
Synthesis of Aroma Compounds as a Function of Different Nitrogen Sources in Fermentations Using Non-Saccharomyces Wine Yeasts
 , , ,
, , ,
Abstract
:1. Introduction

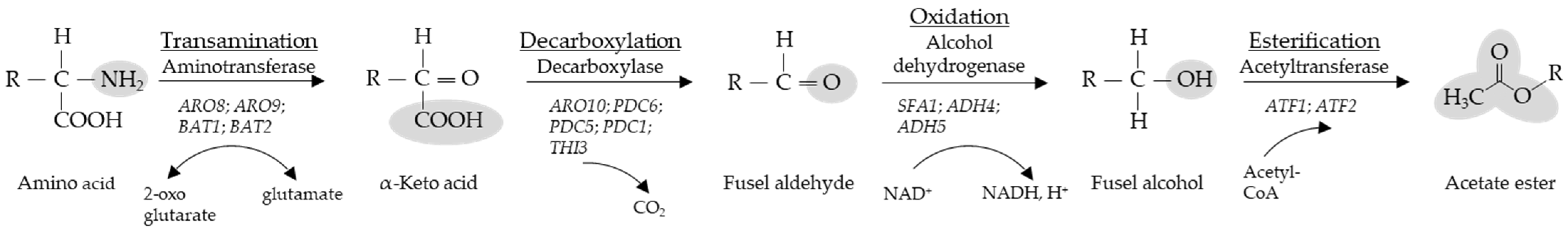{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | α-Keto Acid | Fusel Aldehyde | Fusel Alcohol | Acetate Ester | Odor Descriptor of Acetate Ester |
|---|---|---|---|---|---|
| Leu | α-Keto-isocaproate | 3-Methyl- butanal | Isoamyl alcohol | Isoamyl acetate | Banana, sweet, fruity |
| Val | α-Keto-isovalerate | 2-Methyl- propanal | Isobutanol | Isobutyl acetate | Sweet, fruity, tropical |
| Ile | α-Keto-3-methylvalerate | 2-Methyl- butanal | 2-Methylbutanol | 2-Methylbutyl acetate | Fruity, tropical, overripe fruit |
| Phe | Phenylpyruvate | 2-Phenyl- ethanal | 2-Phenyl ethanol | 2-Phenylethyl acetate | Floral, rose, honey |
| Tyr | 4-Hydroxyphenylpyruvate | 2-(4-Hydroxyphenyl) ethanal | Tyrosol | Tyrosol acetate | Fruity, flowery |
| Trp | Indol-3-pyruvate | 2-(Indol-3-yl) ethanal | Tryptophol | Tryptophol acetate | Fruity, flowery |
| Met | 2-Keto-4-methylthio-2-oxobutyrate | 3-(Methylthio) propanal | Methionol | Methionyl acetate | Cabbage, potato, mushroom |
| Thr | α-Keto-butyrate | Butanal | Butanol | Butyl acetate | Solvent, fruity, banana |
| Cys | 3-Mercapto- pyruvate | Mercaptoacetaldehyde | 2-Mercaptoethanol | 2-Mercaptoethyl acetate | Roasted |
2. Materials and Methods
2.1. Yeast Strains and Media
2.2. Fermentation Experiments
2.3. Analysis of Amino Acid Uptake with Ion Exchange Chromatography (IEC)
2.4. High-Performance Liquid Chromatography (HPLC) Analysis
2.5. Analysis of VOCs with Head-Space Solid Phase Microextraction Gas Chromatography (HS-SPME-GC-MS)
2.6. Statistical Evaluation and Software
3. Results
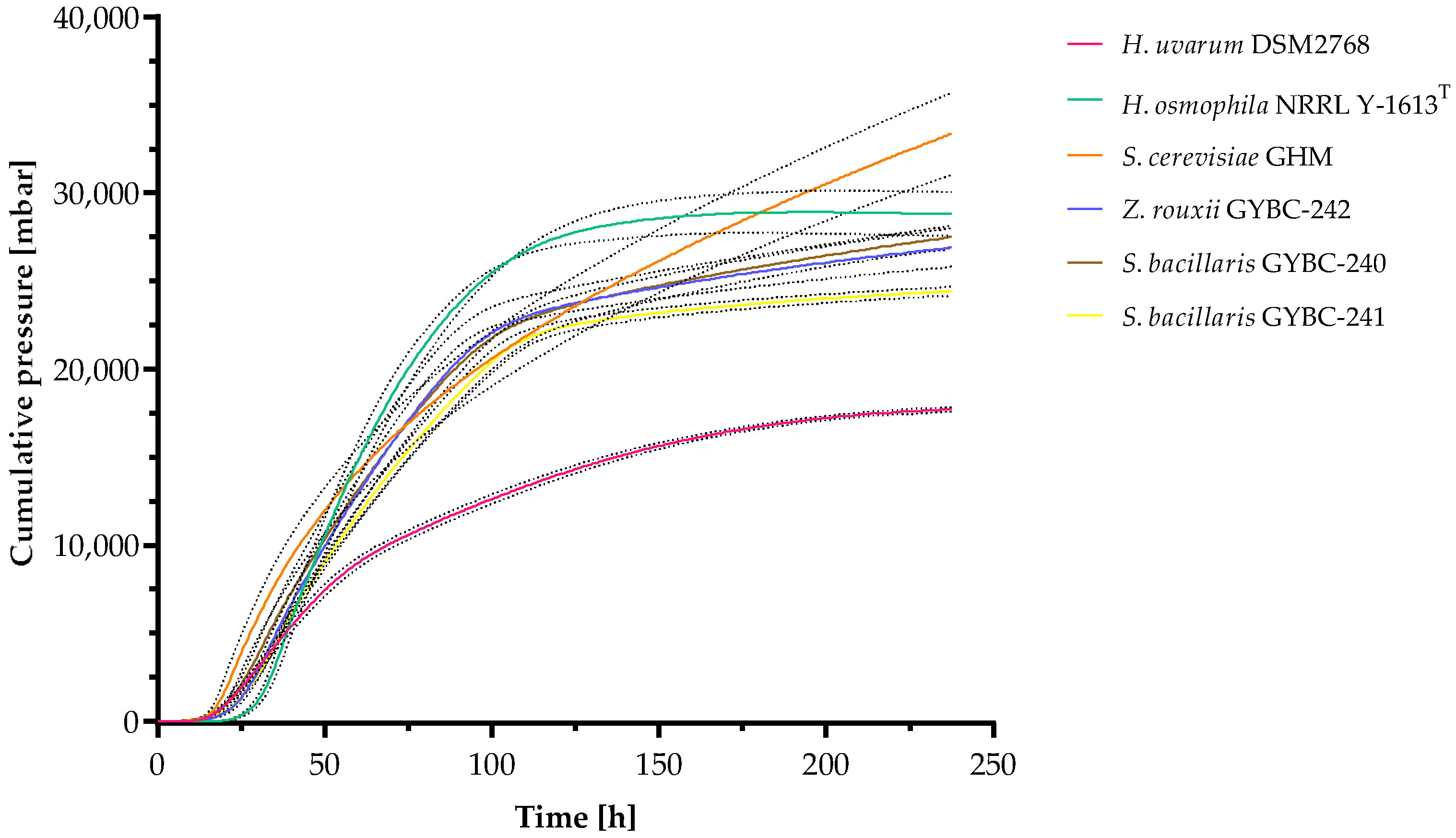
3.1. Fermentation Kinetics
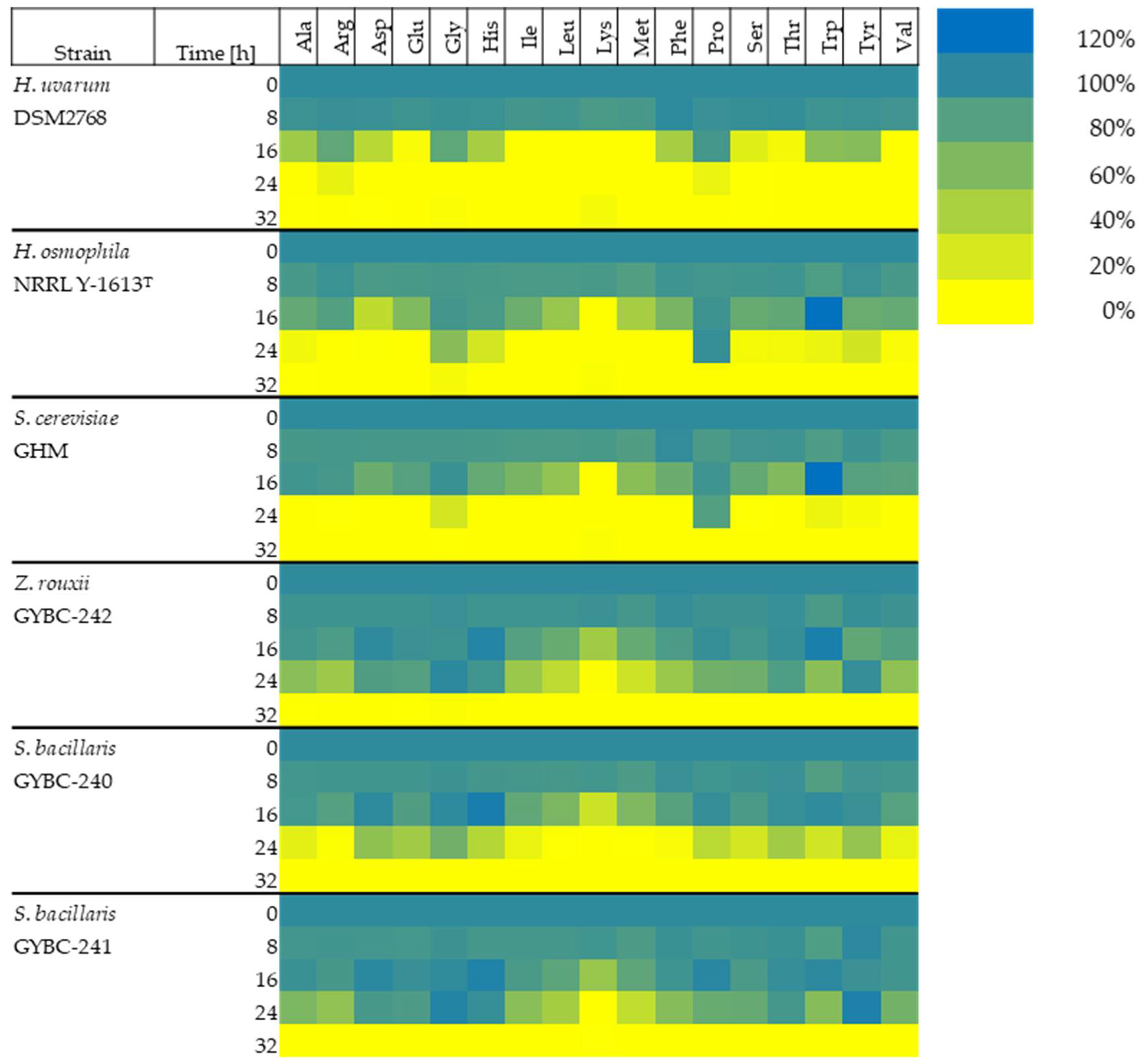
3.2. Evaluation of Amino Acid Consumption Rate
3.3. Analysis of Sugar Content
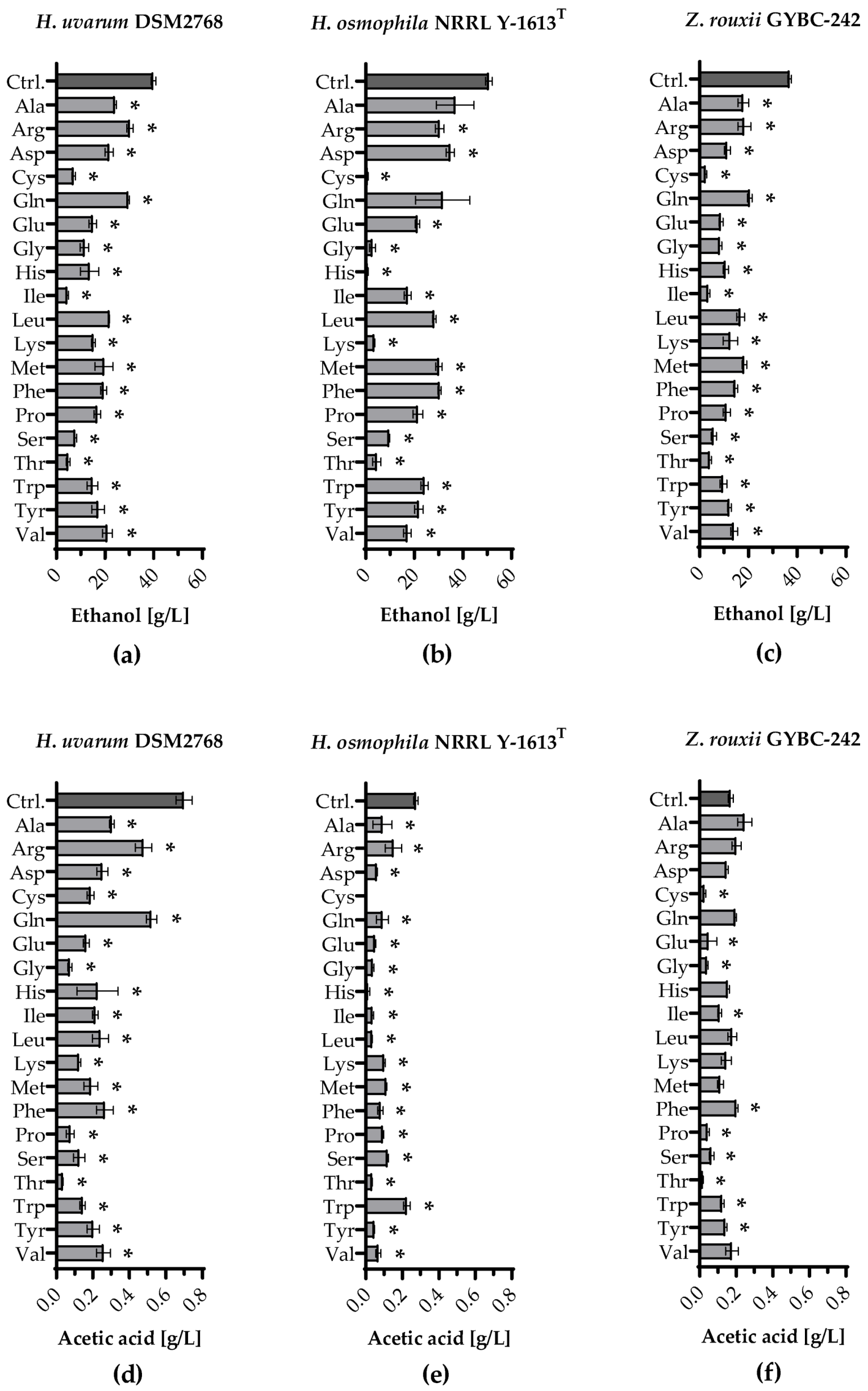
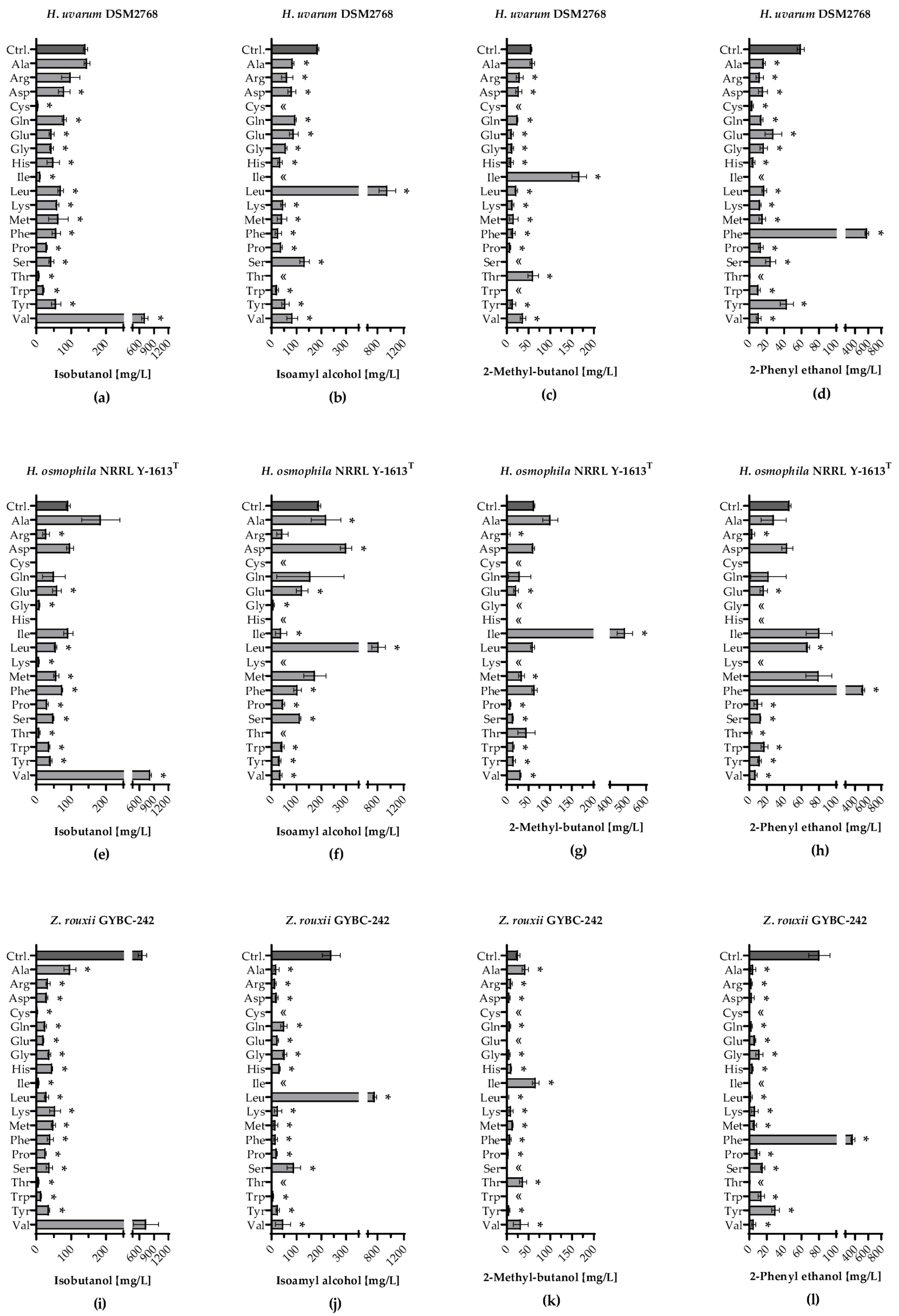
3.4. Volatile Organic Compounds Produced during Single Amino Acid Fermentations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bisson, L.F. Stuck and sluggish fermentations. Am. J. Enol. Vitic. 1999, 50, 107. [Google Scholar] [CrossRef]
- Bell, S.-J.; Henschke, P.A. Implications of nitrogen nutrition for grapes, fermentation and wine. Aust. J. Grape Wine Res. 2005, 11, 242–295. [Google Scholar] [CrossRef]
- Kessi-Pérez, E.I.; Molinet, J.; Martínez, C. Disentangling the genetic bases of Saccharomyces cerevisiae nitrogen consumption and adaptation to low nitrogen environments in wine fermentation. Biol. Res. 2020, 53, 2. [Google Scholar] [CrossRef] [PubMed]
- Molinet, J.; Cubillos, F.A.; Salinas, F.; Liti, G.; Martínez, C. Genetic variants of TORC1 signaling pathway affect nitrogen consumption in Saccharomyces cerevisiae during alcoholic fermentation. PLoS ONE 2019, 14, e0220515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapp, A.; Versini, G. Influence of nitrogen compounds in grapes on aroma compounds of wines. In Food Flavors: Generation, Analysis and Process Influence, Proceedings of the 8th International Flavor Conference, Cos, Greece, 6–8 July 1995; Elsevier: Amsterdam, The Netherlands, 1995; pp. 1659–1694. ISBN 9780444820136. [Google Scholar]
- Hernández-Orte, P.; Cacho, J.F.; Ferreira, V. Relationship between varietal amino acid profile of grapes and wine aromatic composition. Experiments with model solutions and chemometric study. J. Agric. Food Chem. 2002, 50, 2891–2899. [Google Scholar] [CrossRef]
- Dzialo, M.C.; Park, R.; Steensels, J.; Lievens, B.; Verstrepen, K.J. Physiology, ecology and industrial applications of aroma formation in yeast. FEMS Microbiol. Rev. 2017, 41, S95–S128. [Google Scholar] [CrossRef] [Green Version]
- Burin, V.M.; Gomes, T.M.; Caliari, V.; Rosier, J.P.; Bordignon Luiz, M.T. Establishment of influence the nitrogen content in musts and volatile profile of white wines associated to chemometric tools. Microchem. J. 2015, 122, 20–28. [Google Scholar] [CrossRef]
- Jiranek, V.; Langridge, P.; Henschke, P.A. Regulation of hydrogen sulfide liberation in wine-producing Saccharomyces cerevisiae strains by assimilable nitrogen. Appl. Environ. Microbiol. 1995, 61, 461–467. [Google Scholar] [CrossRef] [Green Version]
- Hazelwood, L.A.; Daran, J.-M.; van Maris, A.J.A.; Pronk, J.T.; Dickinson, J.R. The Ehrlich pathway for fusel alcohol production: A century of research on Saccharomyces cerevisiae metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef] [Green Version]
- Drumonde-Neves, J.; Fernandes, T.; Lima, T.; Pais, C.; Franco-Duarte, R. Learning from 80 years of studies: A comprehensive catalogue of non-Saccharomyces yeasts associated with viticulture and winemaking. FEMS Yeast Res. 2021, 21, foab017. [Google Scholar] [CrossRef]
- Van Wyk, N.; Grossmann, M.; Wendland, J.; von Wallbrunn, C.; Pretorius, I.S. The whiff of wine yeast innovation: Strategies for enhancing aroma production by yeast during wine fermentation. J. Agric. Food Chem. 2019, 67, 13496–13505. [Google Scholar] [CrossRef]
- Domizio, P.; Romani, C.; Lencioni, L.; Comitini, F.; Gobbi, M.; Mannazzu, I.; Ciani, M. Outlining a future for non-Saccharomyces yeasts: Selection of putative spoilage wine strains to be used in association with Saccharomyces cerevisiae for grape juice fermentation. Int. J. Food Microbiol. 2011, 147, 170–180. [Google Scholar] [CrossRef]
- Van Wyk, N.; von Wallbrunn, C.; Swiegers, J.H.; Pretorius, I.S. Biotechnology of Wine Yeasts. In Encyclopedia of Mycology; Elsevier: Amsterdam, The Netherlands, 2021; pp. 428–446. ISBN 9780323851800. [Google Scholar]
- Roca-Mesa, H.; Sendra, S.; Mas, A.; Beltran, G.; Torija, M.-J. Nitrogen preferences during alcoholic fermentation of different non-Saccharomyces yeasts of oenological interest. Microorganisms 2020, 8, 157. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.; Seguinot, P.; Sanchez, I.; Ortiz-Julien, A.; Heras, J.M.; Querol, A.; Camarasa, C.; Guillamón, J.M. Nitrogen sources preferences of non-Saccharomyces yeasts to sustain growth and fermentation under winemaking conditions. Food Microbiol. 2020, 85, 103287. [Google Scholar] [CrossRef]
- Lleixà, J.; Martín, V.; Giorello, F.; Portillo, M.C.; Carrau, F.; Beltran, G.; Mas, A. Analysis of the NCR mechanisms in Hanseniaspora vineae and Saccharomyces cerevisiae during winemaking. Front. Genet. 2018, 9, 747. [Google Scholar] [CrossRef] [Green Version]
- Gobert, A.; Tourdot-Maréchal, R.; Sparrow, C.; Morge, C.; Alexandre, H. Influence of nitrogen status in wine alcoholic fermentation. Food Microbiol. 2019, 83, 71–85. [Google Scholar] [CrossRef]
- Crépin, L.; Nidelet, T.; Sanchez, I.; Dequin, S.; Camarasa, C. Sequential use of nitrogen compounds by Saccharomyces cerevisiae during wine fermentation: A model based on kinetic and regulation characteristics of nitrogen permeases. Appl. Environ. Microbiol. 2012, 78, 8102–8111. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.; Pierce, J.S. Absorption of amino acids from wort by yeasts. J. Inst. Brew. 1964, 70, 307–315. [Google Scholar] [CrossRef]
- Valera, M.J.; Olivera, V.; Boido, E.; Dellacassa, E.; Carrau, F. Wine aroma characterization of the two main fermentation yeast species of the apiculate genus Hanseniaspora. Fermentation 2021, 7, 162. [Google Scholar] [CrossRef]
- Vermeulen, C.; Lejeune, I.; Tran, T.T.H.; Collin, S. Occurrence of polyfunctional thiols in fresh lager beers. J. Agric. Food Chem. 2006, 54, 5061–5068. [Google Scholar] [CrossRef]
- Bely, M.; Sablayrolles, J.-M.; Barre, P. Automatic detection of assimilable nitrogen deficiencies during alcoholic fermentation in oenological conditions. J. Ferm. Bioeng. 1990, 70, 246–252. [Google Scholar] [CrossRef]
- Su, Y.; Origone, A.C.; Rodríguez, M.E.; Querol, A.; Guillamón, J.M.; Lopes, C.A. Fermentative behaviour and competition capacity of cryotolerant Saccharomyces species in different nitrogen conditions. Int. J. Food Microbiol. 2019, 291, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seguinot, P.; Bloem, A.; Brial, P.; Meudec, E.; Ortiz-Julien, A.; Camarasa, C. Analysing the impact of the nature of the nitrogen source on the formation of volatile compounds to unravel the aroma metabolism of two non-Saccharomyces strains. Int. J. Food Microbiol. 2020, 316, 108441. [Google Scholar] [CrossRef] [PubMed]
- Van Wyk, N.; Michling, F.; Bergamo, D.; Brezina, S.; Pretorius, I.S.; von Wallbrunn, C.; Wendland, J. Effect of isomixing on grape must fermentations of ATF1-overexpressing wine yeast strains. Foods 2020, 9, 717. [Google Scholar] [CrossRef] [PubMed]
- Csapó, J.; Lóki, K.; Csapóné Kiss, Z.; Albert, C. Separation and determination of the amino acids by ion exchange column chromatography applying postcolumn derivatization. Acta Univ. Sapientiae Aliment. 2008, 1, 5–29. [Google Scholar]
- Krause, B.; Löhnertz, O. Saccharopin und Pipecolinsäure: Diagnostische Biomarker in der klassischen Aminosäureanalytik. MTA-Dialog 2017, 18, 316–321. [Google Scholar] [CrossRef]
- Badura, J.; van Wyk, N.; Brezina, S.; Pretorius, I.S.; Rauhut, D.; Wendland, J.; von Wallbrunn, C. Development of genetic modification tools for Hanseniaspora uvarum. Int. J. Mol. Sci. 2021, 22, 1943. [Google Scholar] [CrossRef]
- Scansani, S.; van Wyk, N.; Nader, K.B.; Beisert, B.; Brezina, S.; Fritsch, S.; Semmler, H.; Pasch, L.; Pretorius, I.S.; von Wallbrunn, C.; et al. The film-forming Pichia spp. in a winemaker’s toolbox: A simple isolation procedure and their performance in a mixed-culture fermentation of Vitis vinifera L. cv. Gewürztraminer must. Int. J. Food Microbiol. 2022, 365, 109549. [Google Scholar] [CrossRef]
- Tarasov, A.; Garzelli, F.; Schuessler, C.; Fritsch, S.; Loisel, C.; Pons, A.; Patz, C.-D.; Rauhut, D.; Jung, R. Wine storage at cellar vs. room conditions: Changes in the aroma composition of Riesling wine. Molecules 2021, 26, 6256. [Google Scholar] [CrossRef]
- Jung, R.; Kumar, K.; Patz, C.; Rauhut, D.; Tarasov, A.; Schüßler, C. Influence of transport temperature profiles on wine quality. Food Packag. Shelf Life 2021, 29, 100706. [Google Scholar] [CrossRef]
- Mas, A.; Guillamon, J.M.; Torija, M.J.; Beltran, G.; Cerezo, A.B.; Troncoso, A.M.; Garcia-Parrilla, M.C. Bioactive compounds derived from the yeast metabolism of aromatic amino acids during alcoholic fermentation. Biomed Res. Int. 2014, 2014, 898045. [Google Scholar] [CrossRef]
- Van Leeuwen, C.; Destrac-Irvine, A. Modified grape composition under climate change conditions requires adaptations in the vineyard. OENO One 2017, 51, 147. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Carrasco-Quiroz, M.; Martínez-Gil, A.M.; Pérez-Álvarez, E.P.; Garde-Cerdán, T.; Moreno-Simunovic, Y. Grape and wine amino acid composition from Carignan noir grapevines growing under rainfed conditions in the Maule Valley, Chile: Effects of location and rootstock. Food Res. Int. 2018, 105, 344–352. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Garde-Cerdán, T.; Rubio-Bretón, P.; Pérez-Álvarez, E.P. Study of must and wine amino acids composition after seaweed applications to Tempranillo blanco grapevines. Food Chem. 2020, 308, 125605. [Google Scholar] [CrossRef]
- Torrea, D.; Varela, C.; Ugliano, M.; Ancin-Azpilicueta, C.; Francis, I.L.; Henschke, P.A. Comparison of inorganic and organic nitrogen supplementation of grape juice—Effect on volatile composition and aroma profile of a Chardonnay wine fermented with Saccharomyces cerevisiae yeast. Food Chem. 2011, 127, 1072–1083. [Google Scholar] [CrossRef]
- Du Plessis, H.W.; Du Toit, M.; Hoff, J.W.; Hart, R.S.; Ndimba, B.K.; Jolly, N.P. Characterisation of non-Saccharomyces yeasts using different methodologies and evaluation of their compatibility with malolactic fermentation. S. Afr. J. Enol. Vitic. 2017, 38, 46–63. [Google Scholar] [CrossRef]
- Domizio, P.; Liu, Y.; Bisson, L.F.; Barile, D. Use of non-Saccharomyces wine yeasts as novel sources of mannoproteins in wine. Food Microbiol. 2014, 43, 5–15. [Google Scholar] [CrossRef]
- Steenwyk, J.L.; Opulente, D.A.; Kominek, J.; Shen, X.-X.; Zhou, X.; Labella, A.L.; Bradley, N.P.; Eichman, B.F.; Čadež, N.; Libkind, D.; et al. Extensive loss of cell-cycle and DNA repair genes in an ancient lineage of bipolar budding yeasts. PLOS Biol. 2019, 17, e3000255. [Google Scholar] [CrossRef] [Green Version]
- Barrajón-Simancas, N.; Giese, E.; Arévalo-Villena, M.; Ubeda, J.; Briones, A. Amino acid uptake by wild and commercial yeasts in single fermentations and co-fermentations. Food Chem. 2011, 127, 441–446. [Google Scholar] [CrossRef]
- Gobert, A.; Tourdot-Maréchal, R.; Morge, C.; Sparrow, C.; Liu, Y.; Quintanilla-Casas, B.; Vichi, S.; Alexandre, H. Non-Saccharomyces yeasts nitrogen source preferences: Impact on sequential fermentation and wine volatile compounds profile. Front. Microbiol. 2017, 8, 2175. [Google Scholar] [CrossRef] [Green Version]
- Žulj, M.M.; Tomaz, I.; Bandić, L.M.; Puhelek, I.; Korenika, A.J.; Jeromel, A. Influence of different yeast strains on metabolism of tryptophan and indole-3-acetic acid during fermentation. S. Afr. J. Enol. Vitic. 2016, 36, 44–49. [Google Scholar] [CrossRef]
- Malaney, G.W.; Tanner, R.D.; Rodrigues, A.M. The production of extracellular and intracellular free amino acids during aerated fermentation of glucose by baker’s yeast (Saccharomyces cerevisiae). Folia Microbiol. 1991, 36, 468–477. [Google Scholar] [CrossRef] [PubMed]
- Magyar, I.; Tóth, T. Comparative evaluation of some oenological properties in wine strains of Candida stellata, Candida zemplinina, Saccharomyces uvarum and Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Leandro, M.J.; Cabral, S.; Prista, C.; Loureiro-Dias, M.C.; Sychrová, H. The high-capacity specific fructose facilitator ZrFfz1 is essential for the fructophilic behavior of Zygosaccharomyces rouxii CBS 732T. Eukaryot. Cell 2014, 13, 1371–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rantsiou, K.; Englezos, V.; Torchio, F.; Risse, P.-A.; Cravero, F.; Gerbi, V.; Rolle, L.; Cocolin, L. Modeling of the fermentation behavior of Starmerella bacillaris. Am. J. Enol. Vitic. 2017, 68, 378–385. [Google Scholar] [CrossRef]
- Ehrlich, F. Über die Bedingungen der Fuselölbildung und über ihren Zusammenhang mit dem Eiweißaufbau der Hefe. Ber. Dtsch. Chem. Ges. 1907, 40, 1027–1047. [Google Scholar] [CrossRef] [Green Version]
- Neubauer, O.; Fromherz, K. Über den Abbau der Aminosäuren bei der Hefegärung. Hoppe-Seyler’s Z. Physiol. Chem. 1910, 70, 326–350. [Google Scholar] [CrossRef] [Green Version]
- Fairbairn, S.; McKinnon, A.; Musarurwa, H.T.; Ferreira, A.C.; Bauer, F.F. The impact of single amino acids on growth and volatile aroma production by Saccharomyces cerevisiae strains. Front. Microbiol. 2017, 8, 2554. [Google Scholar] [CrossRef] [Green Version]
- Espinosa Vidal, E.; de Morais, M.A.; François, J.M.; de Billerbeck, G.M. Biosynthesis of higher alcohol flavour compounds by the yeast Saccharomyces cerevisiae: Impact of oxygen availability and responses to glucose pulse in minimal growth medium with leucine as sole nitrogen source. Yeast 2015, 32, 47–56. [Google Scholar] [CrossRef]
- Martin, V.; Valera, M.; Medina, K.; Boido, E.; Carrau, F. Oenological impact of the Hanseniaspora/Kloeckera yeast genus on wines—A Review. Fermentation 2018, 4, 76. [Google Scholar] [CrossRef]
- Ingledew, W.M.; Magnus, C.A.; Sosulski, F.W. Influence of oxygen on proline utilization during the wine fermentation. Am. J. Enol. Vitic. 1987, 38, 246–248. [Google Scholar]






| Component | Concentration |
|---|---|
| Sugars | [g/L] |
| Glucose | 100 |
| Fructose | 100 |
| Acids | [g/L] |
| DL-Malic acid | 5 |
| Citric Acid | 0.5 |
| L-(+)-Tartaric acid | 3 |
| Minerals | [mg/L] |
| KH2PO4 | 750 |
| K2SO4 | 500 |
| MgSO4 | 250 |
| CaCl2 | 160 |
| NaCl | 200 |
| Trace elements | [mg/L] |
| MnSO4 | 4 |
| ZnSO4 | 4 |
| CuSO4 | 1 |
| KI | 1 |
| CoCl2 | 0.4 |
| H3BO3 | 1 |
| (NH4)6Mo7O24 | 1 |
| Vitamins | [mg/L] |
| Myo-inositol | 20 |
| Calcium pantothenate | 1.5 |
| Nicotinic acid | 2 |
| Thiamine hydrochloride | 0.25 |
| Pyridoxine hydrochloride | 0.25 |
| Biotin | 0.003 |
| Phytosterol solution for 100 L of SM | |
| β-Sitosterol | 500 mg |
| Tween 80 | 16.7 mL |
| Ethanol pure | 16.7 mL |
| Amino Acid | SM-mix [mg/L] * | SM-AA [mg/L] * |
|---|---|---|
| L-alanine | 48 | 891 |
| L-arginine | 123 | 581 |
| L-asparagine | 0 | 661 |
| L-aspartic acid | 15 | 1331 |
| L-cysteine | 4 | 1212 |
| L-glutamine | 40 | 1471 |
| L-glutamic acid | 166 | 731 |
| Glycine | 6 | 751 |
| L-histidine | 11 | 1552 |
| L-isoleucine | 11 | 1312 |
| L-leucine | 16 | 1312 |
| L-lysine | 6 | 731 |
| L-methionine | 10 | 1492 |
| L-phenylalanine | 13 | 1652 |
| L-proline | 202 | 1151 |
| L-serine | 26 | 1051 |
| L-threonine | 25 | 1191 |
| L-tryptophan | 59 | 2042 |
| L-tyrosine | 6 | 1812 |
| L-valine | 15 | 1172 |
| NH4Cl | 151 | 535 |
| Strain | Glucose [g/L] | Fructose [g/L] | Total Sugar [g/L] | Glucose * [%] | Fructose * [%] |
|---|---|---|---|---|---|
| H. uvarum DSM2768 | 52.46 ± 0.74 | 62.67 ± 1.70 | 115.13 ± 2.45 | 45.57 | 54.43 |
| H. osmophila NRRL Y-1613T | 33.29 ± 0.79 | 55.79 ± 0.71 | 89.08 ± 1.36 | 37.37 | 62.63 |
| S. cerevisiae GHM | 30.70 ± 0.80 | 68.83 ± 0.90 | 99.54 ± 1.70 | 30.84 | 69.15 |
| Z. rouxii GYBC-242 | 98.18 ± 1.31 | 20.09 ± 0.86 | 118.26 ± 2.17 | 83.02 | 16.99 |
| S. bacillaris GYBC-240 | 95.06 ± 0.84 | 19.84 ± 0.68 | 114.90 ± 0.17 | 82.73 | 17.27 |
| S. bacillaris GYBC-241 | 97.18 ± 0.94 | 22.46 ± 1.00 | 119.65 ± 0.49 | 81.22 | 18.77 |
| SM c | 101.24 ± 2.17 | 102.06 ± 2.20 | 203.30 ± 4.36 | 49.80 | 50.20 |
 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Total Alcohols a [g/L] | Total Acetate Esters b [mg/L] | Total Ethyl Esters c [mg/L] | |||||||
| H.u. | H.o. | Z.r. | H.u. | H.o. | Z.r. | H.u. | H.o. | Z.r. | |
| Ctrl. | 0.44 | 0.39 | 1.01 | 3.47 | 3.58 | 0.01 | 1.88 | 0.78 | 0.28 |
| Ala | 0.31 | 0.53 | 0.16 | 0.43 | 1.45 | 0.10 | 0.50 | 0.26 | 0.43 |
| Arg | 0.20 | 0.08 | 0.06 | 0.26 | 0.50 | 0.04 | 0.36 | 0.07 | 0.34 |
| Asp | 0.20 | 0.50 | 0.06 | 0.62 | 1.96 | 0.26 | 0.44 | 0.25 | 0.40 |
| Cys | 0.01 | 0.00 | 0.00 | 0.08 | 0.00 | 0.02 | 0.08 | 0.00 | 0.03 |
| Gln | 0.22 | 0.26 | 0.08 | 0.27 | 0.66 | 0.19 | 0.45 | 0.15 | 0.46 |
| Glu | 0.17 | 0.22 | 0.05 | 1.47 | 1.21 | 0.49 | 0.45 | 0.03 | 0.37 |
| Gly | 0.13 | 0.02 | 0.11 | 0.13 | 0.03 | 0.09 | 0.06 | 0.00 | 0.09 |
| His | 0.10 | 0.00 | 0.09 | 0.41 | 0.00 | 0.20 | 0.17 | 0.00 | 0.25 |
| Ile | 0.18 | 0.69 | 0.07 | 0.23 | 2.57 | 0.17 | 0.00 | 0.01 | 0.00 |
| Leu | 1.06 | 1.00 | 0.79 | 6.40 | 4.08 | 5.92 | 0.38 | 0.03 | 0.34 |
| Lys | 0.13 | 0.01 | 0.09 | 0.51 | 0.05 | 0.15 | 0.22 | 0.04 | 0.24 |
| Met | 0.14 | 0.35 | 0.08 | 0.06 | 2.30 | 0.03 | 0.14 | 0.05 | 0.28 |
| Phe | 0.68 | 0.76 | 0.43 | 3.15 | 7.99 | 2.68 | 0.18 | 0.08 | 0.24 |
| Pro | 0.09 | 0.09 | 0.06 | 0.41 | 0.44 | 0.15 | 0.26 | 0.00 | 0.34 |
| Ser | 0.20 | 0.19 | 0.14 | 0.13 | 0.35 | 0.68 | 0.00 | 0.00 | 0.06 |
| Thr | 0.07 | 0.05 | 0.04 | 0.40 | 0.12 | 0.11 | 0.52 | 0.49 | 0.77 |
| Trp | 0.05 | 0.11 | 0.03 | 0.06 | 0.04 | 0.10 | 0.19 | 0.07 | 0.28 |
| Tyr | 0.17 | 0.10 | 0.10 | 0.47 | 0.09 | 0.47 | 0.26 | 0.00 | 0.40 |
| Val | 0.84 | 0.90 | 0.82 | 0.41 | 0.36 | 0.34 | 5.25 | 3.30 | 13.68 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badura, J.; Medić, M.; Wyk, N.v.; Krause, B.; Semmler, H.; Brezina, S.; Pretorius, I.S.; Rauhut, D.; Wallbrunn, C.v. Synthesis of Aroma Compounds as a Function of Different Nitrogen Sources in Fermentations Using Non-Saccharomyces Wine Yeasts. Microorganisms 2023, 11, 14. https://doi.org/10.3390/microorganisms11010014
Badura J, Medić M, Wyk Nv, Krause B, Semmler H, Brezina S, Pretorius IS, Rauhut D, Wallbrunn Cv. Synthesis of Aroma Compounds as a Function of Different Nitrogen Sources in Fermentations Using Non-Saccharomyces Wine Yeasts. Microorganisms. 2023; 11(1):14. https://doi.org/10.3390/microorganisms11010014
Chicago/Turabian StyleBadura, Jennifer, Marko Medić, Niël van Wyk, Birgit Krause, Heike Semmler, Silvia Brezina, Isak S. Pretorius, Doris Rauhut, and Christian von Wallbrunn. 2023. "Synthesis of Aroma Compounds as a Function of Different Nitrogen Sources in Fermentations Using Non-Saccharomyces Wine Yeasts" Microorganisms 11, no. 1: 14. https://doi.org/10.3390/microorganisms11010014
APA StyleBadura, J., Medić, M., Wyk, N. v., Krause, B., Semmler, H., Brezina, S., Pretorius, I. S., Rauhut, D., & Wallbrunn, C. v. (2023). Synthesis of Aroma Compounds as a Function of Different Nitrogen Sources in Fermentations Using Non-Saccharomyces Wine Yeasts. Microorganisms, 11(1), 14. https://doi.org/10.3390/microorganisms11010014