
Landscape Composition and Soil Physical–Chemical Properties Drive the Assemblages of Bacteria and Fungi in Conventional Vegetable Fields

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Landscape Analysis
2.2. Soil Sample Collection and Characterization
2.3. Determination of Soil Chemical Elements
2.4. Soil DNA Extraction, High-Throughput Sequencing, and Data Analysis
2.5. Statistical Analysis
3. Results
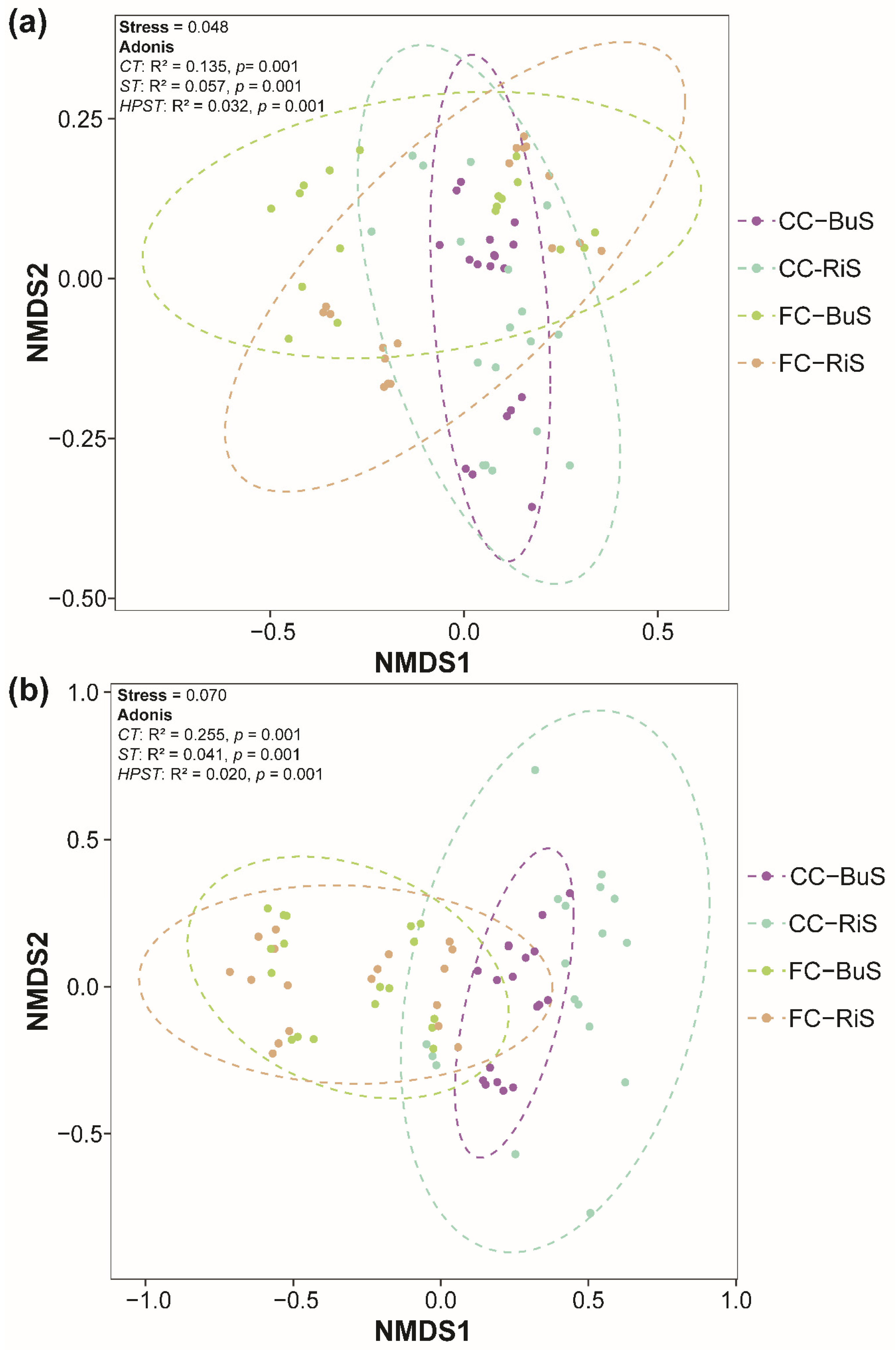
3.1. Soil Microbial Community Compositions
3.2. Variability of Soil Microbial Diversity in Different Local Field Factors
3.3. Integrative Analysis of the Factors Shaping the Soil Microbial Community
3.3.1. Effect of Different Factors on Shaping the Soil Bacterial Community
3.3.2. Effect of Different Factors on Shaping the Soil Fungal Community
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burges, A.; Alkorta, I.; Epelde, L.; Garbisu, C. From phytoremediation of soil contaminants to phytomanagement of ecosystem services in metal contaminated sites. Int. J. Phytoremediat. 2018, 20, 384–397. [Google Scholar] [CrossRef] [PubMed]
- Ngatia, L.W.; Moriasi, D.; Grace, J.M., III; Fu, R.; Gardner, C.S.; Taylor, R.W. Land Use Change Affects Soil Organic Carbon: An Indicator of Soil Health. In Environmental Health; IntechOpen: London, UK, 2021. [Google Scholar]
- Franklin, R.B.; Mills, A.L. Importance of spatially structured environmental heterogeneity in controlling microbial community composition at small spatial scales in an agricultural field. Soil Biol. Biochem. 2009, 41, 1833–1840. [Google Scholar] [CrossRef]
- Wang, H.; Marshall, C.W.; Cheng, M.; Xu, H.; Li, H.; Yang, X.; Zheng, T. Changes in land use driven by urbanization impact nitrogen cycling and the microbial community composition in soils. Sci. Rep. 2017, 7, 44049. [Google Scholar] [CrossRef] [PubMed]
- Mathew, R.P.; Feng, Y.; Githinji, L.J.M.; Ankumah, R.O.; Balkcom, K.S. Impact of No-Tillage and Conventional Tillage Systems on Soil Microbial Communities. Appl. Environ. Soil Sci. 2012, 2012, 548620. [Google Scholar] [CrossRef]
- Guo, X.; Chen, H.; Meng, M.; Biswas, S.; Ye, L.; Zhang, J. Effects of land use change on the composition of soil microbial communities in a managed subtropical forest. For. Ecol. Manag. 2016, 373, 93–99. [Google Scholar] [CrossRef]
- Thomson, B.C.; Tisserant, E.; Plassart, P.; Uroz, S.; Griffiths, R.; Hannula, S.E.; Buée, M.; Mougel, C.; Ranjard, L.; Van Veen, J.A.; et al. Soil conditions and land use intensification effects on soil microbial communities across a range of European field sites. Soil Biol. Biochem. 2015, 88, 403–413. [Google Scholar] [CrossRef]
- Fan, Z.; Lu, S.; Liu, S.; Guo, H.; Wang, T.; Zhou, J.; Peng, X. Changes in Plant Rhizosphere Microbial Communities under Different Vegetation Restoration Patterns in Karst and Non-karst Ecosystems. Sci. Rep. 2019, 9, 8761. [Google Scholar] [CrossRef] [PubMed]
- Steenwerth, K.L.; Jackson, L.E.; Calderón, F.J.; Stromberg, M.R.; Scow, K.M. Soil microbial community composition and land use history in cultivated and grassland ecosystems of coastal California. Soil Biol. Biochem. 2002, 34, 1599–1611. [Google Scholar] [CrossRef]
- Berg, G.; Smalla, K. Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol. Ecol. 2009, 68, 1–13. [Google Scholar] [CrossRef]
- Buée, M.; de Boer, W.; Martin, F.; Van Overbeek, L.; Jurkevitch, E. The rhizosphere zoo: An overview of plant-associated communities of microorganisms, including phages, bacteria, archaea, and fungi, and of some of their structuring factors. Plant Soil 2009, 321, 189–212. [Google Scholar] [CrossRef]
- Prashar, P.; Kapoor, N.; Sachdeva, S. Rhizosphere: Its structure, bacterial diversity and significance. Rev. Environ. Sci. Bio Technol. 2013, 13, 63–77. [Google Scholar] [CrossRef]
- York, L.M.; Carminati, A.; Mooney, S.J.; Ritz, K.; Bennett, M.J. The holistic rhizosphere: Integrating zones, processes, and semantics in the soil influenced by roots. J. Exp. Bot. 2016, 67, 3629–3643. [Google Scholar] [CrossRef] [PubMed]
- de la Porte, A.; Schmidt, R.; Yergeau, É.; Constant, P. A gaseous milieu: Extending the boundaries of the rhizosphere. Trends Microbiol. 2020, 28, 536–542. [Google Scholar] [CrossRef] [PubMed]
- Farrar, J.; Hawes, M.; Jones, D.; Lindow, S. How roots control the flux of carbon to the rhizosphere. Ecology 2003, 84, 827–837. [Google Scholar] [CrossRef]
- Razavi, B.S.; Zarebanadkouki, M.; Blagodatskaya, E.; Kuzyakov, Y. Rhizosphere shape of lentil and maize: Spatial distribution of enzyme activities. Soil Biol. Biochem. 2016, 96, 229–237. [Google Scholar] [CrossRef]
- Badri, D.V.; Zolla, G.; Bakker, M.; Manter, D.K.; Vivanco, J.M. Potential impact of soil microbiomes on the leaf metabolome and on herbivore feeding behavior. New Phytol. 2013, 198, 264–273. [Google Scholar] [CrossRef]
- Arora, N.K.; Tewari, S.; Singh, R. Multifaceted Plant-Associated Microbes and Their Mechanisms Diminish the Concept of Direct and Indirect PGPRs. In Plant-Microbe Symbiosis, Fundamentals and Advances; Arora, N.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 411–449. [Google Scholar]
- Wang, Q.; Jiang, X.; Guan, D.; Wei, D.; Zhao, B.; Ma, M.; Chen, S.; Li, L.; Cao, F.; Li, J. Long-term fertilization changes bacterial diversity and bacterial communities in the maize rhizosphere of Chinese Mollisols. Appl. Soil Ecol. 2018, 125, 88–96. [Google Scholar] [CrossRef]
- Mendes, L.W.; Kuramae, E.E.; Navarrete, A.A.; Van Veen, J.A.; Tsai, S.M. Taxonomical and functional microbial community selection in soybean rhizosphere. ISME J. 2014, 8, 1577–1587. [Google Scholar] [CrossRef]
- Bakker, M.G.; Chaparro, J.M.; Manter, D.; Vivanco, J.M. Impacts of bulk soil microbial community structure on rhizosphere microbiomes of Zea mays. Plant Soil 2015, 392, 115–126. [Google Scholar] [CrossRef]
- French, K.; Tkacz, A.; Turnbull, L.A. Conversion of grassland to arable decreases microbial diversity and alters community composition. Appl. Soil Ecol. 2017, 110, 43–52. [Google Scholar] [CrossRef]
- Lan, G.; Li, Y.; Jatoi, M.T.; Tan, Z.; Wu, Z.; Xie, G. Change in Soil Microbial Community Compositions and Diversity Following the Conversion of Tropical Forest to Rubber Plantations in Xishuangbanan, Southwest China. Trop. Conserv. Sci. 2017, 10, 1940082917733230. [Google Scholar] [CrossRef]
- Wolińska, A.; Górniak, D.; Zielenkiewicz, U.; Goryluk-Salmonowicz, A.; Kuźniar, A.; Stępniewska, Z.; Błaszczyk, M. Microbial biodiversity in arable soils is affected by agricultural practices. Int. Agrophys. 2017, 31, 259–271. [Google Scholar] [CrossRef]
- Hallama, M.; Pekrun, C.; Lambers, H.; Kandeler, E. Hidden miners—the roles of cover crops and soil microorganisms in phosphorus cycling through agroecosystems. Plant Soil 2019, 434, 7–45. [Google Scholar] [CrossRef]
- Dahal, B.; NandaKafle, G.; Perkins, L.; Brözel, V.S. Diversity of free-Living nitrogen fixing Streptomyces in soils of the badlands of South Dakota. Microbiol. Res. 2017, 195, 31–39. [Google Scholar] [CrossRef]
- Kumar, S.; Suyal, D.C.; Yadav, A.; Shouche, Y.; Goel, R. Microbial diversity and soil physiochemical characteristic of higher altitude. PLoS ONE 2019, 14, e0213844. [Google Scholar] [CrossRef]
- Xu, Y.; Du, A.; Wang, Z.; Zhu, W.; Li, C.; Wu, L. Effects of different rotation periods of Eucalyptus plantations on soil physiochemical properties, enzyme activities, microbial biomass and microbial community structure and diversity. For. Ecol. Manag. 2019, 456, 117683. [Google Scholar] [CrossRef]
- Zhao, Z.; Liu, G.; Liu, Q.; Huang, C.; Li, H.; Wu, C. Distribution Characteristics and Seasonal Variation of Soil Nutrients in the Mun River Basin, Thailand. Int. J. Environ. Res. Public Health 2018, 15, 1818. [Google Scholar] [CrossRef]
- Bao, S.D. Soil and Agricultural Chemistry Analysis; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Qiu, J.; Cao, J.; Lan, G.; Liang, Y.; Wang, H.; Li, Q. The Influence of Land Use Patterns on Soil Bacterial Community Structure in the Karst Graben Basin of Yunnan Province, China. Forests 2020, 11, 51. [Google Scholar] [CrossRef]
- Pang, Y.P.; Liu, K.; Zhao, Z.L.; Li, X.T. Study on Identification Method of Fructus Lycii Origin by Mineral Elements. Adv. Mater. Res. 2014, 989–994, 1368–1371. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Bittinger, K.; Bushman, F.D.; DeSantis, T.Z.; Andersen, G.L.; Knight, R. PyNAST: A flexible tool for aligning sequences to a template alignment. Bioinformatics 2010, 26, 266–267. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Ssekagiri, A.T.; Sloan, W.; Ijaz, U.Z. microbiomeSeq: An R package for analysis of microbial communities in an environmental context. In Proceedings of the ISCB Africa ASBCB Conference, Kumasi, Ghana, 10–12 October 2017; Available online: Https://Github.Com/Umerijaz/MicrobiomeSeq (accessed on 14 February 2022).
- Causey, B.D. Parametric estimation of the number of classes in a population. J. Appl. Stat. 2002, 29, 925–934. [Google Scholar] [CrossRef]
- Shannon, C.E. The mathematical theory of communication. MD Comput. 1997, 14, 306–317. [Google Scholar] [PubMed]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Community Ecology Package, R Packag version (2.6-2); 2022. Available online: https://github.com/vegandevs/vegan (accessed on 27 April 2022).
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Belsley, D.A.; Kuh, E.; Welsch, R.E. Regression Diagnostics: Identifying Influential Data and Sources of Collinearity; John Wiley & Sons: Hoboken, NJ, USA, 2005; Volume 571. [Google Scholar]
- Legendre, P.; Oksanen, J.; ter Braak, C.J.F. Testing the significance of canonical axes in redundancy analysis. Methods Ecol. Evol. 2011, 2, 269–277. [Google Scholar] [CrossRef]
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W.; Liaw, A.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S.; et al. gplots: Various R Programming Tools for Plotting Data, R Packag version (3.1.3); 2022; Available online: https://CRAN.R-project.org/package=gplots (accessed on 15 May 2022).
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.A.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Lauber, C.L.; Strickland, M.S.; Bradford, M.A.; Fierer, N. The influence of soil properties on the structure of bacterial and fungal communities across land-use types. Soil Biol. Biochem. 2008, 40, 2407–2415. [Google Scholar] [CrossRef]
- Tayyab, M.; Islam, W.; Lee, C.G.; Pang, Z.; Khalil, F.; Lin, S.; Lin, W.; Zhang, H. Short-Term Effects of Different Organic Amendments on Soil Fungal Composition. Sustainability 2019, 11, 198. [Google Scholar] [CrossRef]
- Gill, A.S.; Purnell, K.; Palmer, M.I.; Stein, J.; McGuire, K.L. Microbial Composition and Functional Diversity Differ Across Urban Green Infrastructure Types. Front. Microbiol. 2020, 11, 912. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tremblay, J.; Bainard, L.D.; Cade-Menun, B.; Hamel, C. Long-term effects of nitrogen and phosphorus fertilization on soil microbial community structure and function under continuous wheat production. Environ. Microbiol. 2020, 22, 1066–1088. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, P.; de Figueroa, L.I.; Pajot, H.F. Dual Purpose of ligninolytic- basidiomycetes: Mycoremediation of bioethanol distillation vinasse coupled to sustainable bio-based compounds production. Fungal Biol. Rev. 2020, 34, 25–40. [Google Scholar] [CrossRef]
- Wilhelm, R.C.; Cardenas, E.; Leung, H.; Szeitz, A.; Jensen, L.D.; Mohn, W.W. Long-term enrichment of stress-tolerant cellulolytic soil populations following timber harvesting evidenced by multi-omic stable isotope probing. Front Microbiol. 2017, 8, 537. [Google Scholar]
- Hol, W.H.G.; de Boer, W.; Termorshuizen, A.J.; Meyer, K.M.; Schneider, J.H.M.; van Dam, N.M.; Van Veen, J.A.; van der Putten, W.H. Reduction of rare soil microbes modifies plant-herbivore interactions. Ecol. Lett. 2010, 13, 292–301. [Google Scholar] [CrossRef]
- Li, X.; Rui, J.; Mao, Y.; Yannarell, A.; Mackie, R. Dynamics of the bacterial community structure in the rhizosphere of a maize cultivar. Soil Biol. Biochem. 2014, 68, 392–401. [Google Scholar] [CrossRef]
- Fierer, N.; Lauber, C.L.; Ramirez, K.S.; Zaneveld, J.; Bradford, M.A.; Knight, R. Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME J. 2012, 6, 1007–1017. [Google Scholar] [CrossRef]
- Chakdar, H.; Dastager, S.G.; Khire, J.M.; Rane, D.; Dharne, M.S. Characterization of mineral phosphate solubilizing and plant growth promoting bacteria from termite soil of arid region. 3 Biotech 2018, 8, 463. [Google Scholar] [CrossRef]
- Liang, J.-L.; Liu, J.; Jia, P.; Yang, T.-T.; Zeng, Q.-W.; Zhang, S.-C.; Liao, B.; Shu, W.-S.; Li, J.-T. Novel phosphate-solubilizing bacteria enhance soil phosphorus cycling following ecological restoration of land degraded by mining. ISME J. 2020, 14, 1600–1613. [Google Scholar] [CrossRef]
- Wu, Y.; Shen, Y.; Zhu, Y.; Mupunga, J.; Zou, L.; Liu, C.; Liu, S.; Mao, J. Broccoli ingestion increases the glucosinolate hydrolysis activity of microbiota in the mouse gut. Int. J. Food Sci. Nutr. 2019, 70, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Taye, Z.M.; Helgason, B.L.; Bell, J.K.; Norris, C.E.; Vail, S.; Robinson, S.J.; Parkin, I.A.P.; Arcand, M.; Mamet, S.; Links, M.G.; et al. Core and Differentially Abundant Bacterial Taxa in the Rhizosphere of Field Grown Brassica napus Genotypes: Implications for Canola Breeding. Front. Microbiol. 2020, 10, 3007. [Google Scholar] [CrossRef] [PubMed]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; van der Putten, W.H. Going back to the roots: The microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Hardoim, P.R.; Sessitsch, A.; Reinhold-Hurek, B.; van Overbeek, L.S.; Van, E. Assessment of rice root endophytes and their potential for plant growth promotion. In Bact Endophytes Rice—Their Divers Charact Perspect Groningen; Hardoim, P.R., Ed.; 2011; pp. 77–100. Available online: https://pure.rug.nl/ws/portalfiles/portal/2529913/ThesisHardoim.pdf (accessed on 5 June 2022).
- Li, Y.H.; Zhu, J.N.; Zhai, Z.H.; Zhang, Q. Endophytic bacterial diversity in roots of Phragmites australis in constructed Beijing Cuihu Wetland (China). FEMS Microbiol. Lett. 2010, 309, 84–93. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, N.; Hu, Y.; Sun, G. Changes in soil physicochemical properties and soil bacterial community in mulberry (Morus alba L.)/alfalfa (Medicago sativa L.) intercropping system. Microbiologyopen 2018, 7, e00555. [Google Scholar] [CrossRef]
- Fawaz, M.N. Revealing the Ecological Role of Gemmatimonadetes through Cultivation and Molecular Analysis of Agricultural Soils. Master’s Thesis, University of Tennessee, Knoxville, TN, USA, 2013. [Google Scholar]
- Yuan, X.; Knelman, J.E.; Wang, D.; Goebl, A.; Gasarch, E.; Seastedt, T.R. Patterns of Soil Bacterial Richness and Composition Tied to Plant Richness, Soil Nitrogen, and Soil Acidity in Alpine Tundra. Arct. Antarct. Alp. Res. 2017, 49, 441–453. [Google Scholar] [CrossRef]
- Boparai, J.K.; Sharma, P.K. Metagenomics and Metatranscriptomics Approaches in Understanding and Discovering Novel Molecules in Rhizosphere Environment. In Omics Science for Rhizosphere Biology; Springer: Berlin/Heidelberg, Germany, 2021; pp. 41–52. [Google Scholar]
- Paul, A.; Dziallas, C.; Zwirnmann, E.; Gjessing, E.T.; Grossart, H.-P. UV irradiation of natural organic matter (NOM): Impact on organic carbon and bacteria. Aquat. Sci. 2012, 74, 443–454. [Google Scholar] [CrossRef]
- Whalley, W.R.; Riseley, B.; Leeds-Harrison, P.B.; Bird, N.R.A.; Leech, P.K.; Adderley, W.P. Structural differences between bulk and rhizosphere soil. Eur. J. Soil Sci. 2005, 56, 353–360. [Google Scholar] [CrossRef]
- Simões, M.F.; Antunes, A.; Ottoni, C.; Amini, M.S.; Alam, I.; Alzubaidy, H.; Mokhtar, N.-A.; Archer, J.A.; Bajic, V.B. Soil and Rhizosphere Associated Fungi in Gray Mangroves (Avicennia marina) from the Red Sea—A Metagenomic Approach. Genom. Proteom. Bioinform. 2015, 13, 310–320. [Google Scholar] [CrossRef]
- Sugiyama, A. The soybean rhizosphere: Metabolites, microbes, and beyond—A review. J. Adv. Res. 2019, 19, 67–73. [Google Scholar] [CrossRef]
- Wang, C.; Jiang, K.; Zhou, J.; Wu, B. Solidago canadensis invasion affects soil N-fixing bacterial communities in heterogeneous landscapes in urban ecosystems in East China. Sci. Total Environ. 2018, 631–632, 702–713. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Fu, B.J.; Gao, G.Y.; Yao, X.L.; Zhou, J. Soil moisture and evapotranspiration of different land cover types in the Loess Plateau, China. Hydrol. Earth Syst. Sci. 2012, 16, 2883–2892. [Google Scholar] [CrossRef]
- Yadav, A.N.; Kumar, R.; Kumar, S.; Kumar, V.; Sugitha, T.; Singh, B.; Chauahan, V.S.; Dhaliwal, H.S.; Saxena, A.K. Beneficial microbiomes: Biodiversity and potential biotechnological applications for sustainable agriculture and human health. J. Appl. Biol. Biotechnol. 2017, 5, 45–57. [Google Scholar] [CrossRef]
- Ghosh, S.K.; Bera, T.; Chakrabarty, A.M. Microbial siderophore–A boon to agricultural sciences. Biol Control. 2020, 144, 104214. [Google Scholar] [CrossRef]
- Dalal, R.C.; Allen, D.E.; Livesley, S.J.; Richards, G. Magnitude and biophysical regulators of methane emission and consumption in the Australian agricultural, forest, and submerged landscapes: A review. Plant Soil 2008, 309, 43–76. [Google Scholar] [CrossRef]
- Kaiser, C.; Koranda, M.; Kitzler, B.; Fuchslueger, L.; Schnecker, J.; Schweiger, P.; Rasche, F.; Zechmeister-Boltenstern, S.; Sessitsch, A.; Richter, A. Belowground carbon allocation by trees drives seasonal patterns of extracellular enzyme activities by altering microbial community composition in a beech forest soil. New Phytol. 2010, 187, 843–858. [Google Scholar] [CrossRef]
- Ma, B.; Cai, Y.; Bork, E.W.; Chang, S.X. Defoliation intensity and elevated precipitation effects on microbiome and interactome depend on site type in northern mixed-grass prairie. Soil Biol. Biochem. 2018, 122, 163–172. [Google Scholar] [CrossRef]
- Orkin, J.D.; Campos, F.A.; Myers, M.S.; Hernandez, S.E.C.; Guadamuz, A.; Melin, A.D. Seasonality of the gut microbiota of free-ranging white-faced capuchins in a tropical dry forest. ISME J. 2019, 13, 183–196. [Google Scholar] [CrossRef]
- Hug, L.A.; Castelle, C.J.; Wrighton, K.C.; Thomas, B.C.; Sharon, I.; Frischkorn, K.R.; Williams, K.H.; Tringe, S.G.; Banfield, J.F. Community genomic analyses constrain the distribution of metabolic traits across the Chloroflexi phylum and indicate roles in sediment carbon cycling. Microbiome 2013, 1, 22. [Google Scholar] [CrossRef]
- Muturi, S.M.; Muthui, L.W.; Njogu, P.M.; Onguso, J.M.; Wachira, F.N.; Opiyo, S.O.; Pelle, R. Metagenomics survey unravels diversity of biogas microbiomes with potential to enhance productivity in Kenya. PLoS ONE 2021, 16, e0244755. [Google Scholar] [CrossRef]
- Zhao, S.; Qiu, S.; Xu, X.; Ciampitti, I.A.; Zhang, S.; He, P. Change in straw decomposition rate and soil microbial community composition after straw addition in different long-term fertilization soils. Appl. Soil Ecol. 2019, 138, 123–133. [Google Scholar] [CrossRef]
- Okal, E.J.; Aslam, M.M.; Karanja, J.K.; Nyimbo, W.J. Mini review: Advances in understanding regulation of cellulase enzyme in white-rot basidiomycetes. Microb. Pathog. 2020, 147, 104410. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, N.; Zhang, J.; Hu, Y.; Cai, D.; Guo, J.; Wu, D.; Sun, G. Soil Physicochemical Properties and the Rhizosphere Soil Fungal Community in a Mulberry (Morus alba L.)/Alfalfa (Medicago sativa L.) Intercropping System. Forests 2019, 10, 167. [Google Scholar] [CrossRef]
- Ranjan Rout, G.; Das, P. Effect of metal toxicity on plant growth and metabolism: I. Zinc. In Sustainable Agriculture; Springer: Dordrecht, The Netherlands, 2009; pp. 873–884. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Bacteria | Fungi | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genus | Family | Order | Class | Genus | Family | Order | Class | |||||||||
| F | p-Value | F | p-Value | F | p-Value | F | p-Value | F | p-Value | F | p-Value | F | p-Value | F | p-Value | |
| RDA Models | 34.634 | 0.001 | 34.883 | 0.001 | 27.976 | 0.001 | 23.181 | 0.001 | 38.746 | 0.001 | 38.474 | 0.001 | 37.994 | 0.001 | 38.153 | 0.001 |
| Physical–chemical properties | ||||||||||||||||
| Phosphorus (P) | 19.214 | 0.001 | 19.7822 | 0.001 | 14.9689 | 0.001 | 18.5632 | 0.001 | 28.3599 | 0.001 | 20.9856 | 0.001 | 20.5855 | 0.001 | 16.2982 | 0.001 |
| Zinc (Zn) | 50.1576 | 0.001 | 56.7291 | 0.001 | 42.674 | 0.001 | 25.7486 | 0.001 | 76.3496 | 0.001 | 68.1285 | 0.001 | 69.0908 | 0.001 | 80.1055 | 0.001 |
| Manganese (Mn) | 128.0654 | 0.001 | 133.6238 | 0.001 | 97.6005 | 0.001 | 62.3831 | 0.001 | 86.8815 | 0.001 | 84.988 | 0.001 | 84.6954 | 0.001 | 98.778 | 0.001 |
| Iron (Fe) | 4.87 | 0.003 | 4.5214 | 0.006 | 4.6515 | 0.002 | 3.5794 | 0.007 | 17.8615 | 0.001 | 13.8971 | 0.001 | 14.9338 | 0.001 | 11.0018 | 0.001 |
| Potassium (K) | 32.2536 | 0.001 | 27.3877 | 0.001 | 26.5556 | 0.001 | 24.7061 | 0.001 | 60.6288 | 0.001 | 76.7898 | 0.001 | 80.2399 | 0.001 | 72.6909 | 0.001 |
| Nitrogen (N) | 4.2549 | 0.007 | 6.6598 | 0.001 | 6.5038 | 0.003 | 4.992 | 0.003 | 10.7669 | 0.001 | 11.7058 | 0.001 | 12.6321 | 0.001 | 17.9998 | 0.001 |
| Calcium (Ca) | 39.5991 | 0.001 | 34.0089 | 0.001 | 30.0193 | 0.001 | 27.4398 | 0.001 | 28.406 | 0.001 | 28.6641 | 0.001 | 30.827 | 0.001 | 26.0269 | 0.001 |
| Land use variables | ||||||||||||||||
| Brassica | 19.1152 | 0.001 | 16.7318 | 0.001 | 13.1291 | 0.001 | 5.5959 | 0.001 | 41.9085 | 0.001 | 42.9349 | 0.001 | 40.7998 | 0.001 | 44.5503 | 0.001 |
| Non-brassica | 17.4948 | 0.001 | 12.0336 | 0.001 | 9.4166 | 0.001 | 10.5305 | 0.001 | 20.4728 | 0.001 | 15.9562 | 0.001 | 19.4612 | 0.001 | 15.9813 | 0.001 |
| Cultivated | 68.6182 | 0.001 | 71.9228 | 0.001 | 55.3713 | 0.001 | 43.1516 | 0.001 | 39.5823 | 0.001 | 53.9929 | 0.001 | 51.8795 | 0.001 | 41.8935 | 0.001 |
| Fallow | 28.9311 | 0.001 | 26.7255 | 0.001 | 19.0214 | 0.001 | 21.5353 | 0.001 | 35.8917 | 0.001 | 18.0619 | 0.001 | 18.1037 | 0.001 | 13.3909 | 0.001 |
| Forest | 52.392 | 0.001 | 36.0979 | 0.001 | 30.7278 | 0.001 | 18.8807 | 0.001 | 21.662 | 0.001 | 21.7392 | 0.001 | 22.784 | 0.001 | 25.6496 | 0.001 |
| Grassland | 4.7679 | 0.003 | 5.8625 | 0.002 | 5.6152 | 0.001 | 3.5213 | 0.006 | 4.6772 | 0.001 | 8.8551 | 0.001 | 9.3071 | 0.001 | 9.7474 | 0.001 |
| Local variables | ||||||||||||||||
| Host plant | 11.7741 | 0.001 | 29.9131 | 0.001 | 21.128 | 0.001 | 29.5702 | 0.001 | 82.7351 | 0.001 | 79.338 | 0.001 | 65.4933 | 0.001 | 69.0251 | 0.001 |
| Soil type | 37.9961 | 0.001 | 41.2481 | 0.001 | 42.2607 | 0.001 | 47.5125 | 0.001 | 25.0033 | 0.001 | 31.0807 | 0.001 | 29.0829 | 0.001 | 29.1517 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, U.; Saqib, H.S.A.; Islam, W.; Prashant, P.; Patel, N.; Chen, W.; Yang, F.; You, M.; He, W. Landscape Composition and Soil Physical–Chemical Properties Drive the Assemblages of Bacteria and Fungi in Conventional Vegetable Fields. Microorganisms 2022, 10, 1202. https://doi.org/10.3390/microorganisms10061202
Kumar U, Saqib HSA, Islam W, Prashant P, Patel N, Chen W, Yang F, You M, He W. Landscape Composition and Soil Physical–Chemical Properties Drive the Assemblages of Bacteria and Fungi in Conventional Vegetable Fields. Microorganisms. 2022; 10(6):1202. https://doi.org/10.3390/microorganisms10061202
Chicago/Turabian StyleKumar, Uttam, Hafiz Sohaib Ahmed Saqib, Waqar Islam, Parmar Prashant, Nidhibahen Patel, Wei Chen, Feiying Yang, Minsheng You, and Weiyi He. 2022. "Landscape Composition and Soil Physical–Chemical Properties Drive the Assemblages of Bacteria and Fungi in Conventional Vegetable Fields" Microorganisms 10, no. 6: 1202. https://doi.org/10.3390/microorganisms10061202
APA StyleKumar, U., Saqib, H. S. A., Islam, W., Prashant, P., Patel, N., Chen, W., Yang, F., You, M., & He, W. (2022). Landscape Composition and Soil Physical–Chemical Properties Drive the Assemblages of Bacteria and Fungi in Conventional Vegetable Fields. Microorganisms, 10(6), 1202. https://doi.org/10.3390/microorganisms10061202