ | + | ++ | +++ |
| F25 | + | ++ | ++ | ++ | ++ | +++ | ++ | ++ |
| F26 | + | ++ | ++ | ++ | ++ | ++ | ++ | +++ |
| F28 | + | ++ | ++ | +++ | ++ | ++ | ++ | ++ |
| F31 | + | +++ | ++ | ++ | ++ | ++ | ++ | +++ |
| F37 | + | ++ | ++ | ++ | + | ++ | ++ | ++ |
| F40 | + | + | + | + | + | + | + | + |
| F41 | + | ++ | + | + | + | + | + | + |
| F43 | + | + | ++ | + | + | + | + | + |
| F46 | + | ++ | ++ | + | + | + | + | + |
| Isolate | Lysozyme Tolerance | Antibiotic Susceptibility | EPS Production | ||||
|---|---|---|---|---|---|---|---|
| 0 min | 90 min | Vancomycin | Erythromycin | Penicillin | Clindamycin | ||
| F1 | 8.1 ± 0.47 c | 8.3 ± 0.27 b,c | R | S | S | S | - |
| F5 | 8.1 ± 0.45 c | 8.1 ± 0.04 d | R | S | S | S | - |
| F8 | 8.8 ± 0.57 a | 8.4 ± 0.49 b | R | R | S | R | + |
| F13 | 8.5 ± 0.49 b | 8.2 ± 0.25 c | R | MS | S | R | + |
| F15 | 8.5 ± 0.31 b | 8.3 ± 0.34 b,c | R | S | S | MS | + |
| F18 | 8.1 ± 0.35 c | 8.2 ± 0.32 c | R | S | S | R | + |
| F21 | 8.0 ± 0.43 c | 8.2 ± 0.27 c | R | S | S | S | - |
| F23 | 8.8 ± 0.36 a | 8.2 ± 0.25 c | R | S | S | S | - |
| F25 | 8.1 ± 0.59 c | 8.3 ± 0.25 b,c | R | S | S | S | - |
| F26 | 8.8 ± 0.40 a | 8.4 ± 0.32 b | R | S | S | S | - |
| F28 | 8.0 ± 0.52 c | 8.5 ± 0.47 a | R | S | S | S | + |
| F31 | 8.6 ± 0.27 a,b | 8.2 ± 0.23 c | R | MS | S | S | + |
| F37 | 8.4 ± 0.33 b,c | 8.3 ± 0.29 b,c | R | MS | MS | MS | + |
| F40 | 8.4 ± 0.32 b,c | 8.4 ± 0.42 b | R | S | S | S | + |
| F41 | 8.4 ± 0.33 b,c | 8.4 ± 0.31 b | R | S | S | MS | + |
| F43 | 8.5 ± 0.46 b | 8.3 ± 0.35 b,c | R | S | S | S | - |
| F46 | 8.4 ± 0.37 b,c | 8.6 ± 0.45 a | R | S | S | S | - |
| Sample | Organism | Accession No |
|---|---|---|
| F1 | Enterococcus faecium | MW721241 |
| F5 | Enterococcus durans | MW721242 |
| F8 | Enterococcus lactis | MW721243 |
| F13 | Enterococcus faecium | MW721244 |
| F15 | Enterococcus faecium | MW721245 |
| F18 | Enterococcus faecium | MW721246 |
| F21 | Pediococcus acidilactici | MW721247 |
| F23 | Enterococcus durans | MW721248 |
| F25 | Enterococcus faecium | MW721249 |
| F26 | Enterococcus durans | MW721250 |
| F28 | Pediococcus acidilactici | MW721251 |
| F31 | Enterococcus faecium | MW721252 |
| F37 | Enterococcus faecium | MW721253 |
| F40 | Enterococcus durans | MW721254 |
| F41 | Enterococcus durans | MW721255 |
| F43 | Enterococcus durans | MW721256 |
| F46 | Enterococcus faecalis | MW721257 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alameri, F.; Tarique, M.; Osaili, T.; Obaid, R.; Abdalla, A.; Masad, R.; Al-Sbiei, A.; Fernandez-Cabezudo, M.; Liu, S.-Q.; Al-Ramadi, B.; et al. Lactic Acid Bacteria Isolated from Fresh Vegetable Products: Potential Probiotic and Postbiotic Characteristics Including Immunomodulatory Effects. Microorganisms 2022, 10, 389. https://doi.org/10.3390/microorganisms10020389
Alameri F, Tarique M, Osaili T, Obaid R, Abdalla A, Masad R, Al-Sbiei A, Fernandez-Cabezudo M, Liu S-Q, Al-Ramadi B, et al. Lactic Acid Bacteria Isolated from Fresh Vegetable Products: Potential Probiotic and Postbiotic Characteristics Including Immunomodulatory Effects. Microorganisms. 2022; 10(2):389. https://doi.org/10.3390/microorganisms10020389
Chicago/Turabian StyleAlameri, Fatima, Mohammad Tarique, Tareq Osaili, Riyad Obaid, Abdelmoneim Abdalla, Razan Masad, Ashraf Al-Sbiei, Maria Fernandez-Cabezudo, Shao-Quan Liu, Basel Al-Ramadi, and et al. 2022. "Lactic Acid Bacteria Isolated from Fresh Vegetable Products: Potential Probiotic and Postbiotic Characteristics Including Immunomodulatory Effects" Microorganisms 10, no. 2: 389. https://doi.org/10.3390/microorganisms10020389
APA StyleAlameri, F., Tarique, M., Osaili, T., Obaid, R., Abdalla, A., Masad, R., Al-Sbiei, A., Fernandez-Cabezudo, M., Liu, S. -Q., Al-Ramadi, B., & Ayyash, M. (2022). Lactic Acid Bacteria Isolated from Fresh Vegetable Products: Potential Probiotic and Postbiotic Characteristics Including Immunomodulatory Effects. Microorganisms, 10(2), 389. https://doi.org/10.3390/microorganisms10020389