
Live Influenza Vaccine Provides Early Protection against Homologous and Heterologous Influenza and May Prevent Post-Influenza Pneumococcal Infections in Mice

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Influenza Viruses
2.3. Infectious Pneumococci
2.4. Immunization and Sample Collection
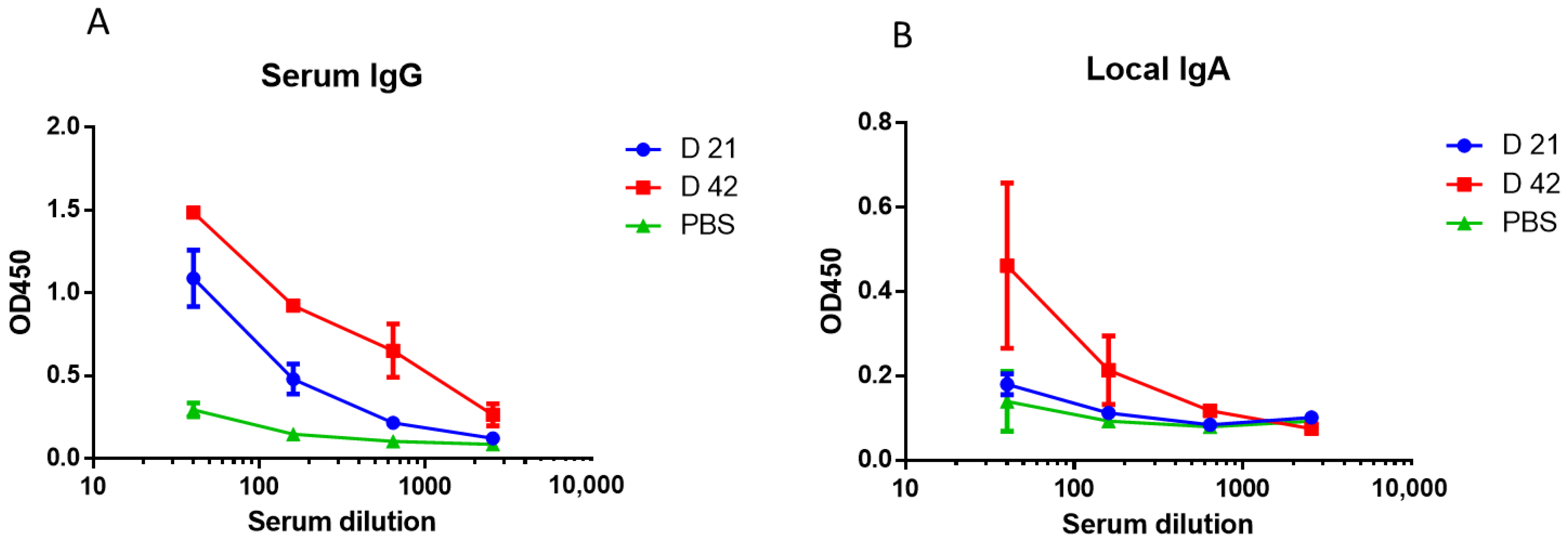
2.5. Immunogenicity
2.6. Early Protection against Influenza Infection
2.7. Early Cytokine Production In Vitro
2.8. Post-Influenza Pneumococcal Pneumonia in Mice
2.9. Statistical Analysis
3. Results
3.1. Study of Early Protection against Homologous and Heterologous Influenza Infections after A/Mallard Netherlands/12/2000(H7N3) LAIV Immunization
3.2. Early Cytokine Production in THP-1 Cell Line
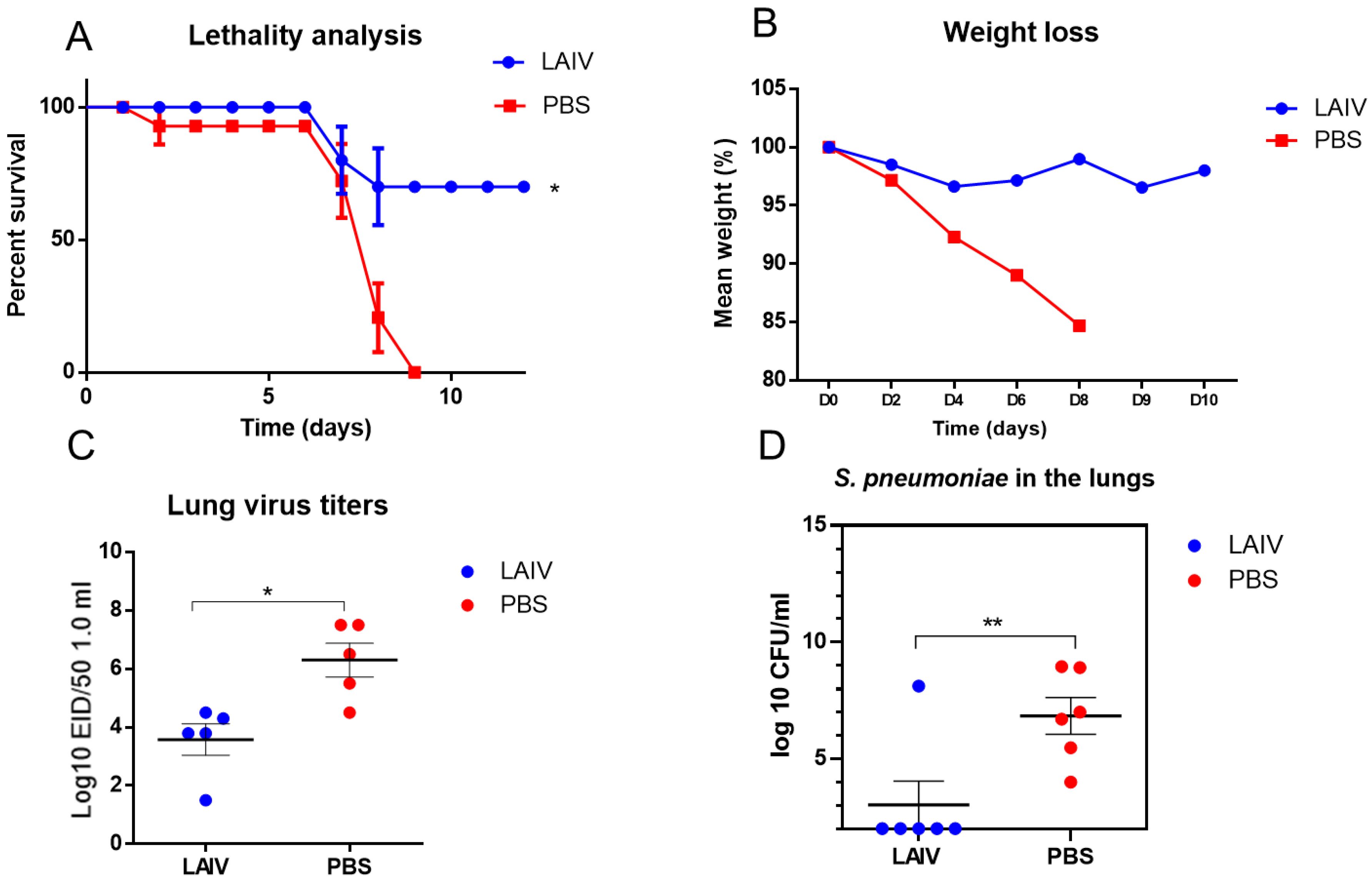
3.3. Evaluation of the Pneumococcal Invasion following Influenza Infection
3.4. The Effect of Immunization with LAIV on the Resistance to Secondary Post-Influenza Pneumococcal Pneumonia
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rudenko, L.; Isakova-Sivak, I. Pandemic preparedness with live attenuated influenza vaccines based on A/Leningrad/134/17/57 master donor virus. Expert Rev. Vaccines 2015, 14, 395–412. [Google Scholar] [CrossRef]
- Pebody, R.; McMenamin, J.; Nohynek, H. Live attenuated influenza vaccine (LAIV): Recent effectiveness results from the USA and implications for LAIV programmes elsewhere. Arch. Dis. Child. 2018, 103, 101–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMichael, A.J.; Gotch, F.M.; Noble, G.R.; Beare, P.A. Cytotoxic T-cell immunity to influenza. N. Engl. J. Med. 1983, 309, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.J.; Hung, M.C.; O’Halloran, A.C.; Ding, H.; Srivastav, A.; Williams, W.W.; Singleton, J.A. Seasonal influenza vaccination coverage trends among adult populations, US, 2010–2016. Am. J. Prev. Med. 2019, 57, 458–469. [Google Scholar] [CrossRef] [PubMed]
- Grohskopf, L.A.; Alyanak, E.; Broder, K.R.; Walter, E.B.; Fry, A.M.; Jernigan, D.B. Prevention and control of seasonal influenza with vaccines: Recommendations of the Advisory Committee on Immunization Practices—United States, 2019–2020 influenza season. MMWR Recomm. Rep. 2019, 68, 1. [Google Scholar] [CrossRef] [Green Version]
- Grohskopf, L.A.; Sokolow, L.Z.; Fry, A.M.; Walter, E.B.; Jernigan, D.B. Update: ACIP recommendations for the use of quadrivalent live attenuated influenza vaccine (LAIV4)—United States, 2018–2019 influenza season. Morb. Mortal. Wkly. Rep. 2018, 67, 643. [Google Scholar] [CrossRef]
- Hoft, D.F.; Lottenbach, K.R.; Blazevic, A.; Turan, A.; Blevins, T.P.; Pacatte, T.P.; Yu, Y.; Mitchell, M.C.; Hoft, S.G.; Belshe, R.B. Comparisons of the humoral and cellular immune responses induced by live attenuated influenza vaccine and inactivated influenza vaccine in adults. Clin. Vaccine Immunol. 2017, 24, e00414-16. [Google Scholar] [CrossRef] [Green Version]
- McCullers, J.A.; McAuley, J.L.; Browall, S.; Iverson, A.R.; Boyd, K.L.; Henriques Normark, B. Influenza enhances susceptibility to natural acquisition of and disease due to Streptococcus pneumoniae in ferrets. J. Infect. Dis. 2010, 202, 1287–1295. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.H.; Meng, F.; Seitz, M.; Valentin-Weigand, P.; Herrler, G. Sialic acid-dependent interactions between influenza viruses and Streptococcus suis affect the infection of porcine tracheal cells. J. Gen. Virol. 2015, 96, 2557–2568. [Google Scholar] [CrossRef]
- Morens, D.M.; Taubenberger, J.K.; Fauci, A.S. Predominant role of bacterial pneumonia as a cause of death in pandemic influenza: Implications for pandemic influenza preparedness. J. Infect. Dis. 2008, 198, 962–970. [Google Scholar] [CrossRef]
- Nelson, G.E.; Gershman, K.A.; Swerdlow, D.L.; Beall, B.W.; Moore, M.R. Invasive pneumococcal disease and pandemic (H1N1) 2009, Denver, Colorado, USA. Emerg. Infect. Dis. 2012, 18, 208. [Google Scholar] [CrossRef] [PubMed]
- Klausberger, M.; Leneva, I.A.; Falynskova, I.N.; Vasiliev, K.; Poddubikov, A.V.; Lindner, C.; Kartaschova, N.P.; Svitich, O.A.; Stukova, M.; Grabherr, R.; et al. The potential of influenza HA-specific immunity in mitigating lethality of postinfluenza pneumococcal infections. Vaccines 2019, 7, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belshe, R.B.; Gruber, W.C. Prevention of otitis media in children with live attenuated influenza vaccine given intranasally. Pediatr. Infect. Dis. J. 2000, 19, S66–S71. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Lee, J.Y.; Jang, Y.H.; Seo, S.U.; Chang, J.; Seong, B.L. Non-specific effect of vaccines: Immediate protection against respiratory syncytial virus infection by a live attenuated influenza vaccine. Front. Microbiol. 2018, 9, 83. [Google Scholar] [CrossRef]
- Lu, X.; Edwards, L.E.; Desheva, J.A.; Nguyen, D.C.; Rekstin, A.; Stephenson, I.; Szretter, K.; Cox, N.J.; Rudenko, L.G.; Klimov, A.; et al. Cross-protective immunity in mice induced by live-attenuated or inactivated vaccines against highly pathogenic influenza A (H5N1) viruses. Vaccine 2006, 24, 6588–6593. [Google Scholar] [CrossRef]
- Kramskaya, T.; Leontieva, G.; Desheva, Y.; Grabovskaya, K.; Gupalova, T.; Rudenko, L.; Suvorov, A. Combined immunization with attenuated live influenza vaccine and chimeric pneumococcal recombinant protein improves the outcome of virus-bacterial infection in mice. PLoS ONE 2019, 14, e0222148. [Google Scholar]
- Desheva, Y.A.; Leontieva, G.F.; Kramskaya, T.A.; Smolonogina, T.A.; Grabovskaya, K.B.; Kiseleva, I.V.; Rudenko, L.G.; Suvorov, A.N. Evaluation in mouse model of combined virus-bacterial vaccine based on attenuated influenza A (H7N3) virus and the group B streptococcus recombinant polypeptides. Open Microbiol. J. 2016, 10, 168. [Google Scholar] [CrossRef] [Green Version]
- Kiseleva, I.; Rekstin, A.; Al Farroukh, M.; Bazhenova, E.; Katelnikova, A.; Puchkova, L.; Rudenko, L. Non-mouse-adapted H1N1pdm09 virus as a model for influenza research. Viruses 2020, 12, 590. [Google Scholar] [CrossRef]
- Landgraf, G.; Desheva, Y.A.; Rudenko, L.G. Evaluation of influenza A and B cold-adapted reassortant virus reproduction in trivalent live influenza vaccines. Virus Res. 2021, 300, 198396. [Google Scholar] [CrossRef]
- Rudenko, L.G. Live attenuated cold-adapted influenza vaccine in Russia: Advantages, further research and development. In Options for the Control of Influenza VI; International Medical Press Ltd.: London, UK; Atlanta, GA, USA, 2008; pp. 122–124. [Google Scholar]
- Bai, L.; Zhao, Y.; Dong, J.; Liang, S.; Guo, M.; Liu, X.; Wang, X.; Huang, Z.; Sun, X.; Zhang, Z.; et al. Coinfection with influenza A virus enhances SARS-CoV-2 infectivity. Cell Res. 2021, 31, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, L.G.; Arden, N.H.; Grigorieva, E.; Naychin, A.; Rekstin, A.; Klimov, A.I.; Donina, S.; Desheva, J.; Holman, R.C.; DeGuzman, A.; et al. Immunogenicity and efficacy of Russian live attenuated and US inactivated influenza vaccines used alone and in combination in nursing home residents. Vaccine 2000, 19, 308–318. [Google Scholar] [PubMed]
- Grigor’eva, E.P.; Desheva IuA, S.A.; Donina, A.N.; Naĭkhin, A.R.; Rekstin, I.B.; Barantseva, E.A.; Zavitkova, T.M.; Moskvicheva, V.G.; Zhavoronkov, L.; Rudenko, G. The comparative characteristics of the safety, immunogenic activity and prophylactic potency of the adult and children types of live influenza vaccine in schoolchildren aged 7–14 years. Vopr Virusol. 2002, 47, 24–27. [Google Scholar]
- Rathnasinghe, R.; Salvatore, M.; Zheng, H.; Jangra, S.; Kehrer, T.; Mena, I.; Schotsaert, M.; Muster, T.; Palese, P.; García-Sastre, A. Interferon mediated prophylactic protection against respiratory viruses conferred by a prototype live attenuated influenza virus vaccine lacking non-structural protein 1. Sci. Rep. 2021, 11, 22164. [Google Scholar] [CrossRef] [PubMed]
- Desheva, Y.A.; Leontieva, G.F.; Kramskaya, T.A.; Landgraf, G.O.; Sychev, I.A.; Rekstin, A.R.; Suvorov, A.N. Factors of early protective action of live influenza vaccine combined with recombinant bacterial polypeptides against homologous and heterologous influenza infection. Heliyon 2019, 5, e01154. [Google Scholar]
- McNab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O’Garra, A. Type I interferons in infectious disease. Nat. Rev. 2015, 15, 87–103. [Google Scholar]
- Hopkins, S.J. The pathophysiological role of cytokines. Leg. Med. 2003, 5, S45–S57. [Google Scholar]
- Feghali, C.A.; Wright, T.M. Cytokines in acute and chronic inflammation. Front. Biosci. 1997, 2, d12–d26. [Google Scholar]
- Rubino, G.; Bulati, M.; Aiello, A.; Aprile, S.; Gambino, C.M.; Gervasi, F.; Caruso, C.; Accardi, G. Sicilian centenarian offspring are more resistant to immune ageing. Aging Clin. Exp. Res. 2019, 31, 125–133. [Google Scholar]
- Sridhar, S.; Begom, S.; Hoschler, K.; Bermingham, A.; Adamson, W.; Carman, W.; Riley, S.; Lalvani, A. Longevity and determinants of protective humoral immunity after pandemic influenza infection. Am. J. Respir. Crit. Care Med. 2015, 191, 325–332. [Google Scholar]
- Poland, G.A. Influenza vaccine failure: Failure to protect or failure to understand? Expert Rev. Vaccines 2018, 17, 495–502. [Google Scholar] [CrossRef]
- Louie, J.; Jean, C.; Chen, T.H.; Park, S.; Ueki, R.; Harper, T.; Blau, D.M. Bacterial coinfections in lung tissue specimens from fatal cases of 2009 pandemic influenza A (H1N1)-United States, May–August 2009. Morb. Mortal. Wkly. Rep. 2009, 58, 1071–1074. [Google Scholar]
- Haynes, L.; Szaba, F.M.; Eaton, S.M.; Kummer, L.W.; Lanthier, P.A.; Petell, A.H.; Smiley, S.T. Immunity to the conserved influenza nucleoprotein reduces susceptibility to secondary bacterial infections. J. Immunol. 2012, 189, 4921–4929. [Google Scholar] [CrossRef] [Green Version]
- McCullers, J.A.; Huber, V.C. Correlates of vaccine protection from influenza and its complications. Hum. Vaccines Immunother. 2012, 8, 34–44. [Google Scholar] [CrossRef]
- Makhmudova, N.R.; Leneva, I.A.; Larionova, N.V.; Poddubikov, A.V.; Falynskova, I.N.; Kartashova, N.P.; Svitich, O.A. The safety of attenuated and recombinant nasal influenza vaccines in terms of the development of secondary bacterial superinfection. Z. Mikrobiol. Epidemiol. Immunobiol. 2019, 6, 30–39. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccine Strain | Subtype/Lineage | ‘Wild-Type’ Parent Virus |
|---|---|---|
| A/17/Mallard Netherlands/00/95 | (H7N3) | A/Mallard Netherlands/12/2000 |
| A/17/South Africa/2013/01 | (H1N1)pdm09 | A/South Africa/3626/2013 |
| A/17/New York/15/5364 | A/17/New York/15/5364 | A/New York/61/15 |
| A/17/Hong Kong/14/8296 | (H3N2) | Hong Kong/4801/14 |
| B/60/Brisbane/08/83 | B/Victoria | Brisbane/60/08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Desheva, Y.; Leontieva, G.; Kramskaya, T.; Losev, I.; Rekstin, A.; Petkova, N.; Kudar, P.; Suvorov, A. Live Influenza Vaccine Provides Early Protection against Homologous and Heterologous Influenza and May Prevent Post-Influenza Pneumococcal Infections in Mice. Microorganisms 2022, 10, 1150. https://doi.org/10.3390/microorganisms10061150
Desheva Y, Leontieva G, Kramskaya T, Losev I, Rekstin A, Petkova N, Kudar P, Suvorov A. Live Influenza Vaccine Provides Early Protection against Homologous and Heterologous Influenza and May Prevent Post-Influenza Pneumococcal Infections in Mice. Microorganisms. 2022; 10(6):1150. https://doi.org/10.3390/microorganisms10061150
Chicago/Turabian StyleDesheva, Yulia, Galina Leontieva, Tatiana Kramskaya, Igor Losev, Andrey Rekstin, Nadezhda Petkova, Polina Kudar, and Alexander Suvorov. 2022. "Live Influenza Vaccine Provides Early Protection against Homologous and Heterologous Influenza and May Prevent Post-Influenza Pneumococcal Infections in Mice" Microorganisms 10, no. 6: 1150. https://doi.org/10.3390/microorganisms10061150
APA StyleDesheva, Y., Leontieva, G., Kramskaya, T., Losev, I., Rekstin, A., Petkova, N., Kudar, P., & Suvorov, A. (2022). Live Influenza Vaccine Provides Early Protection against Homologous and Heterologous Influenza and May Prevent Post-Influenza Pneumococcal Infections in Mice. Microorganisms, 10(6), 1150. https://doi.org/10.3390/microorganisms10061150