
Effect of Input C/N Ratio on Bacterial Community of Water Biofloc and Shrimp Gut in a Commercial Zero-Exchange System with Intensive Production of Penaeus vannamei

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Trial Shrimp and System Preparation
2.2. Trial Design and Culture Management
2.3. Water Quality Monitoring and Shrimp Performance Determination
2.4. Microbial Sampling, DNA Extraction and qPCR Analysis
2.5. HiSeq Sequencing and Data Processing
2.6. Statistical Analysis
3. Results
3.1. Water Quality and Shrimp Performance in Intensive BFT Systems
3.2. Effect of C/N Ratio on Nitrogen Dynamics and Biofloc Management
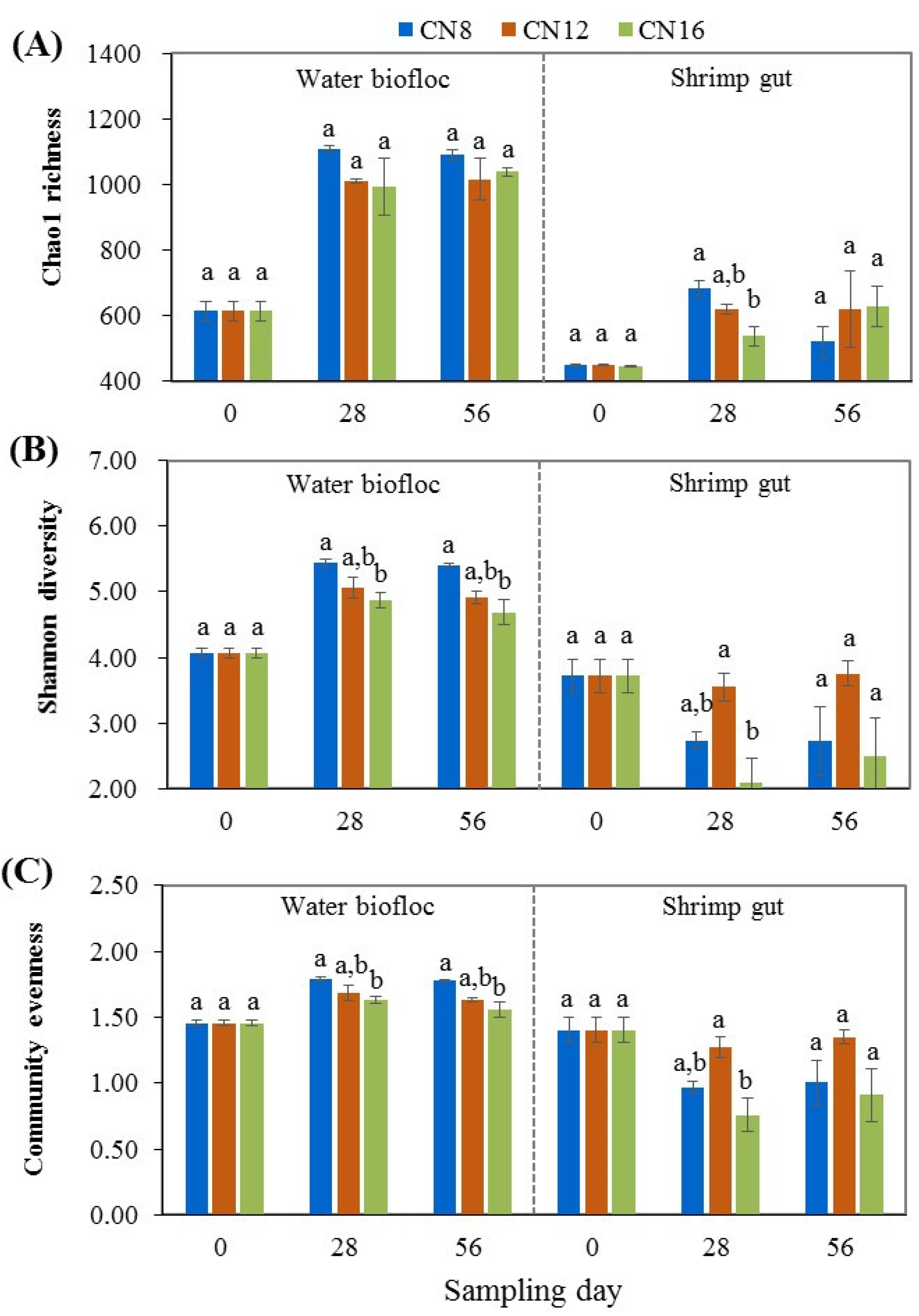
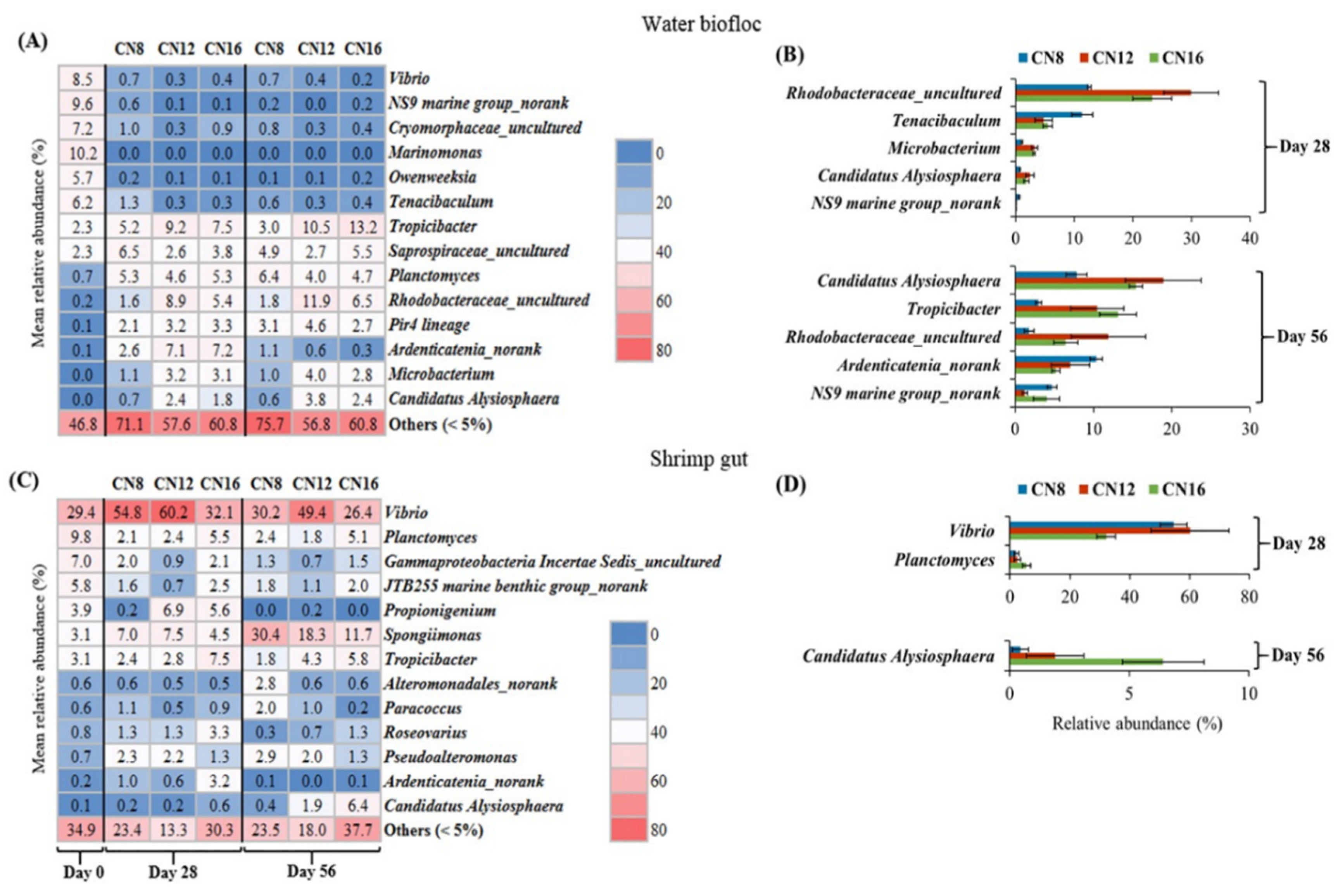
3.3. Effect of C/N Ratio on Bacterial Community in Water Biofloc and Shrimp Gut
3.4. Relationship of Bacterial Community between Water Biofloc and Shrimp Gut
4. Discussion
4.1. C/N Ratio Affected Insignificantly on Shrimp Production Performance in Intensive BFT System
4.2. C/N Ratio Altered Nitrogen Dynamics and Culture Management in Intensive BFT System
4.3. C/N Ratio Affected the Bacterial Community in Water Biofloc and Shrimp Gut
4.4. High Similarity of Bacterial Community between Water Biofloc and Shrimp Gut
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO (Food and Agriculture Organization of the United Nations). Global Aquaculture Production 1950–2019 (Online Query); FAO Fisheries and Aquaculture Department: Rome, Italy, 2021; Available online: https://www.fao.org/fishery/statistics-query/en/aquaculture (accessed on 15 March 2021).
- Browdy, C.L.; Venero, J.A.; Stokes, A.D.; Leffler, J.W. Superintensive Biofloc production systems technologies for Marine Shrimp Litopenaeus vannamei: Technical challenges and opportunities. In New Technologies in Aquaculture; Burnell, G., Allan, G.L., Eds.; Woodhead Publishing: Cambridge, UK, 2009; pp. 1010–1028. [Google Scholar]
- Kumar, G.; Engle, C.R. Technological advances that led to growth of shrimp, salmon, and tilapia farming. Rev. Fish. Sci. Aquac. 2016, 24, 136–152. [Google Scholar] [CrossRef]
- El-Sayed, A.F.M. Use of biofloc technology in shrimp aquaculture: A comprehensive review, with emphasis on the last decade. Rev. Aquacult. 2021, 13, 676–705. [Google Scholar] [CrossRef]
- Samocha, T.M. Sustainable Biofloc Systems for Marine Shrimp; Academic Press: Cambridge, UK, 2019. [Google Scholar] [CrossRef]
- Walker, D.A.U.; Suazo, M.C.M.; Emerenciano, M.G.C. Biofloc technology: Principles focused on potential species and the case study of Chilean river shrimp Cryphiops caementarius. Rev. Aquac. 2020, 12, 1759–1782. [Google Scholar] [CrossRef]
- Avnimelech, Y. Biofloc Technology—A Practical Guide Book, 3rd ed.; The World Aquaculture Society: Baton Rouge, LA, USA, 2015. [Google Scholar]
- Bossier, P.; Ekasari, J. Biofloc technology application in aquaculture to support sustainable development goals. Microb. Biotechnol. 2017, 10, 1012–1016. [Google Scholar] [CrossRef] [PubMed]
- Khanjani, M.H.; Sharifinia, M. Biofloc technology as a promising tool to improve aquaculture production. Rev. Aquac. 2020, 12, 1836–1850. [Google Scholar] [CrossRef]
- Abakari, G.; Wu, X.; He, X.; Fan, L.P.; Luo, G.Z. Bacteria in biofloc technology aquaculture systems: Roles and mediating factors. Rev. Aquac. 2022; in press. [Google Scholar] [CrossRef]
- Samocha, T.M.; Prangnell, D.I.; Hanson, T.R.; Treece, G.D.; Morris, T.C.; Castro, L.F.; Staresinic, N. Design and Operation of Super Intensive, Biofloc-Dominated Systems for Indoor Production of the Pacific White Shrimp, Litopenaeus vannamei—The Texas A&M AgriLife Research Experience; The World Aquaculture Society: Baton Rouge, LA, USA, 2017. [Google Scholar]
- Kumar, V.; Roy, S.; Behera, B.K.; Swain, H.S.; Das, B.K. Biofloc Microbiome with Bioremediation and Health Benefits. Front. Microbiol. 2021, 12, 741164. [Google Scholar] [CrossRef]
- Dauda, A.B. Biofloc technology: A review on the microbial interactions, operational parameters and implications to disease and health management of cultured aquatic animals. Rev. Aquac. 2020, 12, 1193–1210. [Google Scholar] [CrossRef]
- Xu, W.; Xu, Y.; Su, H.; Hu, X.; Xu, Y.; Li, Z.; Wen, G.; Cao, Y. Production performance, inorganic nitrogen control and bacterial community characteristics in a controlled biofloc-based system for indoor and outdoor super-intensive culture of Litopenaeus vannamei. Aquaculture 2021, 531, 735749. [Google Scholar] [CrossRef]
- Xu, W.J.; Morris, T.C.; Samocha, T.M. Effects of C/N ratio on biofloc development, water quality, and performance of Litopenaeus vannamei juveniles in a biofloc-based, high-density, zero-exchange, outdoor tank system. Aquaculture 2016, 453, 169–175. [Google Scholar] [CrossRef]
- Avnimelech, Y. Carbon nitrogen ratio as a control element in aquaculture systems. Aquaculture 1999, 176, 227–235. [Google Scholar] [CrossRef]
- Emerenciano, M.G.C.; Martínez-Corodva, L.R.; Martínez-Porchas, M.; Miranda-Baeza, A. Biofloc Technology (BFT): A tool for water quality management in aquaculture. In Water Quality; Tutu, H., Ed.; IntechOpen: London, UK, 2017; pp. 91–109. [Google Scholar] [CrossRef] [Green Version]
- Abakari, G.; Luo, G.; Kombat, E.O.; Alhassan, E.H. Supplemental carbon sources applied in biofloc technology aquaculture systems: Types, effects and future research. Rev. Aquac. 2021, 13, 1193–1222. [Google Scholar] [CrossRef]
- Ebeling, J.M.; Timmons, M.B.; Bisogni, J.J. Engineering analysis of the stoichiometry of photoautotrophic, autotrophic, and heterotrophic removal of ammonia-nitrogen in aquaculture systems. Aquaculture 2006, 257, 346–358. [Google Scholar] [CrossRef] [Green Version]
- Ray, A.J.; Lewis, B.L.; Browdy, C.L.; Leffler, J.W. Suspended solids removal to improve shrimp (Litopenaeus vannamei) production and an evaluation of a plant-based feed in minimal-exchange, superintensive culture systems. Aquaculture 2010, 299, 89–98. [Google Scholar] [CrossRef]
- Panigrahi, A.; Saranya, C.; Sundaram, M.; Vinoth Kannan, S.R.; Das, R.R.; Satish Kumar, R.; Rajesh, P.; Otta, S.K. Carbon: Nitrogen (C:N) ratio level variation influences microbial community of the system and growth as well as immunity of shrimp (Litopenaeus vannamei) in biofloc based culture system. Fish Shellfish Immunol. 2018, 81, 329–337. [Google Scholar] [CrossRef]
- Huerta-Rábago, J.A.; Martínez-Porchas, M.; Miranda-Baeza, A.; Nieves-Soto, M.; Rivas-Vega, M.E.; Martínez-Córdova, L.R. Addition of commercial probiotic in a biofloc shrimp farm of Litopenaeus vannamei during the nursery phase: Effect on bacterial diversity using massive sequencing 16S rRNA. Aquaculture 2019, 502, 391–399. [Google Scholar] [CrossRef]
- Vargas-Albores, F.; Martínez-Córdova, L.R.; Gollas-Galván, T.; Garibay-Valdez, E.; Emerenciano, M.G.C.; Lago-Leston, A. Inferring the functional properties of bacterial communities in shrimp-culture bioflocs produced with amaranth and wheat seeds as fouler promoters. Aquaculture 2019, 500, 107–117. [Google Scholar] [CrossRef]
- Jiang, W.; Ren, W.; Li, L.; Dong, S.; Tian, X. Light and carbon sources addition alter microbial community in biofloc-based Litopenaeus vannamei culture systems. Aquaculture 2020, 515, 734572. [Google Scholar] [CrossRef]
- Xu, W.J.; Xu, Y.; Huang, X.S.; Hu, X.J.; Xu, Y.N.; Su, H.C.; Li, Z.J.; Yang, K.; Wen, G.L.; Cao, Y.C. Addition of algicidal bacterium CZBC1 and molasses to inhibit cyanobacteria and improve microbial communities, water quality and shrimp performance in culture systems. Aquaculture 2019, 502, 303–311. [Google Scholar] [CrossRef]
- Zhang, X.C.; Li, X.H.; Lu, J.Q.; Qiu, Q.F.; Chen, J.; Xiong, J.B. Quantifying the importance of external and internal sources to the gut microbiota in juvenile and adult shrimp. Aquaculture 2021, 531, 735910. [Google Scholar] [CrossRef]
- Zhou, L.; Qu, Y.; Qin, J.G.; Chen, L.; Han, F.; Li, E. Deep insight into bacterial community characterization and relationship in the pond water, sediment and the gut of shrimp (Penaeus japonicus). Aquaculture 2021, 539, 736658. [Google Scholar] [CrossRef]
- Chaiyapechara, S.; Rungrassamee, W.; Suriyachay, I.; Kuncharin, Y.; Klanchui, A.; Karoonuthaisiri, N.; Jiravanichpaisal, P. Bacterial community associated with the intestinal tract of P. monodon in commercial farms. Microb. Ecol. 2012, 63, 938–953. [Google Scholar] [CrossRef] [PubMed]
- Cardona, E.; Gueguen, Y.; Magré, K.; Lorgeoux, B.; Piquemal, D.; Pierrat, F.; Noguier, F.; Saulnier, D. Bacterial community characterization of water and intestine of the shrimp Litopenaeus stylirostris in a biofloc system. BMC Microbiol. 2016, 16, 157. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.P.; Huang, L.; Hu, S.T.; Chen, C.; Huang, X.L.; Liu, W.; Wang, S.P.; Zhu, Y.Y.; Zhao, Y.J.; Zhang, D.M. Effects of carbon/nitrogen ratio on growth, intestinal microbiota and metabolome of shrimp (Litopenaeus vannamei). Front. Microbiol. 2020, 11, 652. [Google Scholar] [CrossRef]
- Huang, L.; Guo, H.P.; Chen, C.; Huang, X.L.; Chen, W.; Bao, F.J.; Liu, W.; Wang, S.P.; Zhang, D.M. The bacteria from large-sized bioflocs are more associated with the shrimp gut microbiota in culture system. Aquaculture 2020, 523, 735159. [Google Scholar] [CrossRef]
- APHA. Standard Methods for the Examination of the Water and Wastewater, 22nd ed.; American Public Health Association: Washington, DC, USA, 1998. [Google Scholar]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every base matters: Assessing small subunit rRNA primers for marine microbiomes with mock communities, time series and global field samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef]
- Xu, W.J.; Xu, Y.; Su, H.C.; Hu, X.J.; Yang, K.; Wen, G.L.; Cao, Y.C. Characteristics of Ammonia Removal and Nitrifying Microbial Communities in a Hybrid Biofloc-RAS for Intensive Litopenaeus vannamei Culture: A Pilot-Scale Study. Water 2020, 12, 3000. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Jost, L. The relation between evenness and diversity. Diversity 2010, 2, 207–232. [Google Scholar] [CrossRef]
- Ray, A.J.; Dillon, K.S.; Lotz, J.M. Water quality dynamics and shrimp (Litopenaeus vannamei) production in intensive, mesohaline culture systems with two levels of biofloc management. Aquac. Eng. 2011, 45, 127–136. [Google Scholar] [CrossRef]
- Schveitzer, R.; Arantes, R.; Costodio, P.F.S.; Santo, C.M.D.; Arana, L.V.; Seiffert, W.Q.; Andreatta, E.R. Effect of different biofloc levels on microbial activity, water quality and performance of Litopenaeus vannamei in a tank system operated with no water exchange. Aquac. Eng. 2013, 56, 59–70. [Google Scholar] [CrossRef]
- Furtado, P.S.; Poersch, L.H.; Wasielesky, W. Effect of calcium hydroxide, carbonate and sodium bicarbonate on water quality and zootechnical performance of shrimp Litopenaeus vannamei reared in bio-flocs technology (BFT) systems. Aquaculture 2012, 321, 130–135. [Google Scholar] [CrossRef]
- Xu, W.; Xu, Y.; Su, H.; Hu, X.; Xu, Y.; Li, Z.; Wen, G.; Cao, Y. Effects of feeding frequency on growth, feed utilization, digestive enzyme activity and body composition of Litopenaeus vannamei in biofloc-based zero-exchange intensive systems. Aquaculture 2020, 522, 735079. [Google Scholar] [CrossRef]
- Da Silva, K.R.; Wasielesky, W.; Abreu, P.C. Nitrogen and Phosphorus Dynamics in the Biofloc Production of the Pacific White Shrimp, Litopenaeus vannamei. J. World Aquac. Soc. 2013, 44, 30–41. [Google Scholar] [CrossRef]
- Ray, A.J.; Seaborn, G.; Leffler, J.W.; Wilde, S.B.; Lawson, A.; Browdy, C.L. Characterization of microbial communities in minimal-exchange, intensive aquaculture systems and the effects of suspended solids management. Aquaculture 2010, 310, 130–138. [Google Scholar] [CrossRef]
- Tatari, K.; Musovic, S.; Gülay, A.; Dechesne, A.; Albrechtsen, H.; Smets, B.F. Density and distribution of nitrifying guilds in rapid sand filters for drinking water production: Dominance of Nitrospira spp. Water Res. 2017, 127, 239–248. [Google Scholar] [CrossRef] [Green Version]
- Shang, Q.; Tang, H.; Wang, Y.; Yu, K.; Wang, L.; Zhang, R.; Wang, S.; Xue, R.; Wei, C. Application of enzyme-hydrolyzed cassava dregs as a carbon source in aquaculture. Sci. Total Environ. 2018, 615, 681–690. [Google Scholar] [CrossRef]
- Wei, Y.F.; Liao, S.A.; Wang, A.L. The effect of different carbon sources on the nutritional composition, microbial community and structure of bioflocs. Aquaculture 2016, 465, 88–93. [Google Scholar] [CrossRef]
- Tepaamorndech, S.; Nookaew, I.; Higdon, S.M.; Santiyanont, P.; Visessanguan, W. Metagenomics in bioflocs and their effects on gut microbiome and immune responses in pacific white shrimp. Fish Shellfish Immunol. 2020, 106, 733–741. [Google Scholar] [CrossRef]
- Krummenauer, D.; Abreu, P.C.; Poersch, L.; Reis, P.; Wasielesky, W. The relationship between shrimp (litopenaeus vannamei) size and biofloc consumption determined by the stable isotope technique. Aquaculture 2020, 529, 735635. [Google Scholar] [CrossRef]
- Garibay-Valdez, E.; Martínez-Córdova, L.R.; López-Torres, M.A.; Almendariz-Tapia, F.J.; Martínez-Porchas, M.; Calderón, K. The implication of metabolically active vibrio spp. in the digestive tract of Litopenaeus vannamei for its post-larval development. Sci. Rep. 2020, 10, 11428. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.N.; Wu, J.H.; Zhang, J.J.; Yang, W.; Chen, J.; Xiong, J.B. A meta-analysis reveals universal gut bacterial signatures for diagnosing the incidence of shrimp disease, FEMS Microbiol. Ecol. 2018, 94, fiy147. [Google Scholar] [CrossRef] [Green Version]
- Bueno, E.; Sit, B.; Waldor, M.K.; Cava, F. Anaerobic nitrate reduction divergently governs population expansion of the enteropathogen Vibrio cholerae. Nat. Microbiol. 2018, 3, 1346–1353. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Harvest Weight (g) | Growth Rate (g wk−1) | Survival Rate (%) | Yield (kg m−3) | Feed Conversion Ratio |
|---|---|---|---|---|---|
| CN8 | 15.93 ± 0.30 | 1.66 ± 0.04 | 92.8 ± 2.7 | 4.43 ± 0.18 | 1.43 ± 0.03 |
| CN12 | 16.31 ± 0.60 | 1.70 ± 0.07 | 92.4 ± 3.1 | 4.52 ± 0.15 | 1.38 ± 0.01 |
| CN16 | 16.79 ± 1.10 | 1.76 ± 0.14 | 89.7 ± 3.0 | 4.51 ± 0.15 | 1.39 ± 0.03 |
| Group | CN8 | CN12 | CN16 |
|---|---|---|---|
| Foam fractionator operation time (h) | 336 ± 15 a | 409 ± 27 b | 510 ± 31 c |
| Sodium carbonate addition amount (kg) | 26 ± 3 a | 29 ± 4 a | 39 ± 6 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, W.; Wen, G.; Su, H.; Xu, Y.; Hu, X.; Cao, Y. Effect of Input C/N Ratio on Bacterial Community of Water Biofloc and Shrimp Gut in a Commercial Zero-Exchange System with Intensive Production of Penaeus vannamei. Microorganisms 2022, 10, 1060. https://doi.org/10.3390/microorganisms10051060
Xu W, Wen G, Su H, Xu Y, Hu X, Cao Y. Effect of Input C/N Ratio on Bacterial Community of Water Biofloc and Shrimp Gut in a Commercial Zero-Exchange System with Intensive Production of Penaeus vannamei. Microorganisms. 2022; 10(5):1060. https://doi.org/10.3390/microorganisms10051060
Chicago/Turabian StyleXu, Wujie, Guoliang Wen, Haochang Su, Yu Xu, Xiaojuan Hu, and Yucheng Cao. 2022. "Effect of Input C/N Ratio on Bacterial Community of Water Biofloc and Shrimp Gut in a Commercial Zero-Exchange System with Intensive Production of Penaeus vannamei" Microorganisms 10, no. 5: 1060. https://doi.org/10.3390/microorganisms10051060
APA StyleXu, W., Wen, G., Su, H., Xu, Y., Hu, X., & Cao, Y. (2022). Effect of Input C/N Ratio on Bacterial Community of Water Biofloc and Shrimp Gut in a Commercial Zero-Exchange System with Intensive Production of Penaeus vannamei. Microorganisms, 10(5), 1060. https://doi.org/10.3390/microorganisms10051060