
Complete Genome Analysis of Rhodococcus opacus S8 Capable of Degrading Alkanes and Producing Biosurfactant Reveals Its Genetic Adaptation for Crude Oil Decomposition

 , ,
, , 
Abstract
:1. Introduction
2. Material and Methods
2.1. Bacterial Strains and Cultivation Conditions
2.2. Determination of the Surface Tension
2.3. Whole Genome Sequencing
2.4. Estimation of Expression of Target Genes
2.5. Isolation of Biosurfactants
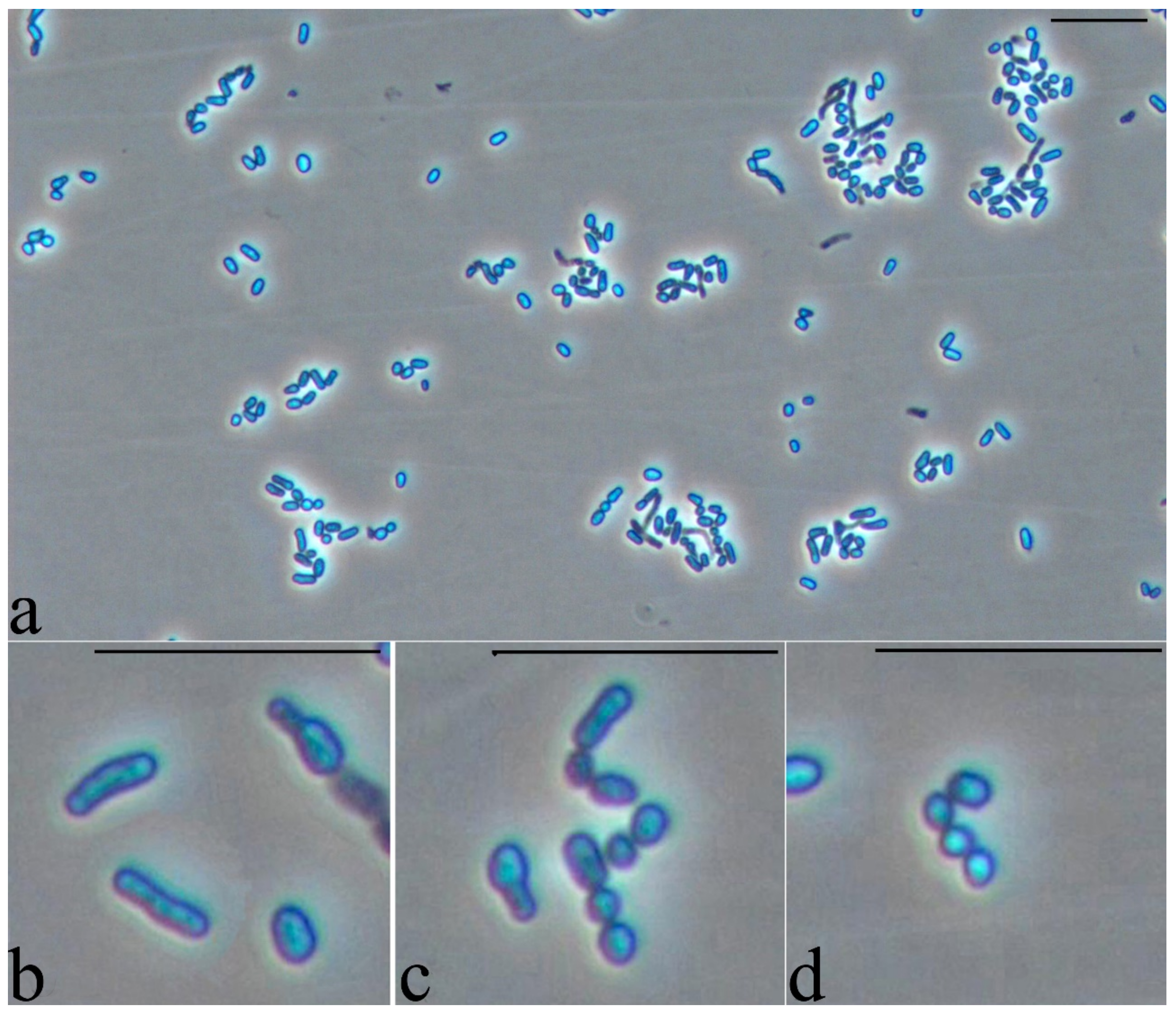
2.6. Microscopy
2.6.1. Phase Contrast Microscopy
2.6.2. Electron Microscopy
3. Results and Discussion
3.1. Surface Tension
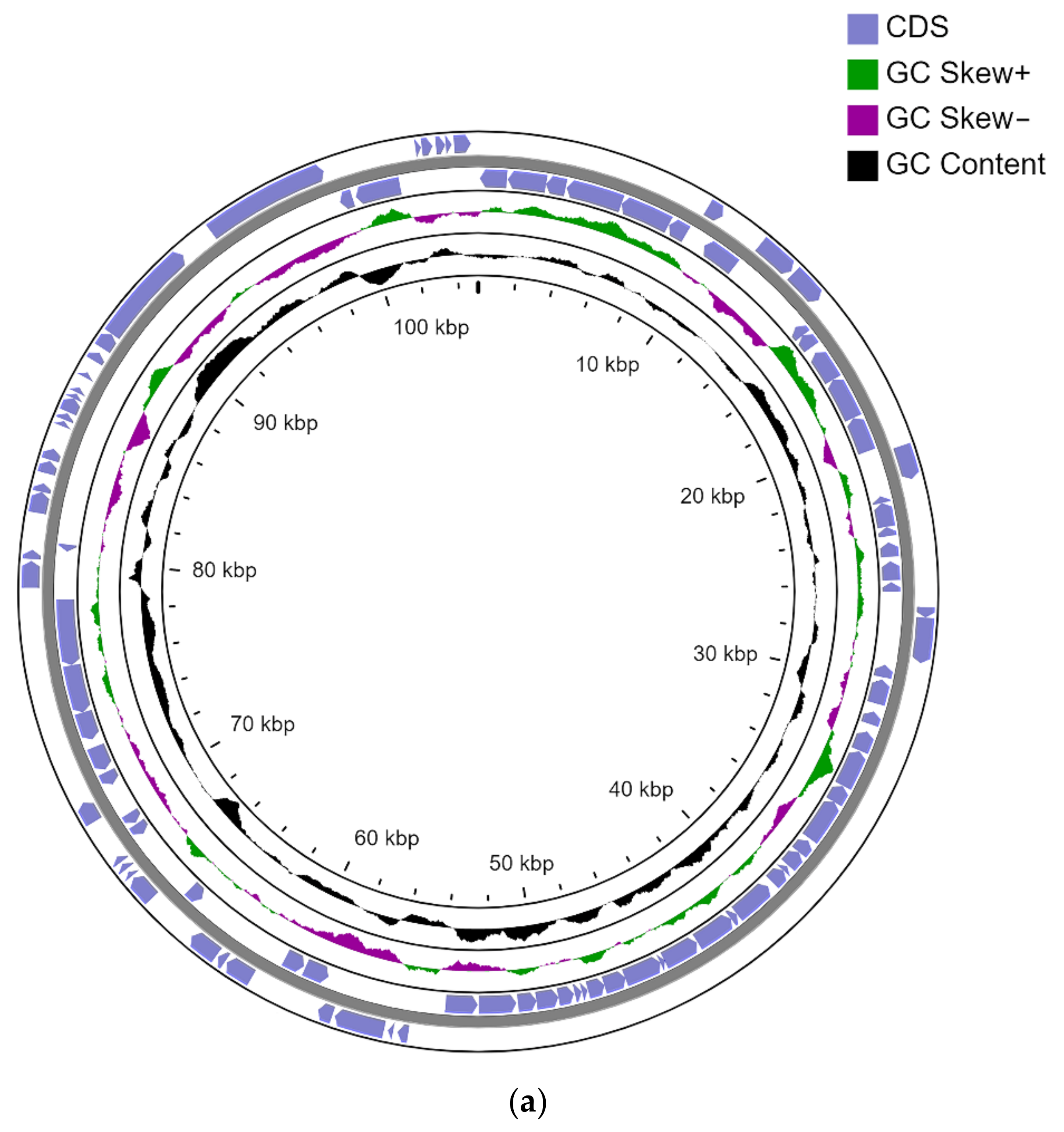
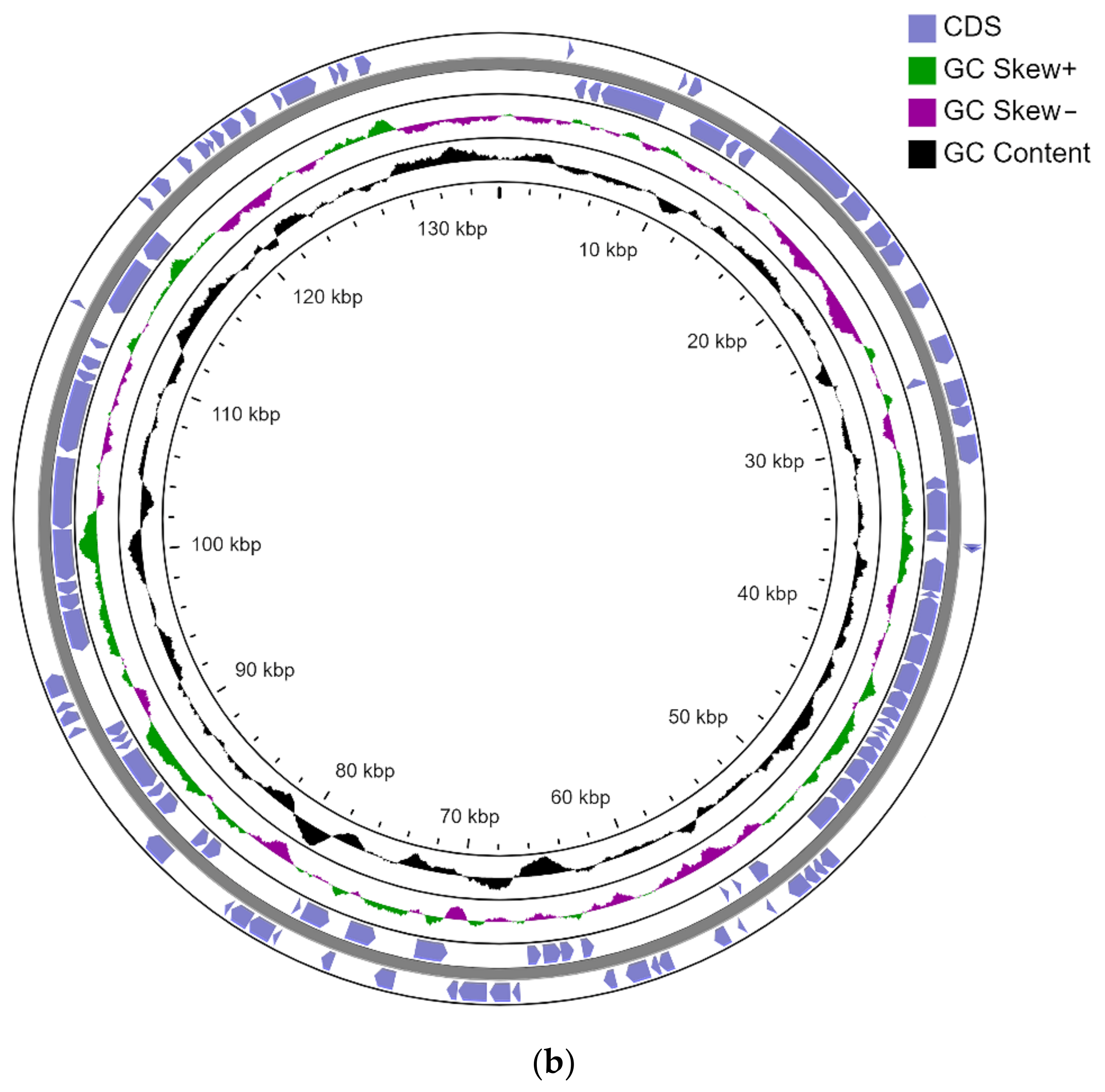
3.2. Genome Sequencing
- Chromosome-CP093380
- pLPS8-CP093381
- pCP105-CP093383
- pCP135-CP093382 (Figure 1).
- naphthalene catabolism gene cluster
- catechol 2,3-dioxygenase (EC 1.13.11.2)
- phthalate catabolism genes
- 3 copies of cytochrome P450 hydroxylase
- The presence of truncated hemoglobin genes in the genome of the studied R. opacus S8 should be noted. The strain R. opacus S8 has 2 genes located on the chromosome. On the contrary, similar genes were not found in the genome of the R. qingshengii strain VT6.
- Trehalose 2-sulfotransferase
- Trehalose-2-sulfate acyltransferase papA2 (2 copies)
- Acyltransferase papA3.
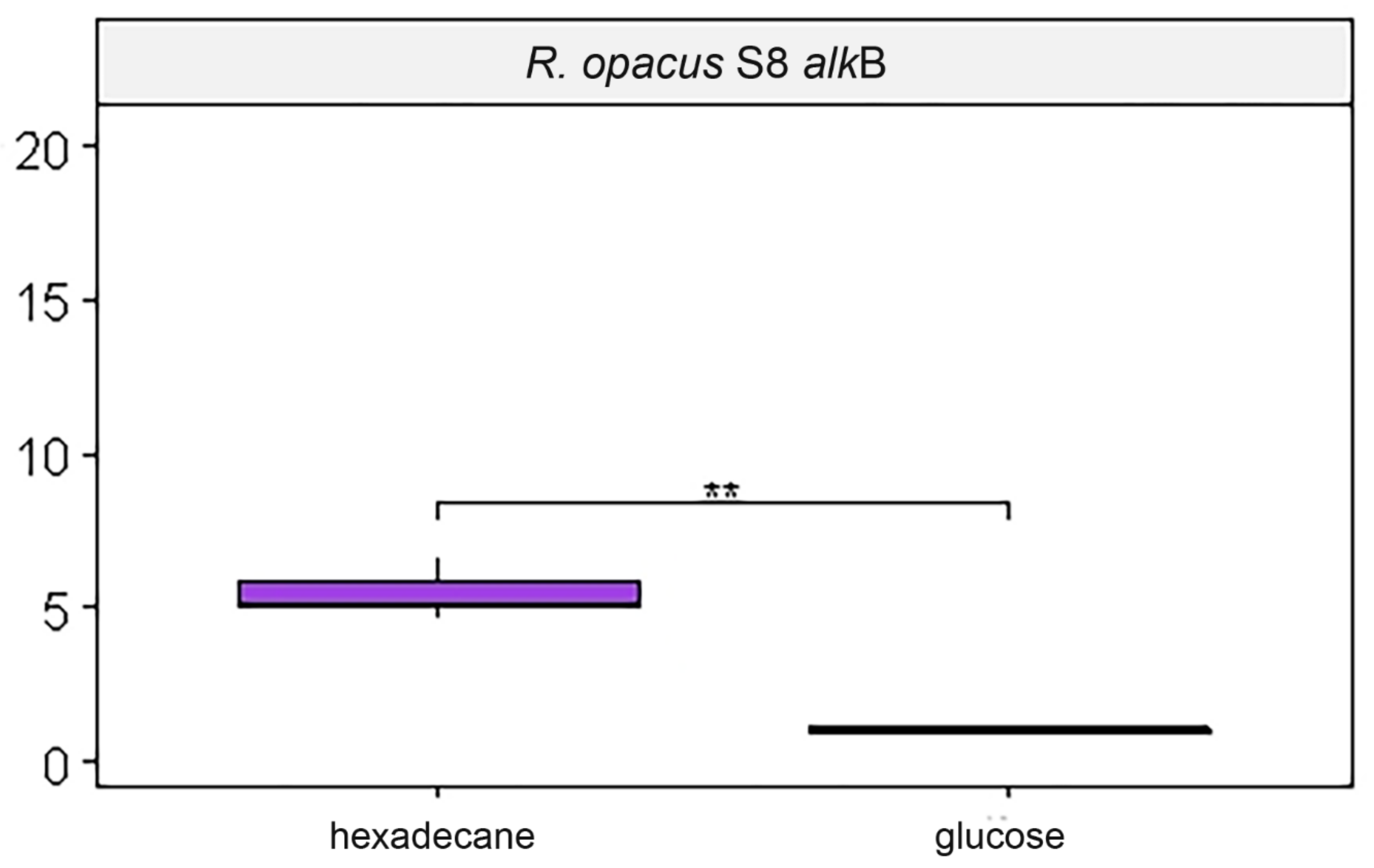
3.3. Estimation of Expression of Target Genes
3.4. Biosurfactants
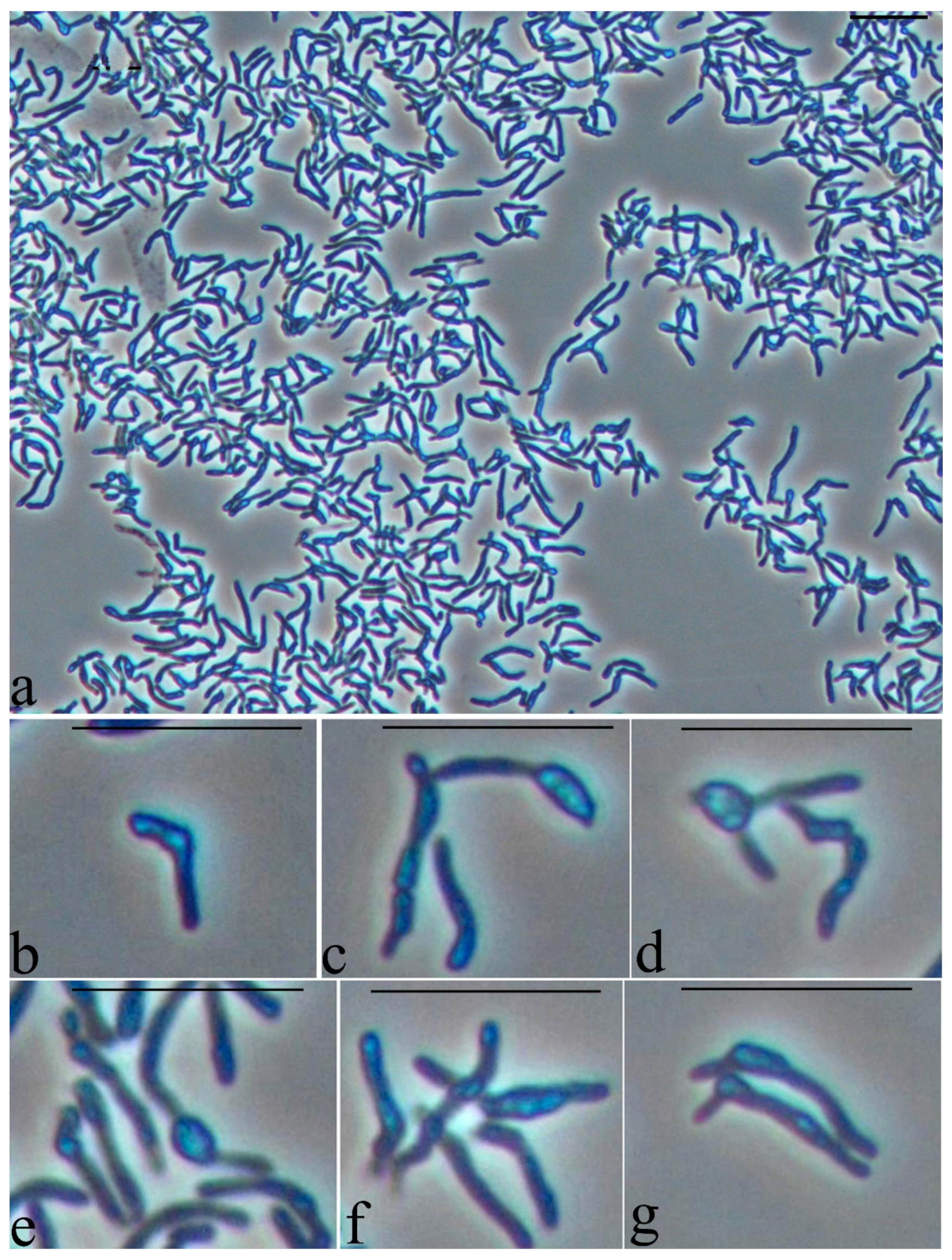
3.5. Study of the Cell Morphology when Interacting with a Hydrophobic Substrate
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vogt, C.; Richnow, H.H. Bioremediation via in situ microbial degradation of organic pollutants. Adv. Biochem. Eng. Biotechnol. 2014, 142, 123–146. [Google Scholar] [CrossRef]
- Zargar, A.N.; Kumar, A.; Sinha, A.; Kumar, A.; Skiadas, I.; Mishra, S.; Srivastava, P. Asphaltene biotransformation for heavy oil upgradation. AMB Expr. 2021, 11, 127. [Google Scholar] [CrossRef] [PubMed]
- Trevors, J.T.; Saier, M.H. The legacy of oil spills. Water Air Soil Pollut. 2010, 211, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yergeau, E.; Sanschagrin, S.; Beaumier, D.; Greer, C.W. Metagenomic analysis of the bioremediation of diesel-contaminated canadian high Arctic soils. PLoS ONE 2012, 7, e30058. [Google Scholar] [CrossRef]
- Roslee, A.F.A.; Zakaria, N.N.; Convey, P.; Zulkharnain, A.; Lee, G.L.Y.; Gomez-Fuentes, C.; Ahmad, S.A. Statistical optimisation of growth conditions and diesel degradation by the Antarctic bacterium, Rhodococcus sp. strain AQ5-07. Extremophiles 2020, 24, 277–291. [Google Scholar] [CrossRef] [PubMed]
- Bell, T.H.; Yergeau, E.; Maynard, C.; Juck, D.; Whyte, L.G.; Greer, C.W. Predictable bacterial composition and hydrocarbon degradation in Arctic soils following diesel and nutrient disturbance. ISME J. 2013, 7, 1200–1210. [Google Scholar] [CrossRef]
- Al-Mailem, D.M.; Al-Deieg, M.; Eliyas, M.; Radwan, S.S. Biostimulation of indigenous microorganisms for bioremediation of oily hypersaline microcosms from the Arabian Gulf Kuwaiti coasts. J. Environ. Manag. 2017, 193, 576–583. [Google Scholar] [CrossRef]
- Perfumo, A.; Smyth, T.; Marchant, R.; Banat, I.M. Production and roles of biosurfactants and bioemulsifiers in accessing hydrophobic substrates. In Handbook of Hydrocarbon and Lipid Microbiology; Timmis, K.N., Ed.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 1502–1512. [Google Scholar] [CrossRef]
- Shahi, A.; Ince, B.; Aydin, S.; Ince, O. Assessment of the horizontal transfer of functional genes as a suitable approach for evaluation of the bioremediation potential of petroleum-contaminated sites: A mini-review. Appl. Microbiol. Biotechnol. 2017, 101, 4341–4348. [Google Scholar] [CrossRef] [PubMed]
- Chettri, B.; Singha, N.A.; Mukherjee, A.; Rai, A.N.; Chattopadhyay, D.; Singh, A.K. Hydrocarbon degradation potential and competitive persistence of hydrocarbonoclastic bacterium Acinetobacter pittii strain ABC. Arch. Microbiol. 2019, 201, 1129–1140. [Google Scholar] [CrossRef]
- Das, D.; Baruah, R.; Roy, A.S.; Singh, A.K.; Boruah, H.P.D.; Kalita, J.; Bora, T.C. Complete genome sequence analysis of Pseudomonas aeruginosa N002 reveals its genetic adaptation for crude oil degradation. Genomics 2015, 105, 182–190. [Google Scholar] [CrossRef]
- Van Hamme, J.D.; Singh, A.; Ward, O.P. Recent advances in petroleum microbiology. Microbiol. Mol. Biol. Rev. 2003, 67, 503–549. [Google Scholar] [CrossRef] [Green Version]
- Franzetti, A.; Tamburini, E.; Banat, I.M. Applications of biological surface active compounds in remediation technologies. In Advances in Experimental Medicine and Biology; Sen, R., Ed.; Springer: New York, NY, USA, 2010; Volume 672. [Google Scholar] [CrossRef]
- Nisenbaum, M.; Corti-Monzón, G.; Villegas-Plazas, M.; Junca, H.; Mangani, A.; Patat, M.L.; González, J.F.; Murialdo, S.E. Enrichment and key features of a robust and consistent indigenous marine-cognate microbial consortium growing on oily bilge wastewaters. Biodegradation 2020, 31, 91–108. [Google Scholar] [CrossRef]
- Margesin, R.; Moertelmaier, C.; Mair, J. Low-temperature biodegradation of petroleum hydrocarbons (n-alkanes, phenol, anthracene, pyrene) by four actinobacterial strains. Intern. Biodeter. Biodegrad. 2012, 84, 185–191. [Google Scholar] [CrossRef]
- Van Hong, T.P.; Chaudhary, D.; Jeong, S.W.; Jaisoo, K. Oil-degrading properties of a psychrotolerant bacterial strain, Rhodococcus sp. Y2-2, in liquid and soil media. World J. Microbiol. Biotechnol. 2018, 34, 33. [Google Scholar] [CrossRef] [PubMed]
- De Coster, W.; D’Hert, S.; Schultz, D.T.; Cruts, M.; Van Broeckhoven, C. NanoPack: Visualizing and processing long-read sequencing data. Bioinformatics 2018, 34, 2666–2669. [Google Scholar] [CrossRef]
- Nurk, S.; Bankevich, A.; Antipov, D.; Pevzner, P.A. Assembling genomes and mini-metagenomes from highly chimeric reads. In Proceedings of the 17th International Conference on Research in Computational Molecular Biology, Beijing, China, 7–10 April 2013. [Google Scholar] [CrossRef]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of long, error-prone reads using repeat graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Milne, I.; Stephen, G.; Bayer, M.; Cock, P.J.A.; Pritchard, L.; Cardle, L.; Shaw, P.D.; Marshall, D. Using Tablet for visual exploration of second-generation sequencing data. Brief. Bioinform. 2013, 14, 193–202. [Google Scholar] [CrossRef]
- Walter, V.; Syldatk, C.; Hausmann, R. Screening concepts for the isolation of biosurfactant producing microorganisms. In Madame Curie Bioscience Database; Sen, R., Ed.; Landes Bioscience: Austin, TX, USA, 2010; Volume 672, Chapter 1; pp. 1–13. [Google Scholar] [CrossRef]
- Solyanikova, I.P.; Suzina, N.E.; Emelyanova, E.V.; Polivtseva, V.N.; Pshenichnikova, A.B.; Lobanok, A.G.; Golovleva, L.A. Morphological, physiological, and biochemical characteristics of a benzoate-degrading strain Rhodococcus opacus 1CP under stress conditions. Microbiology 2017, 86, 202–212. [Google Scholar] [CrossRef]
- Van Hamme, J.D.; Singh, A.; Ward, O.P. Physiological Aspects. Part 1 in a series of papers devoted to surfactants in microbiology and technology. Biotechnol. Adv. 2006, 24, 604–620. [Google Scholar] [CrossRef] [PubMed]
- Bodour, A.A.; Drees, K.P.; Maier, R.M. Distribution of biosurfactant-producing bacteria in undisturbed and contaminated arid southwestern soils. Appl. Environm. Microbiol. 2003, 69, 3280–3287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, N.P.; Landfald, B.; Willassen, N.P. Biological surface-active compounds from marine bacteria. Environ. Technol. 2015, 37, 1151–1158. [Google Scholar] [CrossRef] [PubMed]
- Peng, F.; Wang, Y.; Sun, F.; Liu, Z.; Lai, Q.; Shao, Z. A novel lipopeptide produced by a Pacific Ocean deep-sea bacterium, Rhodococcus sp. TW53. J. Appl. Microbiol. 2008, 105, 698–705. [Google Scholar] [CrossRef]
- Delegan, Y.; Petrikov, K.; Frantsuzova, E.; Bogun, A.; Travkin, V.; Solyanikova, I. Complete genome sequence of Rhodococcus qingshengii VT6, a promising degrader of persistent pollutants and putative biosurfactant producing strain. Microbiol. Resour. Announc. 2022, 11, e01179-21. [Google Scholar] [CrossRef]
- Tazzyman, S.J.; Bonhoeffer, S. Why there are no essential genes on plasmids. Mol. Biol. Evol. 2015, 32, 3079–3088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markande, A.R.; Patel, D.; Varjani, S. A review on biosurfactants: Properties, applications and current developments. Bioresour. Technol. 2021, 330, 124963. [Google Scholar] [CrossRef] [PubMed]
- Quatrini, P.; Scaglione, G.; De Pasquale, C.; Riela, S.; Puglia, A.M. Isolation of Gram-positive n-alkane degraders from a hydrocarbon-contaminated Mediterranean shoreline. J. Appl. Microbiol. 2008, 104, 251–259. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, S.H.; Yoon, J.H.; Lee, P.C. Carotenoid production from n-alkanes with a broad range of chain lengths by the novel species Gordonia ajoucoccus A2(T). Appl. Microbiol. Biotechnol. 2014, 98, 3759–3768. [Google Scholar] [CrossRef]
- Laczi, K.; Kis, Á.; Horváth, B.; Maroti, G.; Hegedus, B.; Perei, K.; Rakhely, G. Metabolic responses of Rhodococcus erythropolis PR4 grown on diesel oil and various hydrocarbons. Appl. Microbiol. Biotechnol. 2015, 99, 9745–9759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, B.; Bouriat, P.; Goulas, F.; Grimaud, R. Behavior of Marinobacter hydrocarbonoclasticus SP17 cells during initiation of biofilm formation at the alkane–water interface. Biotechnol. Bioeng. 2010, 105, 461–468. [Google Scholar] [CrossRef]
- Pepi, M.; Cesaro, A.; Liut, G.; Baldi, F. An antarctic psychrotrophic bacterium Halomonas sp. ANT-3b, growing on n-hexadecane, produces a new emulsyfying glycolipid. FEMS Microbiol. Ecol. 2005, 53, 157–166. [Google Scholar] [CrossRef]
- Cameotra, S.S.; Singh, P. Synthesis of rhamnolipid biosurfactant and mode of hexadecane uptake by Pseudomonas species. Microb. Cell Factories 2009, 8, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, X.; Wu, Z.; Zhang, H.; Lu, D.; Wang, M.; Liu, Y.; Liu, Z. Degradation of hexadecane by Enterobacter cloacae strain TU that secretes an exopolysaccharide as a bioemulsifier. Chemosphere 2010, 80, 951–956. [Google Scholar] [CrossRef]
- Duda, V.I.; Dmitriev, V.V.; Suzina, N.E.; Shorokhova, A.P.; Mishina, G.V. Ultrastructural organization of gas balloons and surface films in colonies of gram-negative bacteria Alcaligenes sp. strain d2. Microbiology 1996, 65, 195–200. [Google Scholar]
- Duda, V.I.; Lebedinsky, A.V.; Dmitriev, V.V.; Suzina, N.E.; Shorokhova, A.P.; Mityushina, L.L. Formation of extracellular gas balloons in the colonies of some Gram-negative bacteria. Microbiology 1999, 68, 69–76. [Google Scholar]
- Wainwright, L.M.; Wang, Y.; Park, S.F.; Yeh, S.-R.; Poole, R.K. Purification and spectroscopic characterization of Ctb, a group III truncated hemoglobin implicated in oxygen metabolism in the food-borne pathogen Campylobacter jejuni. Biochemistry 2006, 45, 6003–6011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duda, V.I.; Ilchenko, A.P.; Dmitriev, V.V.; Shorokhova, A.P.; Suzina, N.E. Isolation and characterization of hemoflavoprotein from the gram-negative bacterium Alcaligenes sp., strain d2. Microbiology 1998, 67, 7–12. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primers | Primer Sequence | Amplicon Size, bp |
|---|---|---|---|
| 16S rRNA | s8_16s_rt_for | GGACGAAGCGAAAGTGACG | 130 |
| s8_16s_rt_rev | CGACAAACCGCCTACGA | ||
| Alkane 1-Monooxygenase 1 | s8_alkb2884_rt_for | CATCAACACCGCACACGAGA | 213 |
| s8_alkb2884_rt_rev | TTCAGGCTTCCCCAGACACT | ||
| Trehalose 2-Sulfotransferase | s8_tst1278_rt_for | TACCTGCCGTCCACCAATC | 183 |
| s8_tst1278_rt_rev | CTTTCCGCCCCACACCC | ||
| Trehalose-2-Sulfate Acyltransferase PapA2 | s8_tsat7567_rt_for | GAAGCGGACCTGACGAAAGA | 150 |
| s8_tsat7567_rt_rev | CGAGAAGGGGGCGGACA |
| Strain | ST, mN/m |
|---|---|
| Rhodococcus sp. Ac2845 | 69 ± 1.1 |
| Rhodococcus sp. P13 | 47 ± 1.2 |
| Rhodococcus sp. P1 | 48 ± 1.1 |
| Rhodococcus sp. G172 | 49 ± 1.3 |
| Rhodococcus sp. 557 | 65 ± 1.6 |
| Rhodococcus opacus S8 | 39 ± 0.8 |
| Rhodococcus ruber P25 | 61 ± 1.4 |
| Rhodococcus sp. P12 | 47 ± 1.2 |
| Rhodococcus sp. VT7 | 52 ± 0.9 |
| Rhodococcus wratislaviensis G10 | 52 ± 1.0 |
| Rhodococcus sp. 412 | 54 ± 1.3 |
| Rhodococcus qingshengii VT6 | 32 ± 0.7 |
| Strain | Genome Size, Mb | Chromosome Size, Mb | Plasmid Size, Mb |
|---|---|---|---|
| S8 | 8.78 | 7.75 | 788 135 105 |
| R7 (GCA_000736435.1) | 10.12 | 8.46 | 656 426 352 191 25 |
| XM24D (GCA_012931745.1) | 9.08 | 7.85, 1.23 | - |
| KT112-7 (GCA_017916235.1) | 8.00 | 7.58 | 282 131 |
| 1CP (GCA_001685605.1) | 8.64 | 7.68 | 64 885 |
| B4 (GCA_000010805.1) | 8.83 | 7.91 | 111 4 558 2.7 245 |
| PD630 (GCA_020542785.1) | 9.17 | 8.37 | 538 168 85 |
| DSM44186 (GCA_019856255.1) | 8.84 | 8.04 | 744 52 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delegan, Y.; Petrikov, K.; Frantsuzova, E.; Rudenko, N.; Solomentsev, V.; Suzina, N.; Travkin, V.; Solyanikova, I.P. Complete Genome Analysis of Rhodococcus opacus S8 Capable of Degrading Alkanes and Producing Biosurfactant Reveals Its Genetic Adaptation for Crude Oil Decomposition. Microorganisms 2022, 10, 1172. https://doi.org/10.3390/microorganisms10061172
Delegan Y, Petrikov K, Frantsuzova E, Rudenko N, Solomentsev V, Suzina N, Travkin V, Solyanikova IP. Complete Genome Analysis of Rhodococcus opacus S8 Capable of Degrading Alkanes and Producing Biosurfactant Reveals Its Genetic Adaptation for Crude Oil Decomposition. Microorganisms. 2022; 10(6):1172. https://doi.org/10.3390/microorganisms10061172
Chicago/Turabian StyleDelegan, Yanina, Kirill Petrikov, Ekaterina Frantsuzova, Natalia Rudenko, Viktor Solomentsev, Nataliya Suzina, Vasili Travkin, and Inna P. Solyanikova. 2022. "Complete Genome Analysis of Rhodococcus opacus S8 Capable of Degrading Alkanes and Producing Biosurfactant Reveals Its Genetic Adaptation for Crude Oil Decomposition" Microorganisms 10, no. 6: 1172. https://doi.org/10.3390/microorganisms10061172
APA StyleDelegan, Y., Petrikov, K., Frantsuzova, E., Rudenko, N., Solomentsev, V., Suzina, N., Travkin, V., & Solyanikova, I. P. (2022). Complete Genome Analysis of Rhodococcus opacus S8 Capable of Degrading Alkanes and Producing Biosurfactant Reveals Its Genetic Adaptation for Crude Oil Decomposition. Microorganisms, 10(6), 1172. https://doi.org/10.3390/microorganisms10061172