
Trends in Microbial Community Composition and Function by Soil Depth


{kind=link}
Abstract
:1. Introduction
2. Soil Biogeochemical Properties and Depth
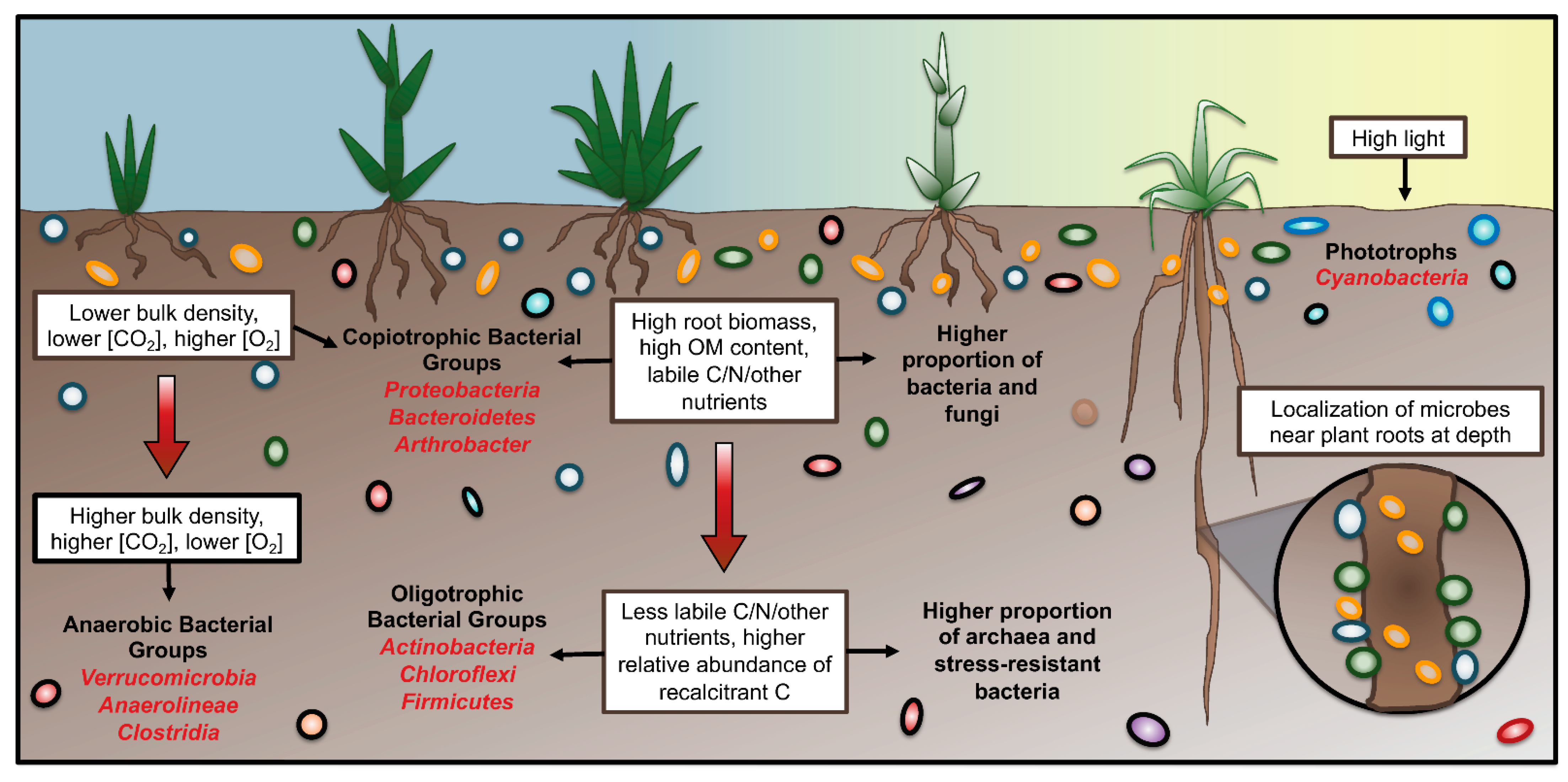
2.1. Soil Biogeochemical Properties
2.2. Soil Biogeochemical Properties and the Microbiome
2.3. Soil Physical Structure
3. Changes in Microbial Community Composition with Depth
3.1. Changes in Biomass and Diversity
3.2. Changes in Bacterial Composition
3.2.1. Changes in Gram-Positive and Gram-Negative Bacteria
3.2.2. Changes in Bacterial Phyla
3.2.3. Changes in Bacterial Classes
3.3. Changes in Archaeal Phyla
3.4. Changes in Fungal Phyla
3.5. Plant Presence and Soil Microbiome Depth Trends
3.6. Changes in Microbial Network Patterns
4. Changes in Microbial Activity with Depth
4.1. Changes in Functional Guilds
4.1.1. Anaerobic Guilds
4.1.2. Starvation Responses
5. Microbial Nutrient Cycling and Soil Depth
5.1. Carbon Cycling
5.1.1. Carbon Availability by Depth
5.1.2. Carbon Kinetics by Depth
5.1.3. Changes in Carbon Metabolism Patterns by Depth
5.2. Nitrogen Cycling
5.3. Positive Priming
6. Conclusions and Future Directions for Modeling Approaches
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lomander, A.; Kätterer, T.; Andrén, O. Carbon Dioxide Evolution from Top- and Subsoil as Affected by Moisture and Constant and Fluctuating Temperature. Soil Biol. Biochem. 1998, 30, 2017–2022. [Google Scholar] [CrossRef]
- Jones, D.L.; Magthab, E.A.; Gleeson, D.B.; Hill, P.W.; Sánchez-Rodríguez, A.R.; Roberts, P.; Ge, T.; Murphy, D.V. Microbial Competition for Nitrogen and Carbon Is as Intense in the Subsoil as in the Topsoil. Soil Biol. Biochem. 2018, 117, 72–82. [Google Scholar] [CrossRef]
- Meyer, N.; Welp, G.; Rodionov, A.; Borchard, N.; Martius, C.; Amelung, W. Nitrogen and Phosphorus Supply Controls Soil Organic Carbon Mineralization in Tropical Topsoil and Subsoil. Soil Biol. Biochem. 2018, 119, 152–161. [Google Scholar] [CrossRef]
- Rumpel, C.; Chabbi, A.; Marschner, B. Carbon Storage and Sequestration in Subsoil Horizons: Knowledge, Gaps and Potentials. In Recarbonization of the Biosphere; Lal, R., Lorenz, K., Hüttl, R.F., Schneider, B.U., von Braun, J., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 445–464. ISBN 978-94-007-4158-4. [Google Scholar]
- Fierer, N.; Schimel, J.P.; Holden, P.A. Variations in Microbial Community Composition through Two Soil Depth Profiles. Soil Biol. Biochem. 2003, 35, 167–176. [Google Scholar] [CrossRef]
- Angst, G.; Messinger, J.; Greiner, M.; Häusler, W.; Hertel, D.; Kirfel, K.; Kögel-Knabner, I.; Leuschner, C.; Rethemeyer, J.; Mueller, C.W. Soil Organic Carbon Stocks in Topsoil and Subsoil Controlled by Parent Material, Carbon Input in the Rhizosphere, and Microbial-Derived Compounds. Soil Biol. Biochem. 2018, 122, 19–30. [Google Scholar] [CrossRef]
- Salome, C.; Nunan, N.; Pouteau, V.; Lerch, T.Z.; Chenu, C. Carbon Dynamics in Topsoil and in Subsoil May Be Controlled by Different Regulatory Mechanisms: Carbon Dynamics in Topsoil and in Subsoil. Glob. Change Biol. 2010, 16, 416–426. [Google Scholar] [CrossRef]
- Yao, X.; Zhang, N.; Zeng, H.; Wang, W. Effects of Soil Depth and Plant–Soil Interaction on Microbial Community in Temperate Grasslands of Northern China. Sci. Total Environ. 2018, 630, 96–102. [Google Scholar] [CrossRef]
- Jobbágy, E.G.; Jackson, R.B. The Distribution of Soil Nutrients with Depth: Global Patterns and the Imprint of Plants. Biogeochemistry 2001, 53, 51–77. [Google Scholar] [CrossRef]
- Kaiser, J.-P.; Bollag, J.-M. Microbial Activity in the Terrestrial Subsurface. Experientia 1990, 46, 797–806. [Google Scholar] [CrossRef]
- Iqbal, J.; Thomasson, J.A.; Jenkins, J.N.; Owens, P.R.; Whisler, F.D. Spatial Variability Analysis of Soil Physical Properties of Alluvial Soils. Soil Sci. Soc. Am. J. 2005, 69, 1338–1350. [Google Scholar] [CrossRef]
- Keshavarzi, A.; Tuffour, H.O.; Bagherzadeh, A.; Vasu, D. Spatial and Fractal Characterization of Soil Properties across Soil Depth in an Agricultural Field, Northeast Iran. Eurasian J. Soil Sci. 2018, 7, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Allison, V.J.; Yermakov, Z.; Miller, R.M.; Jastrow, J.D.; Matamala, R. Using Landscape and Depth Gradients to Decouple the Impact of Correlated Environmental Variables on Soil Microbial Community Composition. Soil Biol. Biochem. 2007, 39, 505–516. [Google Scholar] [CrossRef]
- Brewer, T.E.; Aronson, E.L.; Arogyaswamy, K.; Billings, S.A.; Botthoff, J.K.; Campbell, A.N.; Dove, N.C.; Fairbanks, D.; Gallery, R.E.; Hart, S.C.; et al. Ecological and Genomic Attributes of Novel Bacterial Taxa That Thrive in Subsurface Soil Horizons. mBio 2019, 10, e01318-19. [Google Scholar] [CrossRef] [Green Version]
- Eilers, K.G.; Debenport, S.; Anderson, S.; Fierer, N. Digging Deeper to Find Unique Microbial Communities: The Strong Effect of Depth on the Structure of Bacterial and Archaeal Communities in Soil. Soil Biol. Biochem. 2012, 50, 58–65. [Google Scholar] [CrossRef]
- Bai, R.; Wang, J.-T.; Deng, Y.; He, J.-Z.; Feng, K.; Zhang, L.-M. Microbial Community and Functional Structure Significantly Varied among Distinct Types of Paddy Soils but Responded Differently along Gradients of Soil Depth Layers. Front. Microbiol. 2017, 8, 945. [Google Scholar] [CrossRef]
- van Leeuwen, J.P.; Djukic, I.; Bloem, J.; Lehtinen, T.; Hemerik, L.; de Ruiter, P.C.; Lair, G.J. Effects of Land Use on Soil Microbial Biomass, Activity and Community Structure at Different Soil Depths in the Danube Floodplain. Eur. J. Soil Biol. 2017, 79, 14–20. [Google Scholar] [CrossRef]
- Blume, E.; Bischoff, M.; Reichert, J.M.; Moorman, T.; Konopka, A.; Turco, R.F. Surface and Subsurface Microbial Biomass, Community Structure and Metabolic Activity as a Function of Soil Depth and Season. Appl. Soil Ecol. 2002, 20, 171–181. [Google Scholar] [CrossRef]
- Tripathi, B.M.; Kim1, H.M.; Jung, J.Y.; Nam, S.; Ju, H.T.; Kim, M.; Lee, Y.K. Distinct Taxonomic and Functional Profiles of the Microbiome Associated with Different Soil Horizons of a Moist Tussock Tundra in Alaska. Front. Microbiol. 2019, 10, 1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falkowski, P.G.; Fenchel, T.; Delong, E.F. The Microbial Engines That Drive Earth’s Biogeochemical Cycles. Science 2008, 320, 1034–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahu, N.; Vasu, D.; Sahu, A.; Lal, N.; Singh, S.K. Strength of Microbes in Nutrient Cycling: A Key to Soil Health. In Agriculturally Important Microbes for Sustainable Agriculture; Meena, V.S., Mishra, P.K., Bisht, J.K., Pattanayak, A., Eds.; Springer: Singapore, 2017; pp. 69–86. ISBN 978-981-10-5588-1. [Google Scholar]
- Jiao, S.; Chen, W.; Wang, J.; Du, N.; Li, Q.; Wei, G. Soil Microbiomes with Distinct Assemblies through Vertical Soil Profiles Drive the Cycling of Multiple Nutrients in Reforested Ecosystems. Microbiome 2018, 6, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkinson, D.S.; Poulton, P.R.; Bryant, C. The Turnover of Organic Carbon in Subsoils. Part 1. Natural and Bomb Radiocarbon in Soil Profiles from the Rothamsted Long-Term Field Experiments. Eur. J. Soil. Sci. 2008, 59, 391–399. [Google Scholar] [CrossRef]
- Spohn, M.; Klaus, K.; Wanek, W.; Richter, A. Microbial Carbon Use Efficiency and Biomass Turnover Times Depending on Soil Depth—Implications for Carbon Cycling. Soil Biol. Biochem. 2016, 96, 74–81. [Google Scholar] [CrossRef]
- Gougoulias, C.; Clark, J.M.; Shaw, L.J. The Role of Soil Microbes in the Global Carbon Cycle: Tracking the below-Ground Microbial Processing of Plant-Derived Carbon for Manipulating Carbon Dynamics in Agricultural Systems: Role of Soil Microbes in Global Carbon Cycle: Carbon Tracking & Agro-Cosystem Management. J. Sci. Food Agric. 2014, 94, 2362–2371. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Huang, Y.; An, S.; Sun, H.; Bhople, P.; Chen, Z. Soil Physicochemical and Microbial Characteristics of Contrasting Land-Use Types along Soil Depth Gradients. Catena 2018, 162, 345–353. [Google Scholar] [CrossRef]
- Richter, D.D.; Markewitz, D. How Deep Is Soil? BioScience 1995, 45, 600–609. [Google Scholar] [CrossRef]
- Seuradge, B.J.; Oelbermann, M.; Neufeld, J.D. Depth-Dependent Influence of Different Land-Use Systems on Bacterial Biogeography. FEMS Microbiol. Ecol. 2017, 93, fiw239. [Google Scholar] [CrossRef] [Green Version]
- Tian, Q.; Yang, X.; Wang, X.; Liao, C.; Li, Q.; Wang, M.; Wu, Y.; Liu, F. Microbial Community Mediated Response of Organic Carbon Mineralization to Labile Carbon and Nitrogen Addition in Topsoil and Subsoil. Biogeochemistry 2016, 128, 125–139. [Google Scholar] [CrossRef]
- Feng, H.; Guo, J.; Wang, W.; Song, X.; Yu, S. Soil Depth Determines the Composition and Diversity of Bacterial and Archaeal Communities in a Poplar Plantation. Forests 2019, 10, 550. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Yang, F.; Gao, J.; Peng, Z.; Chen, W. Subsoil Microbial Community Responses to Air Exposure and Legume Growth Depend on Soil Properties across Different Depths. Sci. Rep. 2019, 9, 18536. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Zhang, Q.; Tian, T.; Li, D.; Cheng, G.; Mu, J.; Wu, Q.; Niu, F.; Stegen, J.C.; An, L.; et al. Relative Roles of Deterministic and Stochastic Processes in Driving the Vertical Distribution of Bacterial Communities in a Permafrost Core from the Qinghai-Tibet Plateau, China. PLoS ONE 2015, 10, e0145747. [Google Scholar] [CrossRef] [Green Version]
- Thompson, L.R.; Sanders, J.G.; McDonald, D.; Amir, A.; Ladau, J.; Locey, K.J.; Prill, R.J.; Tripathi, A.; Gibbons, S.M.; Ackermann, G.; et al. A Communal Catalogue Reveals Earth’s Multiscale Microbial Diversity. Nature 2017, 551, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Hansen, K.; Thimonier, A.; Clarke, N.; Staelens, J.; Žlindra, D.; Waldner, P.; Marchetto, A. Atmospheric Deposition to Forest Ecosystems. In Developments in Environmental Science; Elsevier: Amsterdam, The Netherlands, 2013; Volume 12, pp. 337–374. ISBN 978-0-08-098222-9. [Google Scholar]
- Gadd, G.M.; Fomina, M.; Burford, E.P.; Lappin-Scott, H. Fungal Roles and Function in Rock, Mineral and Soil Transformations. In Micro-Organisms and Earth Systems—Advances in Geomicrobiology; Gadd, G., Semple, K., Eds.; Cambridge University Press: Cambridge, UK, 2005; pp. 201–232. ISBN 978-0-511-75485-2. [Google Scholar]
- Garcia Arredondo, M.; Lawrence, C.R.; Schulz, M.S.; Tfaily, M.M.; Kukkadapu, R.; Jones, M.E.; Boye, K.; Keiluweit, M. Root-Driven Weathering Impacts on Mineral-Organic Associations in Deep Soils over Pedogenic Time Scales. Geochim. Cosmochim. Acta 2019, 263, 68–84. [Google Scholar] [CrossRef]
- Ping, C.L.; Jastrow, J.D.; Jorgenson, M.T.; Michaelson, G.J.; Shur, Y.L. Permafrost Soils and Carbon Cycling. Soil 2015, 1, 147–171. [Google Scholar] [CrossRef] [Green Version]
- Shuster, W.D.; Subler, S.; McCoy, E.L. Deep-Burrowing Earthworm Additions Changed the Distribution of Soil Organic Carbon in a Chisel-Tilled Soil. Soil Biol. Biochem. 2001, 33, 983–996. [Google Scholar] [CrossRef]
- Tian, L.; Zhao, L.; Wu, X.; Fang, H.; Zhao, Y.; Yue, G.; Liu, G.; Chen, H. Vertical Patterns and Controls of Soil Nutrients in Alpine Grassland: Implications for Nutrient Uptake. Sci. Total Environ. 2017, 607–608, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, I.C.; Welty, A.; Blazewicz, S.J.; Bru, D.; Rouard, N.; Breuil, M.-C.; Gessler, A.; Galiano, L.; Miranda, J.C.; Spor, A.; et al. Depth Matters: Effects of Precipitation Regime on Soil Microbial Activity upon Rewetting of a Plant-Soil System. ISME J. 2018, 12, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Hu, H.; Ma, W.; Deng, Y.; Liu, Y.; Hao, B.; Zhang, X.; Dimitrov, D.; Feng, X.; Wang, Z. Contrasting Biogeographic Patterns of Bacterial and Archaeal Diversity in the Top- and Subsoils of Temperate Grasslands. mSystems 2019, 4, e00566-19. [Google Scholar] [CrossRef] [Green Version]
- Wuddivira, M.N.; Camps-Roach, G. Effects of Organic Matter and Calcium on Soil Structural Stability. Eur. J. Soil Sci. 2007, 58, 722–727. [Google Scholar] [CrossRef]
- Rowley, M.C. Calcium-Mediated Stabilisation of Soil Organic Carbon. Biogeochemistry 2018, 137, 27–49. [Google Scholar] [CrossRef] [Green Version]
- Hansel, C.M.; Fendorf, S.; Jardine, P.M.; Francis, C.A. Changes in Bacterial and Archaeal Community Structure and Functional Diversity along a Geochemically Variable Soil Profile. AEM 2008, 74, 1620–1633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolitch, O.; Feucherolles, M.; Churin, J.-L.; Fauchery, L.; Turpault, M.-P.; Uroz, S. A Microcosm Approach Highlights the Response of Soil Mineral Weathering Bacterial Communities to an Increase of K and Mg Availability. Sci. Rep. 2019, 9, 14403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sher, Y.; Baker, N.R.; Herman, D.; Fossum, C.; Hale, L.; Zhang, X.; Nuccio, E.; Saha, M.; Zhou, J.; Pett-Ridge, J.; et al. Microbial Extracellular Polysaccharide Production and Aggregate Stability Controlled by Switchgrass (Panicum Virgatum) Root Biomass and Soil Water Potential. Soil Biol. Biochem. 2020, 143, 107742. [Google Scholar] [CrossRef]
- Blevins, R.L.; Frye, W.W. Conservation Tillage: An Ecological Approach to Soil Management. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 1993; Volume 51, pp. 33–78. ISBN 978-0-12-000751-6. [Google Scholar]
- Banfield, J.F.; Barker, W.W.; Welch, S.A.; Taunton, A. Biological Impact on Mineral Dissolution: Application of the Lichen Model to Understanding Mineral Weathering in the Rhizosphere. Proc. Natl. Acad. Sci. USA 1999, 96, 3404–3411. [Google Scholar] [CrossRef] [Green Version]
- Osińska-Jaroszuk, M.; Jarosz-Wilkołazka, A.; Jaroszuk-Ściseł, J.; Szałapata, K.; Nowak, A.; Jaszek, M.; Ozimek, E.; Majewska, M. Extracellular Polysaccharides from Ascomycota and Basidiomycota: Production Conditions, Biochemical Characteristics, and Biological Properties. World J. Microbiol. Biotechnol. 2015, 31, 1823–1844. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, I.D.A.; Volpiano, C.G.; Vargas, L.K.; Granada, C.E.; Lisboa, B.B.; Passaglia, L.M.P. Use of Mineral Weathering Bacteria to Enhance Nutrient Availability in Crops: A Review. Front. Plant Sci. 2020, 11, 590774. [Google Scholar] [CrossRef] [PubMed]
- Welch, S.A.; Barker, W.W.; Banfield, J.F. Microbial Extracellular Polysaccharides and Plagioclase Dissolution. Geochim. Cosmochim. Acta 1999, 63, 1405–1419. [Google Scholar] [CrossRef]
- Liu, M.; Han, G.; Zhang, Q. Effects of Soil Aggregate Stability on Soil Organic Carbon and Nitrogen under Land Use Change in an Erodible Region in Southwest China. Int. J. Environ. Res. Public Health 2019, 16, 3809. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Wang, R.; Yang, X.; Sun, B.; Li, Q. Soil Aggregation and Aggregating Agents as Affected by Long Term Contrasting Management of an Anthrosol. Sci. Rep. 2016, 6, 39107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cania, B.; Vestergaard, G.; Krauss, M.; Fliessbach, A.; Schloter, M.; Schulz, S. A Long-Term Field Experiment Demonstrates the Influence of Tillage on the Bacterial Potential to Produce Soil Structure-Stabilizing Agents Such as Exopolysaccharides and Lipopolysaccharides. Environ. Microbiome 2019, 14, 1. [Google Scholar] [CrossRef]
- Franzluebbers, A.J.; Stuedemann, J.A.; Wilkinson, S.R. Bermudagrass Management in the Southern Piedmont USA: I. Soil and Surface Residue Carbon and Sulfur. Soil Sci. Soc. Am. J. 2001, 65, 834–841. [Google Scholar] [CrossRef]
- Ball, B.C.; Campbell, D.J.; Douglas, J.T.; Henshall, J.K.; O’Sullivan, M.F. Soil Structural Quality, Compaction and Land Management. Eur. J. Soil Sci. 2008, 48, 593–601. [Google Scholar] [CrossRef]
- Kay, B.D.; VandenBygaart, A.J. Conservation Tillage and Depth Stratification of Porosity and Soil Organic Matter. Soil Tillage Res. 2002, 66, 107–118. [Google Scholar] [CrossRef]
- Dungait, J.A.J.; Hopkins, D.W.; Gregory, A.S.; Whitmore, A.P. Soil Organic Matter Turnover Is Governed by Accessibility Not Recalcitrance. Glob. Change Biol. 2012, 18, 1781–1796. [Google Scholar] [CrossRef]
- Kuka, K.; Franko, U.; Rühlmann, J. Modelling the Impact of Pore Space Distribution on Carbon Turnover. Ecol. Model. 2007, 208, 295–306. [Google Scholar] [CrossRef]
- Maeght, J.-L.; Rewald, B.; Pierret, A. How to Study Deep Roots—and Why It Matters. Front. Plant Sci. 2013, 4, 299. [Google Scholar] [CrossRef] [Green Version]
- Van Gestel, M.; Ladd, J.N.; Amato, M. Microbial Biomass Responses to Seasonal Change and Imposed Drying Regimes at Increasing Depths of Undisturbed Topsoil Profiles. Soil Biol. Biochem. 1992, 24, 103–111. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P. A Proposed Mechanism for the Pulse in Carbon Dioxide Production Commonly Observed Following the Rapid Rewetting of a Dry Soil. Soil Sci. Soc. Am. J. 2003, 67, 798. [Google Scholar] [CrossRef]
- Young, I.M.; Crawford, J.W.; Nunan, N.; Otten, W.; Spiers, A. Chapter 4 Microbial Distribution in Soils. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2008; Volume 100, pp. 81–121. ISBN 978-0-12-374361-9. [Google Scholar]
- Garbeva, P.; van Veen, J.A.; van Elsas, J.D. Microbial Diversity in Soil: Selection of Microbial Populations by Plant and Soil Type and Implications for Disease Suppressiveness. Annu. Rev. Phytopathol. 2004, 42, 243–270. [Google Scholar] [CrossRef] [PubMed]
- Hamarashid, N.H.; Othman, M.A.; Hussain, M.-A.H. Effects of Soil Texture on Chemical Compositions, Microbial Populations, and Carbon Mineralization in Soil. Egypt. J. Exp. Biol. 2010, 6, 59–64. [Google Scholar]
- Carney, K.M.; Matson, P.A. Plant Communities, Soil Microorganisms, and Soil Carbon Cycling: Does Altering the World Belowground Matter to Ecosystem Functioning? Ecosystems 2005, 8, 928–940. [Google Scholar] [CrossRef]
- Naveed, M. Spatial Variability of Microbial Richness and Diversity and Relationships with Soil Organic Carbon, Texture and Structure across an Agricultural Field. Appl. Soil Ecol. 2016, 12, 44–55. [Google Scholar] [CrossRef]
- Dequiedt, S.; Saby, N.P.A.; Lelièvre, M.; Jolivet, C.; Thioulouse, J.; Toutain, B.; Arrouays, D.; Bispo, A.; Lemanceau, P.; Ranjard, L. Biogeographical Patterns of Soil Molecular Microbial Biomass as Influenced by Soil Characteristics and Management. Glob. Ecol. Biogeogr. 2011, 20, 641–652. [Google Scholar] [CrossRef]
- Nsabimana, D.; Haynes, R.J.; Wallis, F.M. Size, Activity and Catabolic Diversity of the Soil Microbial Biomass as Affected by Land Use. Appl. Soil Ecol. 2004, 26, 81–92. [Google Scholar] [CrossRef]
- Sun, R.; Li, W.; Dong, W.; Tian, Y.; Hu, C.; Liu, B. Tillage Changes Vertical Distribution of Soil Bacterial and Fungal Communities. Front. Microbiol. 2018, 9, 699. [Google Scholar] [CrossRef] [PubMed]
- Goberna, M.; Insam, H.; Klammer, S.; Pascual, J.A.; Sánchez, J. Microbial Community Structure at Different Depths in Disturbed and Undisturbed Semiarid Mediterranean Forest Soils. Microbiol. Ecol. 2005, 50, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Young, J.M.; Skvortsov, T.; Kelleher, B.P.; Mahaney, W.C.; Somelar, P.; Allen, C.C.R. Effect of Soil Horizon Stratigraphy on the Microbial Ecology of Alpine Paleosols. Sci. Total Environ. 2019, 657, 1183–1193. [Google Scholar] [CrossRef] [Green Version]
- Mueller, R.C.; Belnap, J.; Kuske, C.R. Soil Bacterial and Fungal Community Responses to Nitrogen Addition across Soil Depth and Microhabitat in an Arid Shrubland. Front. Microbiol. 2015, 6, 891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schütz, K.; Kandeler, E.; Nagel, P.; Scheu, S.; Ruess, L. Functional Microbial Community Response to Nutrient Pulses by Artificial Groundwater Recharge Practice in Surface Soils and Subsoils: Soil Microbial Community Response to Groundwater Recharge. FEMS Microbiol. Ecol. 2010, 72, 445–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; He, Z.; Wang, A.; Xie, J.; Wu, L.; Van Nostrand, J.D.; Jin, D.; Shao, Z.; Schadt, C.W.; Zhou, J.; et al. Divergent Responses of Forest Soil Microbial Communities under Elevated CO2 in Different Depths of Upper Soil Layers. Appl. Environ. Microbiol. 2017, 84, e01694-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Penton, C.R.; Xue, C.; Quensen, J.F.; Roley, S.S.; Guo, J.; Garoutte, A.; Zheng, T.; Tiedje, J.M. Soil Depth and Crop Determinants of Bacterial Communities under Ten Biofuel Cropping Systems. Soil Biol. Biochem. 2017, 112, 140–152. [Google Scholar] [CrossRef]
- Tecon, R.; Or, D. Biophysical Processes Supporting the Diversity of Microbial Life in Soil. FEMS Microbiol. Rev. 2017, 41, 599–623. [Google Scholar] [CrossRef]
- Zhou, J.; Xia, B.; Treves, D.S.; Wu, L.-Y.; Marsh, T.L.; O’Neill, R.V.; Palumbo, A.V.; Tiedje, J.M. Spatial and Resource Factors Influencing High Microbial Diversity in Soil. Appl. Environ. Microbiol. 2002, 68, 326–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.J.; Jeong, S.E.; Kim, P.J.; Madsen, E.L.; Jeon, C.O. High Resolution Depth Distribution of Bacteria, Archaea, Methanotrophs, and Methanogens in the Bulk and Rhizosphere Soils of a Flooded Rice Paddy. Front. Microbiol. 2015, 6, 639. [Google Scholar] [CrossRef]
- Sui, X.; Zhang, R.; Frey, B.; Yang, L.; Li, M.-H.; Ni, H. Land Use Change Effects on Diversity of Soil Bacterial, Acidobacterial and Fungal Communities in Wetlands of the Sanjiang Plain, Northeastern China. Sci. Rep. 2019, 9, 18535. [Google Scholar] [CrossRef] [PubMed]
- Bai, T.; Tao, J.; Li, Z.; Shu, M.; Yan, X.; Wang, P.; Ye, C.; Guo, H.; Wang, Y.; Hu, S. Different Microbial Responses in Top- and Sub-soils to Elevated Temperature and Substrate Addition in a Semiarid Grassland on the Loess Plateau. Eur. J. Soil Sci. 2019, 70, 1025–1036. [Google Scholar] [CrossRef]
- Potthoff, M.; Steenwerth, K.L.; Jackson, L.E.; Drenovsky, R.E.; Scow, K.M.; Joergensen, R.G. Soil Microbial Community Composition as Affected by Restoration Practices in California Grassland. Soil Biol. Biochem. 2006, 38, 1851–1860. [Google Scholar] [CrossRef] [Green Version]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an Ecological Classification of Soil Bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Kramer, C.; Gleixner, G. Soil Organic Matter in Soil Depth Profiles: Distinct Carbon Preferences of Microbial Groups during Carbon Transformation. Soil Biol. Biochem. 2008, 40, 425–433. [Google Scholar] [CrossRef]
- de Vries, F.T.; Shade, A. Controls on Soil Microbial Community Stability under Climate Change. Front. Microbiol. 2013, 4, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stone, M.M.; Kan, J.; Plante, A.F. Parent Material and Vegetation Influence Bacterial Community Structure and Nitrogen Functional Genes along Deep Tropical Soil Profiles at the Luquillo Critical Zone Observatory. Soil Biol. Biochem. 2015, 80, 273–282. [Google Scholar] [CrossRef]
- Grillo, J. Microbial Occurrence and Distribution at Different Rhizosphere Depths of Musa Sapientum var Parasidiaca and Senna Occidentalis. BMRJ 2013, 3, 73–83. [Google Scholar] [CrossRef]
- Steven, B.; Gallegos-Graves, L.V.; Belnap, J.; Kuske, C.R. Dryland Soil Microbial Communities Display Spatial Biogeographic Patterns Associated with Soil Depth and Soil Parent Material. FEMS Microbiol. Ecol. 2013, 86, 101–113. [Google Scholar] [CrossRef]
- Will, C.; Thurmer, A.; Wollherr, A.; Nacke, H.; Herold, N.; Schrumpf, M.; Gutknecht, J.; Wubet, T.; Buscot, F.; Daniel, R. Horizon-Specific Bacterial Community Composition of German Grassland Soils, as Revealed by Pyrosequencing-Based Analysis of 16S rRNA Genes. Appl. Environ. Microbiol. 2010, 76, 6751–6759. [Google Scholar] [CrossRef] [Green Version]
- Ko, D.; Yoo, G.; Yun, S.-T.; Jun, S.-C.; Chung, H. Bacterial and Fungal Community Composition across the Soil Depth Profiles in a Fallow Field. J. Ecol. Environ. 2017, 41, 34. [Google Scholar] [CrossRef] [Green Version]
- Hao, J.; Chai, Y.N.; Ordóñez, R.A.; Wright, E.E.; Archontoulis, S.; Schachtman, D. The Effects of Soil Depth on the Structure of Microbial Communities in Agricultural Soils in Iowa, USA. Appl. Environ. Microbiol. 2020, 87, e02673-20. [Google Scholar] [CrossRef]
- Pereira, A.P.D.A.; de Andrade, P.A.M.; Bini, D.; Durrer, A.; Robin, A.; Bouillet, J.P.; Andreote, F.D.; Cardoso, E.J.B.N. Shifts in the Bacterial Community Composition along Deep Soil Profiles in Monospecific and Mixed Stands of Eucalyptus Grandis and Acacia Mangium. PLoS ONE 2017, 12, e0180371. [Google Scholar] [CrossRef] [Green Version]
- Uksa, M.; Buegger, F.; Gschwendtner, S.; Lueders, T.; Kublik, S.; Kautz, T.; Athmann, M.; Köpke, U.; Munch, J.C.; Schloter, M.; et al. Bacteria Utilizing Plant-Derived Carbon in the Rhizosphere of Triticum Aestivum Change in Different Depths of an Arable Soil: Spatial Distribution of Rhizosphere Bacteria. Environ. Microbiol. Rep. 2017, 9, 729–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeBruyn, J.M.; Nixon, L.T.; Fawaz, M.N.; Johnson, A.M.; Radosevich, M. Global Biogeography and Quantitative Seasonal Dynamics of Gemmatimonadetes in Soil. Appl. Environ. Microbiol. 2011, 77, 6295–6300. [Google Scholar] [CrossRef] [Green Version]
- Fawaz, M.N. Revealing the Ecological Role of Gemmatimonadetes through Cultivation and Molecular Analysis of Agricultural Soils. Master’s Thesis, University of Tennessee, Knoxville, TN, USA, 2013. [Google Scholar]
- Daims, H.; Lebedeva, E.V.; Pjevac, P.; Han, P.; Herbold, C.; Albertsen, M.; Jehmlich, N.; Palatinszky, M.; Vierheilig, J.; Bulaev, A.; et al. Complete Nitrification by Nitrospira Bacteria. Nature 2015, 528, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Orellana, L.H.; Chee-Sanford, J.C.; Sanford, R.A.; Löffler, F.E.; Konstantinidis, K.T. Year-Round Shotgun Metagenomes Reveal Stable Microbial Communities in Agricultural Soils and Novel Ammonia Oxidizers Responding to Fertilization. Appl. Environ. Microbiol. 2017, 84, e01646-17. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, B.M.; Kim, M.; Kim, Y.; Byun, E.; Yang, J.-W.; Ahn, J.; Lee, Y.K. Variations in Bacterial and Archaeal Communities along Depth Profiles of Alaskan Soil Cores. Sci. Rep. 2018, 8, 504. [Google Scholar] [CrossRef] [Green Version]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Klenk, H.-P.; Clément, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tveit, A.; Schwacke, R.; Svenning, M.M.; Urich, T. Organic Carbon Transformations in High-Arctic Peat Soils: Key Functions and Microorganisms. ISME J. 2013, 7, 299–311. [Google Scholar] [CrossRef] [Green Version]
- Mendes, L.W.; de Lima Brossi, M.J.; Kuramae, E.E.; Tsai, S.M. Land-Use System Shapes Soil Bacterial Communities in Southeastern Amazon Region. Appl. Soil Ecol. 2015, 95, 151–160. [Google Scholar] [CrossRef]
- Kielak, A.M.; Barreto, C.C.; Kowalchuk, G.A.; van Veen, J.A.; Kuramae, E.E. The Ecology of Acidobacteria: Moving beyond Genes and Genomes. Front. Microbiol. 2016, 7, 744. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Xiong, J.; Zhang, H.; Feng, Y.; Lin, X.; Li, X.; Liang, W.; Chu, H. Soil PH Drives the Spatial Distribution of Bacterial Communities along Elevation on Changbai Mountain. Soil Biol. Biochem. 2013, 57, 204–211. [Google Scholar] [CrossRef]
- Yun, Y.; Wang, H.; Man, B.; Xiang, X.; Zhou, J.; Qiu, X.; Duan, Y.; Engel, A.S. The Relationship between PH and Bacterial Communities in a Single Karst Ecosystem and Its Implication for Soil Acidification. Front. Microbiol. 2016, 7, 1955. [Google Scholar] [CrossRef]
- Cao, P.; Zhang, L.-M.; Shen, J.-P.; Zheng, Y.-M.; Di, H.J.; He, J.-Z. Distribution and Diversity of Archaeal Communities in Selected Chinese Soils. FEMS Microbiol. Ecol. 2012, 80, 146–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemnitz, D.; Kolb, S.; Conrad, R. High Abundance of Crenarchaeota in a Temperate Acidic Forest Soil: Highly Abundant Crenarchaeota in a Forest Soil. FEMS Microbiol. Ecol. 2007, 60, 442–448. [Google Scholar] [CrossRef] [Green Version]
- Turner, S.; Mikutta, R.; Meyer-Stüve, S.; Guggenberger, G.; Schaarschmidt, F.; Lazar, C.S.; Dohrmann, R.; Schippers, A. Microbial Community Dynamics in Soil Depth Profiles Over 120,000 Years of Ecosystem Development. Front. Microbiol. 2017, 8, 874. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Li, W.; Dumbrell, A.J.; Liu, M.; Li, G.; Wu, M.; Jiang, C.; Li, Z. Spatial Variation in Soil Fungal Communities across Paddy Fields in Subtropical China. mSystems 2020, 5, e00704-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaban, B.; Ng, S.Y.M.; Jarrell, K.F. Archaeal Habitats—From the Extreme to the Ordinary. Can. J. Microbiol. 2006, 52, 73–116. [Google Scholar] [CrossRef] [PubMed]
- Uroz, S.; Ioannidis, P.; Lengelle, J.; Cébron, A.; Morin, E.; Buée, M.; Martin, F. Functional Assays and Metagenomic Analyses Reveals Differences between the Microbial Communities Inhabiting the Soil Horizons of a Norway Spruce Plantation. PLoS ONE 2013, 8, e55929. [Google Scholar] [CrossRef] [PubMed]
- Semenov, M.V.; Manucharova, N.A.; Stepanov, A.L. Distribution of Metabolically Active Prokaryotes (Archaea and Bacteria) throughout the Profiles of Chernozem and Brown Semidesert Soil. Eurasian Soil Sci. 2016, 49, 217–225. [Google Scholar] [CrossRef]
- Lazar, C.S.; Baker, B.J.; Seitz, K.; Hyde, A.S.; Dick, G.J.; Hinrichs, K.-U.; Teske, A.P. Genomic Evidence for Distinct Carbon Substrate Preferences and Ecological Niches of Bathyarchaeota in Estuarine Sediments: Genomic Content of Uncultured Benthic Bathyarchaeota. Environ. Microbiol 2016, 18, 1200–1211. [Google Scholar] [CrossRef]
- Evans, P.N.; Boyd, J.A.; Leu, A.O.; Woodcroft, B.J.; Parks, D.H.; Hugenholtz, P.; Tyson, G.W. An Evolving View of Methane Metabolism in the Archaea. Nat. Rev. Microbiol. 2019, 17, 219–232. [Google Scholar] [CrossRef]
- Pesaro, M.; Widmer, F. Identification of Novel Crenarchaeota and Euryarchaeota Clusters Associated with Different Depth Layers of a Forest Soil. FEMS Microbiol. Ecol. 2002, 42, 89–98. [Google Scholar] [CrossRef]
- Lu, X.; Seuradge, B.J.; Neufeld, J.D. Biogeography of Soil Thaumarchaeota in Relation to Soil Depth and Land Usage. FEMS Microbiol. Ecol. 2017, 93, fiw246. [Google Scholar] [CrossRef]
- Pester, M.; Schleper, C.; Wagner, M. The Thaumarchaeota: An Emerging View of Their Phylogeny and Ecophysiology. Curr. Opin. Microbiol. 2011, 14, 300–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onodera, Y.; Nakagawa, T.; Takahashi, R.; Tokuyama, T. Seasonal Change in Vertical Distribution of Ammonia-Oxidizing Archaea and Bacteria and Their Nitrification in Temperate Forest Soil. Microbes Environ. 2010, 25, 28–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousk, J.; Bååth, E. Growth of Saprotrophic Fungi and Bacteria in Soil: Growth of Saprotrophic Fungi and Bacteria in Soil. FEMS Microbiol. Ecol. 2011, 78, 17–30. [Google Scholar] [CrossRef] [Green Version]
- Degrune, F.; Theodorakopoulos, N.; Colinet, G.; Hiel, M.-P.; Bodson, B.; Taminiau, B.; Daube, G.; Vandenbol, M.; Hartmann, M. Temporal Dynamics of Soil Microbial Communities below the Seedbed under Two Contrasting Tillage Regimes. Front. Microbiol. 2017, 8, 1127. [Google Scholar] [CrossRef] [PubMed]
- Barnard, R.L.; Osborne, C.A.; Firestone, M.K. Responses of Soil Bacterial and Fungal Communities to Extreme Desiccation and Rewetting. ISME J. 2013, 7, 2229–2241. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, G.; Engelhard, M.; Müller, V. Bioenergetics of the Archaea. Microbiol. Mol. Biol. Rev. 1999, 63, 570–620. [Google Scholar] [CrossRef] [Green Version]
- Jumpponen, A.; Jones, K.L.; Blair, J. Vertical Distribution of Fungal Communities in Tallgrass Prairie Soil. Mycologia 2010, 102, 1027–1041. [Google Scholar] [CrossRef] [PubMed]
- Shigyo, N.; Umeki, K.; Hirao, T. Seasonal Dynamics of Soil Fungal and Bacterial Communities in Cool-Temperate Montane Forests. Front. Microbiol. 2019, 10, 1944. [Google Scholar] [CrossRef] [Green Version]
- Struecker, J.; Joergensen, R.G. Microorganisms and Their Substrate Utilization Patterns in Topsoil and Subsoil Layers of Two Silt Loams, Differing in Soil Organic C Accumulation Due to Colluvial Processes. Soil Biol. Biochem. 2015, 91, 310–317. [Google Scholar] [CrossRef]
- Koranda, M.; Kaiser, C.; Fuchslueger, L.; Kitzler, B.; Sessitsch, A.; Zechmeister-Boltenstern, S.; Richter, A. Fungal and Bacterial Utilization of Organic Substrates Depends on Substrate Complexity and N Availability. FEMS Microbiol. Ecol. 2014, 87, 142–152. [Google Scholar] [CrossRef]
- Malik, A.A.; Chowdhury, S.; Schlager, V.; Oliver, A.; Puissant, J.; Vazquez, P.G.M.; Jehmlich, N.; von Bergen, M.; Griffiths, R.I.; Gleixner, G. Soil Fungal:Bacterial Ratios Are Linked to Altered Carbon Cycling. Front. Microbiol. 2016, 7, 1247. [Google Scholar] [CrossRef] [Green Version]
- Rousk, J.; Brookes, P.C.; Bååth, E. Contrasting Soil PH Effects on Fungal and Bacterial Growth Suggest Functional Redundancy in Carbon Mineralization. AEM 2009, 75, 1589–1596. [Google Scholar] [CrossRef] [Green Version]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil Bacterial and Fungal Communities across a PH Gradient in an Arable Soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Strickland, M.S.; Liptzin, D.; Bradford, M.A.; Cleveland, C.C. Global Patterns in Belowground Communities. Ecol. Lett. 2009, 12, 1238–1249. [Google Scholar] [CrossRef]
- Bardgett, R.D.; McAlister, E. The Measurement of Soil Fungal:Bacterial Biomass Ratios as an Indicator of Ecosystem Self-Regulation in Temperate Meadow Grasslands. Biol. Fertil. Soils 1999, 29, 282–290. [Google Scholar] [CrossRef]
- Anderson, C.; Beare, M.; Buckley, H.L.; Lear, G. Bacterial and Fungal Communities Respond Differently to Varying Tillage Depth in Agricultural Soils. PeerJ 2017, 5, e3930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, C.; Geng, Z.; Wang, Q.; Zhang, T.; He, W.; Hou, L.; Wang, Y. Variations in Bacterial and Fungal Communities through Soil Depth Profiles in a Betula Albosinensis Forest. J. Microbiol. 2017, 55, 684–693. [Google Scholar] [CrossRef]
- Schlatter, D.C.; Kahl, K.; Carlson, B.; Huggins, D.R.; Paulitz, T. Fungal Community Composition and Diversity Vary with Soil Depth and Landscape Position in a No-till Wheat-Based Cropping System. FEMS Microbiol. Ecol. 2018, 94, fiy098. [Google Scholar] [CrossRef] [Green Version]
- Potthoff, M.; Jackson, L.E.; Steenwerth, K.L.; Ramirez, I.; Stromberg, M.R.; Rolston, D.E. Soil Biological and Chemical Properties in Restored Perennial Grassland in California. Restor Ecol. 2005, 13, 61–73. [Google Scholar] [CrossRef]
- Jobbágy, E.G.; Jackson, R.B. The Vertical Distribution of Soil Organic Carbon and Its Relation to Climate and Vegetation. Ecol. Appl. 2000, 10, 423–436. [Google Scholar] [CrossRef]
- Marty, C.; Houle, D.; Gagnon, C.; Courchesne, F. The Relationships of Soil Total Nitrogen Concentrations, Pools and C:N Ratios with Climate, Vegetation Types and Nitrate Deposition in Temperate and Boreal Forests of Eastern Canada. Catena 2017, 152, 163–172. [Google Scholar] [CrossRef]
- Emiru, N.; Gebrekidan, H. Effect of Land Use Changes and Soil Depth on Soil Organic Matter, Total Nitrogen and Available Phosphorus Contents of Soils in Senbat Watershed, Western Ethiopia. Am. J. Agric. Biol. Sci. 2013, 8, 206–212. [Google Scholar]
- Jiao, S.; Chen, W.; Wei, G. Resilience and Assemblage of Soil Microbiome in Response to Chemical Contamination Combined with Plant Growth. Appl. Environ. Microbiol. 2019, 85, e02523-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartman, K.; Tringe, S.G. Interactions between Plants and Soil Shaping the Root Microbiome under Abiotic Stress. Biochem. J. 2019, 476, 2705–2724. [Google Scholar] [CrossRef] [Green Version]
- Tkacz, A.; Bestion, E.; Bo, Z.; Hortala, M.; Poole, P.S. Influence of Plant Fraction, Soil, and Plant Species on Microbiota: A Multikingdom Comparison. mBio 2020, 11, e02785-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomson, B.C.; Ostle, N.; McNamara, N.; Bailey, M.J.; Whiteley, A.S.; Griffiths, R.I. Vegetation Affects the Relative Abundances of Dominant Soil Bacterial Taxa and Soil Respiration Rates in an Upland Grassland Soil. Microbiol. Ecol. 2010, 59, 335–343. [Google Scholar] [CrossRef]
- DeAngelis, K.M.; Brodie, E.L.; DeSantis, T.Z.; Andersen, G.L.; Lindow, S.E.; Firestone, M.K. Selective Progressive Response of Soil Microbial Community to Wild Oat Roots. ISME J. 2009, 3, 168–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wang, Z.; Jiang, P.; He, Y.; Mu, Y.; Lv, X.; Zhuang, L. Bacterial Diversity and Community Structure in the Rhizosphere of Four Ferula Species. Sci. Rep. 2018, 8, 5345. [Google Scholar] [CrossRef] [Green Version]
- Gregory, P.J. Roots, Rhizosphere and Soil: The Route to a Better Understanding of Soil Science? Roots, Rhizosphere and Soil. Eur. J. Soil Sci. 2006, 57, 2–12. [Google Scholar] [CrossRef]
- Bourceret, A.; Leyval, C.; Thomas, F.; Cébron, A. Rhizosphere Effect Is Stronger than PAH Concentration on Shaping Spatial Bacterial Assemblages along Centimetre-Scale Depth Gradients. Can. J. Microbiol. 2017, 63, 881–893. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-H.; Crowley, D.E. Rhizosphere Microbial Community Structure in Relation to Root Location and Plant Iron Nutritional Status. Appl. Environ. Microbiol. 2000, 66, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Kabir, M.D.E.; Johansen, C.; Bell, R.W. Subsoil Rhizosphere Modification by Chickpea under a Dry Topsoil: Implications for Phosphorus Acquisition. J. Plant Nutr. Soil Sci. 2015, 178, 904–913. [Google Scholar] [CrossRef] [Green Version]
- Dickey, J.R.; Fordyce, J.A.; Lebeis, S.L. Bacterial Communities of the Salvia Lyrata Rhizosphere Explained by Spatial Structure and Sampling Grain. Microbiol. Ecol. 2020, 80, 846–858. [Google Scholar] [CrossRef] [PubMed]
- Kuske, C.R.; Ticknor, L.O.; Miller, M.E.; Dunbar, J.M.; Davis, J.A.; Barns, S.M.; Belnap, J. Comparison of Soil Bacterial Communities in Rhizospheres of Three Plant Species and the Interspaces in an Arid Grassland. AEM 2002, 68, 1854–1863. [Google Scholar] [CrossRef] [Green Version]
- Uksa, M.; Schloter, M.; Endesfelder, D.; Kublik, S.; Engel, M.; Kautz, T.; Köpke, U.; Fischer, D. Prokaryotes in Subsoil—Evidence for a Strong Spatial Separation of Different Phyla by Analysing Co-Occurrence Networks. Front. Microbiol. 2015, 6, 1269. [Google Scholar] [CrossRef] [Green Version]
- Saleem, M.; Law, A.D.; Sahib, M.R.; Pervaiz, Z.H.; Zhang, Q. Impact of Root System Architecture on Rhizosphere and Root Microbiome. Rhizosphere 2018, 6, 47–51. [Google Scholar] [CrossRef]
- Rosling, A.; Landeweert, R.; Lindahl, B.D.; Larsson, K.-H.; Kuyper, T.W.; Taylor, A.F.S.; Finlay, R.D. Vertical Distribution of Ectomycorrhizal Fungal Taxa in a Podzol Soil Profile. New Phytol. 2003, 159, 775–783. [Google Scholar] [CrossRef] [PubMed]
- Muchoka, J.P.; Mugendi, D.N.; Njiruh, P.N.; Mbugua, P.K.; Njeru, E.M.; Ghilamicael, A.M.; Mutiga, M.I. Fungi Diversity in the Rhizosphere of Aspilia Pruliseta Schweif. Ext Schweif in the Semi-Arid Eastern Kenya. Air Soil Water Res. 2020, 13, 1–10. [Google Scholar] [CrossRef]
- Clemmensen, K.E.; Bahr, A.; Ovaskainen, O.; Dahlberg, A.; Ekblad, A.; Wallander, H.; Stenlid, J.; Finlay, R.D.; Wardle, D.A.; Lindahl, B.D. Roots and Associated Fungi Drive Long-Term Carbon Sequestration in Boreal Forest. Science 2013, 339, 1615–1618. [Google Scholar] [CrossRef] [PubMed]
- Upton, R.N.; Checinska Sielaff, A.; Hofmockel, K.S.; Xu, X.; Polley, H.W.; Wilsey, B.J. Soil Depth and Grassland Origin Cooperatively Shape Microbial Community Co-occurrence and Function. Ecosphere 2020, 11, e02973. [Google Scholar] [CrossRef] [Green Version]
- de Sosa, L.L.; Glanville, H.C.; Marshall, M.R.; Schnepf, A.; Cooper, D.M.; Hill, P.W.; Binley, A.; Jones, D.L. Stoichiometric Constraints on the Microbial Processing of Carbon with Soil Depth along a Riparian Hillslope. Biol. Fertil. Soils 2018, 54, 949–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, S.-R.; Doyle, A.; Holden, P.A.; Schimel, J.P. Drying and Rewetting Effects on C and N Mineralization and Microbial Activity in Surface and Subsurface California Grassland Soils. Soil Biol. Biochem. 2008, 40, 2281–2289. [Google Scholar] [CrossRef]
- Nunan, N.; Wu, K.; Young, I.M.; Crawford, J.W.; Ritz, K. In Situ Spatial Patterns of Soil Bacterial Populations, Mapped at Multiple Scales, in an Arable Soil. Microbiol. Ecol. 2002, 44, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Rey, A.; Petsikos, C.; Jarvis, P.G.; Grace, J. Effect of Temperature and Moisture on Rates of Carbon Mineralization in a Mediterranean Oak Forest Soil under Controlled and Field Conditions. Eur. J. Soil Sci. 2005, 56, 589–599. [Google Scholar] [CrossRef]
- Denef, K.; Six, J.; Paustian, K.; Merckx, R. Importance of Macroaggregate Dynamics in Controlling Soil Carbon Stabilization: Short-Term Effects of Physical Disturbance Induced by Dry–Wet Cycles. Soil Biol. Biochem. 2001, 33, 2145–2153. [Google Scholar] [CrossRef]
- Zhou, J.; Deng, Y.; Shen, L.; Wen, C.; Yan, Q.; Ning, D.; Qin, Y.; Xue, K.; Wu, L.; He, Z.; et al. Temperature Mediates Continental-Scale Diversity of Microbes in Forest Soils. Nat. Commun. 2016, 7, 12083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.K.; Sharma, R.V. Numerical Analysis for Ground Temperature Variation. Geotherm Energy 2017, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Santana, M.M.; Gonzalez, J.M. High Temperature Microbial Activity in Upper Soil Layers. FEMS Microbiol. Lett. 2015, 362, fnv182. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Feng, X.; He, J.-S.; He, H.; Lin, L.; Liu, Z. Comparing Microbial Carbon Sequestration and Priming in the Subsoil versus Topsoil of a Qinghai-Tibetan Alpine Grassland. Soil Biol. Biochem. 2017, 104, 141–151. [Google Scholar] [CrossRef]
- Cook, F.J.; Knight, J.H. Where Does Oxygen Extinction Occur in a Soil Profile? In Proceedings of the 21st International Congress on Modelling and Simulation, Gold Coast, Australia, 29 November–4 December 2015.
- Chirinda, N.; Elsgaard, L.; Thomsen, I.K.; Heckrath, G.; Olesen, J.E. Carbon Dynamics in Topsoil and Subsoil along a Cultivated Toposequence. Catena 2014, 120, 20–28. [Google Scholar] [CrossRef]
- Sexstone, A.J.; Revsbech, N.P.; Parkin, T.B.; Tiedje, J.M. Direct Measurement of Oxygen Profiles and Denitrification Rates in Soil Aggregates. Soil Sci. Soc. Am. J. 1985, 49, 645–651. [Google Scholar] [CrossRef]
- Miller, K.; Lai, C.-T.; Dahlgren, R.; Lipson, D. Anaerobic Methane Oxidation in High-Arctic Alaskan Peatlands as a Significant Control on Net CH4 Fluxes. Soil Syst. 2019, 3, 7. [Google Scholar] [CrossRef] [Green Version]
- Ganzert, L.; Jurgens, G.; Münster, U.; Wagner, D. Methanogenic Communities in Permafrost-Affected Soils of the Laptev Sea Coast, Siberian Arctic, Characterized by 16S RRNA Gene Fingerprints: Methanogenic Communities in Permafrost-Affected Soils. FEMS Microbiol. Ecol. 2007, 59, 476–488. [Google Scholar] [CrossRef] [Green Version]
- Hall, S.J.; Liptzin, D.; Buss, H.L.; DeAngelis, K.; Silver, W.L. Drivers and Patterns of Iron Redox Cycling from Surface to Bedrock in a Deep Tropical Forest Soil: A New Conceptual Model. Biogeochemistry 2016, 130, 177–190. [Google Scholar] [CrossRef] [Green Version]
- Fimmen, R.L.; Richter, D.D.B.; Vasudevan, D.; Williams, M.A.; West, L.T. Rhizogenic Fe–C Redox Cycling: A Hypothetical Biogeochemical Mechanism That Drives Crustal Weathering in Upland Soils. Biogeochemistry 2008, 87, 127–141. [Google Scholar] [CrossRef]
- Castro-Fernandez, V.; Zamora, R.; Morande, A.H.; Vallejos, G.; Gonzalez-Ordenes, F.; Guixé, V. Evolution, Metabolism and Molecular Mechanisms Underlying Extreme Adaptation of Euryarchaeota and Its Biotechnological Potential. In Archaea—New Biocatalysts, Novel Pharmaceuticals and Various Biotechnological Applications; Sghaier, H., Najjari, A., Ghedira, K., Eds.; InTech: London, UK, 2017; ISBN 978-953-51-3569-2. [Google Scholar]
- Reim, A.; Lüke, C.; Krause, S.; Pratscher, J.; Frenzel, P. One Millimetre Makes the Difference: High-Resolution Analysis of Methane-Oxidizing Bacteria and Their Specific Activity at the Oxic–Anoxic Interface in a Flooded Paddy Soil. ISME J. 2012, 6, 2128–2139. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Sekiguchi, Y.; Hanada, S.; Imachi, H.; Ohashi, A.; Harada, H.; Kamagata, Y. Anaerolinea Thermolimosa Sp. Nov., Levilinea Saccharolytica Gen. Nov., Sp. Nov. and Leptolinea Tardivitalis Gen. Nov., Sp. Nov., Novel Filamentous Anaerobes, and Description of the New Classes Anaerolineae Classis Nov. and Caldilineae Classis Nov. in the Bacterial Phylum Chloroflexi. Int. J. Syst. Evol. Microbiol. 2006, 56, 1331–1340. [Google Scholar] [CrossRef] [Green Version]
- Boutard, M.; Cerisy, T.; Nogue, P.-Y.; Alberti, A.; Weissenbach, J.; Salanoubat, M.; Tolonen, A.C. Functional Diversity of Carbohydrate-Active Enzymes Enabling a Bacterium to Ferment Plant Biomass. PLoS Genet 2014, 10, e1004773. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Sui, X.; Hu, Y.; Feng, F. Microbial Community Structure and the Relationship with Soil Carbon and Nitrogen in an Original Korean Pine Forest of Changbai Mountain, China. BMC Microbiol. 2019, 19, 218. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Nie, C.; Liu, Y.; Du, W.; He, P. Soil Microbial Community Composition Closely Associates with Specific Enzyme Activities and Soil Carbon Chemistry in a Long-Term Nitrogen Fertilized Grassland. Sci. Total Environ. 2019, 654, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Chai, H.; Yu, G.; He, N.; Wen, D.; Li, J.; Fang, J. Vertical Distribution of Soil Carbon, Nitrogen, and Phosphorus in Typical Chinese Terrestrial Ecosystems. Chin. Geogr. Sci. 2015, 25, 549–560. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Yu, G.; Zhang, X.; Wang, Q.; Ge, J.; Liu, S. Changes in Nitrogen-Cycling Microbial Communities with Depth in Temperate and Subtropical Forest Soils. Appl. Soil Ecol. 2018, 124, 218–228. [Google Scholar] [CrossRef]
- Rumpel, C.; Kögel-Knabner, I. Deep Soil Organic Matter—a Key but Poorly Understood Component of Terrestrial C Cycle. Plant Soil 2011, 338, 143–158. [Google Scholar] [CrossRef]
- Gross, C.D.; Harrison, R.B. The Case for Digging Deeper: Soil Organic Carbon Storage, Dynamics, and Controls in Our Changing World. Soil Syst. 2019, 3, 28. [Google Scholar] [CrossRef] [Green Version]
- Yan, D.; Li, J.; Pei, J.; Cui, J.; Nie, M.; Fang, C. The Temperature Sensitivity of Soil Organic Carbon Decomposition Is Greater in Subsoil than in Topsoil during Laboratory Incubation. Sci. Rep. 2017, 7, 5181. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, S.; Barot, S.; Barré, P.; Bdioui, N.; Mary, B.; Rumpel, C. Stability of Organic Carbon in Deep Soil Layers Controlled by Fresh Carbon Supply. Nature 2007, 450, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Cleveland, C.C.; Liptzin, D. C:N:P Stoichiometry in Soil: Is There a “Redfield Ratio” for the Microbial Biomass? Biogeochemistry 2007, 85, 235–252. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Manzoni, S.; Moorhead, D.L.; Richter, A. Carbon Use Efficiency of Microbial Communities: Stoichiometry, Methodology and Modelling. Ecol. Lett. 2013, 16, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Allen, A.S.; Schimel, J.P.; Holden, P.A. Controls on Microbial CO2 Production: A Comparison of Surface and Subsurface Soil Horizons. Glob. Change Biol. 2003, 9, 1322–1332. [Google Scholar] [CrossRef] [Green Version]
- Jenkinson, D.S.; Coleman, K. The Turnover of Organic Carbon in Subsoils. Part 2. Modelling Carbon Turnover. Eur. J. Soil Sci. 2008, 59, 400–413. [Google Scholar] [CrossRef]
- Chevallier, T.; Voltz, M.; Blanchart, E.; Chotte, J.L.; Eschenbrenner, V.; Mahieu, M.; Albrecht, A. Spatial and Temporal Changes of Soil C after Establishment of a Pasture on a Long-Term Cultivated (Martinique). Geoderma 2000, 94, 43–58. [Google Scholar] [CrossRef]
- Wiesmeier, M.; Prietzel, J.; Barthold, F.; Spörlein, P.; Geuß, U.; Hangen, E.; Reischl, A.; Schilling, B.; von Lützow, M.; Kögel-Knabner, I. Storage and Drivers of Organic Carbon in Forest Soils of Southeast Germany (Bavaria)—Implications for Carbon Sequestration. For. Ecol. Manag. 2013, 295, 162–172. [Google Scholar] [CrossRef]
- Eusterhues, K.; Rumpel, C.; Kleber, M.; Kögel-Knabner, I. Stabilisation of Soil Organic Matter by Interactions with Minerals as Revealed by Mineral Dissolution and Oxidative Degradation. Org. Geochem. 2003, 34, 1591–1600. [Google Scholar] [CrossRef]
- Matus, F.; Amigo, X.; Kristiansen, S.M. Aluminium Stabilization Controls Organic Carbon Levels in Chilean Volcanic Soils. Geoderma 2006, 132, 158–168. [Google Scholar] [CrossRef] [Green Version]
- Buckeridge, K.M.; Mason, K.E.; McNamara, N.P.; Ostle, N.; Puissant, J.; Goodall, T.; Griffiths, R.I.; Stott, A.W.; Whitaker, J. Environmental and Microbial Controls on Microbial Necromass Recycling, an Important Precursor for Soil Carbon Stabilization. Commun. Earth Environ. 2020, 1, 36. [Google Scholar] [CrossRef]
- Peixoto, L.; Elsgaard, L.; Rasmussen, J.; Kuzyakov, Y.; Banfield, C.C.; Dippold, M.A.; Olesen, J.E. Decreased Rhizodeposition, but Increased Microbial Carbon Stabilization with Soil Depth down to 3.6 m. Soil Biol. Biochem. 2020, 150, 108008. [Google Scholar] [CrossRef]
- Bernal, B.; McKinley, D.C.; Hungate, B.A.; White, P.M.; Mozdzer, T.J.; Megonigal, J.P. Limits to Soil Carbon Stability; Deep, Ancient Soil Carbon Decomposition Stimulated by New Labile Organic Inputs. Soil Biol. Biochem. 2016, 98, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Marschner, B.; Brodowski, S.; Dreves, A.; Gleixner, G.; Gude, A.; Grootes, P.M.; Hamer, U.; Heim, A.; Jandl, G.; Ji, R.; et al. How Relevant Is Recalcitrance for the Stabilization of Organic Matter in Soils? J. Plant Nutr. Soil Sci. 2008, 171, 91–110. [Google Scholar] [CrossRef] [Green Version]
- Mikutta, R.; Kleber, M.; Torn, M.S.; Jahn, R. Stabilization of Soil Organic Matter: Association with Minerals or Chemical Recalcitrance? Biogeochemistry 2006, 77, 25–56. [Google Scholar] [CrossRef]
- Wagai, R.; Mayer, L.M. Sorptive Stabilization of Organic Matter in Soils by Hydrous Iron Oxides. Geochim. Cosmochim. Acta 2007, 71, 25–35. [Google Scholar] [CrossRef]
- Rey, A.; Pegoraro, E.; Jarvis, P.G. Carbon Mineralization Rates at Different Soil Depths across a Network of European Forest Sites (FORCAST). Eur. J. Soil Sci. 2008, 59, 1049–1062. [Google Scholar] [CrossRef]
- Heitkötter, J.; Heinze, S.; Marschner, B. Relevance of Substrate Quality and Nutrients for Microbial C-Turnover in Top- and Subsoil of a Dystric Cambisol. Geoderma 2017, 302, 89–99. [Google Scholar] [CrossRef]
- Heitkötter, J.; Niebuhr, J.; Heinze, S.; Marschner, B. Patterns of Nitrogen and Citric Acid Induced Changes in C-Turnover and Enzyme Activities Are Different in Topsoil and Subsoils of a Sandy Cambisol. Geoderma 2017, 292, 111–117. [Google Scholar] [CrossRef]
- Bossio, D.A.; Scow, K.M. Impacts of Carbon and Flooding on Soil Microbial Communities: Phospholipid Fatty Acid Profiles and Substrate Utilization Patterns. Microbiol. Ecol. 1998, 35, 265–278. [Google Scholar] [CrossRef]
- Kemmitt, S.J.; Lanyon, C.V.; Waite, I.S.; Wen, Q.; Addiscott, T.M.; Bird, N.R.A.; O’Donnell, A.G.; Brookes, P.C. Mineralization of Native Soil Organic Matter Is Not Regulated by the Size, Activity or Composition of the Soil Microbial Biomass—A New Perspective. Soil Biol. Biochem. 2008, 40, 61–73. [Google Scholar] [CrossRef]
- Sanaullah, M.; Chabbi, A.; Leifeld, J.; Bardoux, G.; Billou, D.; Rumpel, C. Decomposition and Stabilization of Root Litter in Top- and Subsoil Horizons: What Is the Difference? Plant Soil 2011, 338, 127–141. [Google Scholar] [CrossRef]
- Liang, Z.; Elsgaard, L.; Nicolaisen, M.H.; Lyhne-Kjærbye, A.; Olesen, J.E. Carbon Mineralization and Microbial Activity in Agricultural Topsoil and Subsoil as Regulated by Root Nitrogen and Recalcitrant Carbon Concentrations. Plant Soil 2018, 433, 65–82. [Google Scholar] [CrossRef] [Green Version]
- Fierer, N.; Craine, J.M.; McLauchlan, K.; Schimel, J.P. Litter Quality and the Temperature Sensitivity of Decomposition. Ecology 2005, 86, 320–326. [Google Scholar] [CrossRef] [Green Version]
- Craine, J.; Spurr, R.; McLauchlan, K.; Fierer, N. Landscape-Level Variation in Temperature Sensitivity of Soil Organic Carbon Decomposition. Soil Biol. Biochem. 2010, 42, 373–375. [Google Scholar] [CrossRef]
- Xu, W.; Li, W.; Jiang, P.; Wang, H.; Bai, E. Distinct Temperature Sensitivity of Soil Carbon Decomposition in Forest Organic Layer and Mineral Soil. Sci. Rep. 2015, 4, 6512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delmont, T.O.; Prestat, E.; Keegan, K.P.; Faubladier, M.; Robe, P.; Clark, I.M.; Pelletier, E.; Hirsch, P.R.; Meyer, F.; Gilbert, J.A.; et al. Structure, Fluctuation and Magnitude of a Natural Grassland Soil Metagenome. ISME J. 2012, 6, 1677–1687. [Google Scholar] [CrossRef] [Green Version]
- Diamond, S.; Andeer, P.F.; Li, Z.; Crits-Christoph, A.; Burstein, D.; Anantharaman, K.; Lane, K.R.; Thomas, B.C.; Pan, C.; Northen, T.R.; et al. Mediterranean Grassland Soil C–N Compound Turnover Is Dependent on Rainfall and Depth, and Is Mediated by Genomically Divergent Microorganisms. Nat. Microbiol. 2019, 4, 1356–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, E.R.; Hatt, J.K.; He, Z.; Wu, L.; Guo, X.; Luo, Y.; Schuur, E.A.G.; Tiedje, J.M.; Zhou, J.; Konstantinidis, K.T. Responses of Tundra Soil Microbial Communities to Half a Decade of Experimental Warming at Two Critical Depths. Proc. Natl. Acad. Sci. USA 2019, 116, 15096–15105. [Google Scholar] [CrossRef] [Green Version]
- Delmont, T.O.; Robe, P.; Cecillon, S.; Clark, I.M.; Constancias, F.; Simonet, P.; Hirsch, P.R.; Vogel, T.M. Accessing the Soil Metagenome for Studies of Microbial Diversity. Appl. Environ. Microbiol. 2011, 77, 1315–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finzi, A.C.; Abramoff, R.Z.; Spiller, K.S.; Brzostek, E.R.; Darby, B.A.; Kramer, M.A.; Phillips, R.P. Rhizosphere Processes Are Quantitatively Important Components of Terrestrial Carbon and Nutrient Cycles. Glob. Change Biol. 2015, 21, 2082–2094. [Google Scholar] [CrossRef] [PubMed]
- Tückmantel, T.; Leuschner, C.; Preusser, S.; Kandeler, E.; Angst, G.; Mueller, C.W.; Meier, I.C. Root Exudation Patterns in a Beech Forest: Dependence on Soil Depth, Root Morphology, and Environment. Soil Biol. Biochem. 2017, 107, 188–197. [Google Scholar] [CrossRef]
- Proctor, C.; He, Y. Modeling Root Exudate Accumulation Gradients to Estimate Net Exudation Rates by Peatland Soil Depth. Plants 2021, 10, 106. [Google Scholar] [CrossRef]
- Qin, H.L.; Zhang, Z.X.; Lu, J.; Zhu, Y.J.; Webster, R.; Liu, X.L.; Yuan, H.Z.; Hou, H.J.; Chen, C.L.; Wei, W.X. Change from Paddy Rice to Vegetable Growing Changes Nitrogen-Cycling Microbial Communities and Their Variation with Depth in the Soil: Microbial Communities’ Response to Change of Land Use. Eur. J. Soil Sci. 2016, 67, 650–658. [Google Scholar] [CrossRef]
- Lu, S.; Liu, X.; Ma, Z.; Liu, Q.; Wu, Z.; Zeng, X.; Shi, X.; Gu, Z. Vertical Segregation and Phylogenetic Characterization of Ammonia-Oxidizing Bacteria and Archaea in the Sediment of a Freshwater Aquaculture Pond. Front. Microbiol. 2016, 6, 1539. [Google Scholar] [CrossRef]
- Feng, J.; Penton, C.R.; He, Z.; Van Nostrand, J.D.; Yuan, M.M.; Wu, L.; Wang, C.; Qin, Y.; Shi, Z.J.; Guo, X.; et al. Long-Term Warming in Alaska Enlarges the Diazotrophic Community in Deep Soils. mBio 2019, 10, e02521-18. [Google Scholar] [CrossRef] [Green Version]
- Levy-Booth, D.J.; Prescott, C.E.; Grayston, S.J. Microbial Functional Genes Involved in Nitrogen Fixation, Nitrification and Denitrification in Forest Ecosystems. Soil Biol. Biochem. 2014, 75, 11–25. [Google Scholar] [CrossRef]
- Uksa, M.; Fischer, D.; Welzl, G.; Kautz, T.; Köpke, U.; Schloter, M. Community Structure of Prokaryotes and Their Functional Potential in Subsoils Is More Affected by Spatial Heterogeneity than by Temporal Variations. Soil Biol. Biochem. 2014, 75, 197–201. [Google Scholar] [CrossRef]
- He, J.-Z.; Hu, H.-W.; Zhang, L.-M. Current Insights into the Autotrophic Thaumarchaeal Ammonia Oxidation in Acidic Soils. Soil Biol. Biochem. 2012, 55, 146–154. [Google Scholar] [CrossRef]
- Zhang, L.-M.; Hu, H.-W.; Shen, J.-P.; He, J.-Z. Ammonia-Oxidizing Archaea Have More Important Role than Ammonia-Oxidizing Bacteria in Ammonia Oxidation of Strongly Acidic Soils. ISME J. 2012, 6, 1032–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzyakov, Y. Priming Effects: Interactions between Living and Dead Organic Matter. Soil Biol. Biochem. 2010, 42, 1363–1371. [Google Scholar] [CrossRef]
- Karhu, K.; Hilasvuori, E.; Fritze, H.; Biasi, C.; Nykänen, H.; Liski, J.; Vanhala, P.; Heinonsalo, J.; Pumpanen, J. Priming Effect Increases with Depth in a Boreal Forest Soil. Soil Biol. Biochem. 2016, 99, 104–107. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L. Phenol Oxidase, Peroxidase and Organic Matter Dynamics of Soil. Soil Biol. Biochem. 2010, 42, 391–404. [Google Scholar] [CrossRef]
- Blagodatskaya, E.; Kuzyakov, Y. Mechanisms of Real and Apparent Priming Effects and Their Dependence on Soil Microbial Biomass and Community Structure: Critical Review. Biol. Fertil. Soils 2008, 45, 115–131. [Google Scholar] [CrossRef]
- Craine, J.M.; Morrow, C.; Fierer, N. Microbial nitrogen limitation increases decomposition. Ecology 2007, 88, 2105–2113. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Senbayram, M.; Blagodatsky, S.; Myachina, O.; Dittert, K.; Lin, X.; Blagodatskaya, E.; Kuzyakov, Y. Soil C and N Availability Determine the Priming Effect: Microbial N Mining and Stoichiometric Decomposition Theories. Glob. Change Biol. 2014, 20, 2356–2367. [Google Scholar] [CrossRef] [PubMed]
- Lomander, A.; Kätterer, T.; Andrén, O. Modelling the Effects of Temperature and Moisture on CO2 Evolution from Top- and Subsoil Using a Multi-Compartment Approach. Soil Biol. Biochem. 1998, 30, 2023–2030. [Google Scholar] [CrossRef]
- van der Voort, T.S.; Mannu, U.; Hagedorn, F.; McIntyre, C.; Walthert, L.; Schleppi, P.; Haghipour, N.; Eglinton, T.I. Dynamics of deep soil carbon—Insights from 14C time series across a climatic gradient. Biogeosciences 2019, 16, 3233–3246. [Google Scholar] [CrossRef] [Green Version]
- Baumert, V.L.; Vasilyeva, N.A.; Vladimirov, A.A.; Meier, I.C.; Kögel-Knabner, I.; Mueller, C.W. Root Exudates Induce Soil Macroaggregation Facilitated by Fungi in Subsoil. Front. Environ. Sci. 2018, 6, 140. [Google Scholar] [CrossRef]
- Abramoff, R.; Xu, X.; Hartman, M.; O’Brien, S.; Feng, W.; Davidson, E.; Finzi, A.; Moorhead, D.; Schimel, J.; Torn, M.; et al. The Millennial Model: In Search of Measurable Pools and Transformations for Modeling Soil Carbon in the New Century. Biogeochemistry 2018, 137, 51–71. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naylor, D.; McClure, R.; Jansson, J. Trends in Microbial Community Composition and Function by Soil Depth. Microorganisms 2022, 10, 540. https://doi.org/10.3390/microorganisms10030540
Naylor D, McClure R, Jansson J. Trends in Microbial Community Composition and Function by Soil Depth. Microorganisms. 2022; 10(3):540. https://doi.org/10.3390/microorganisms10030540
Chicago/Turabian StyleNaylor, Dan, Ryan McClure, and Janet Jansson. 2022. "Trends in Microbial Community Composition and Function by Soil Depth" Microorganisms 10, no. 3: 540. https://doi.org/10.3390/microorganisms10030540
APA StyleNaylor, D., McClure, R., & Jansson, J. (2022). Trends in Microbial Community Composition and Function by Soil Depth. Microorganisms, 10(3), 540. https://doi.org/10.3390/microorganisms10030540