
Biodegradation of Iprodione and Chlorpyrifos Using an Immobilized Bacterial Consortium in a Packed-Bed Bioreactor



Abstract
:1. Introduction
2. Materials and Methods
2.1. Pesticides
2.2. Bacterial Strains and Culture Media
2.3. Pesticide Degradation by Individual Bacterial Strains and Free Bacterial Consortium
2.4. Immobilization of Bacterial Strains
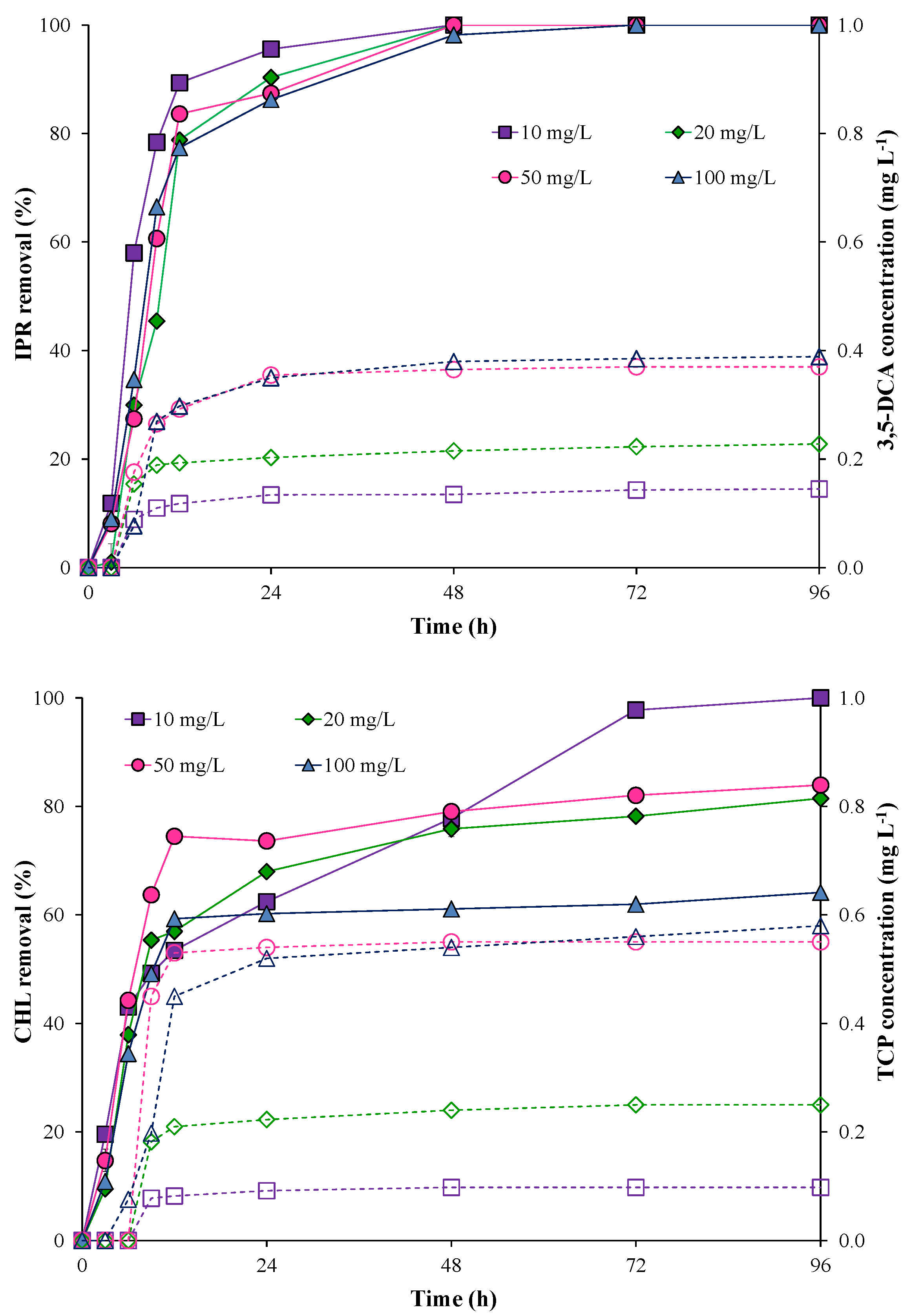
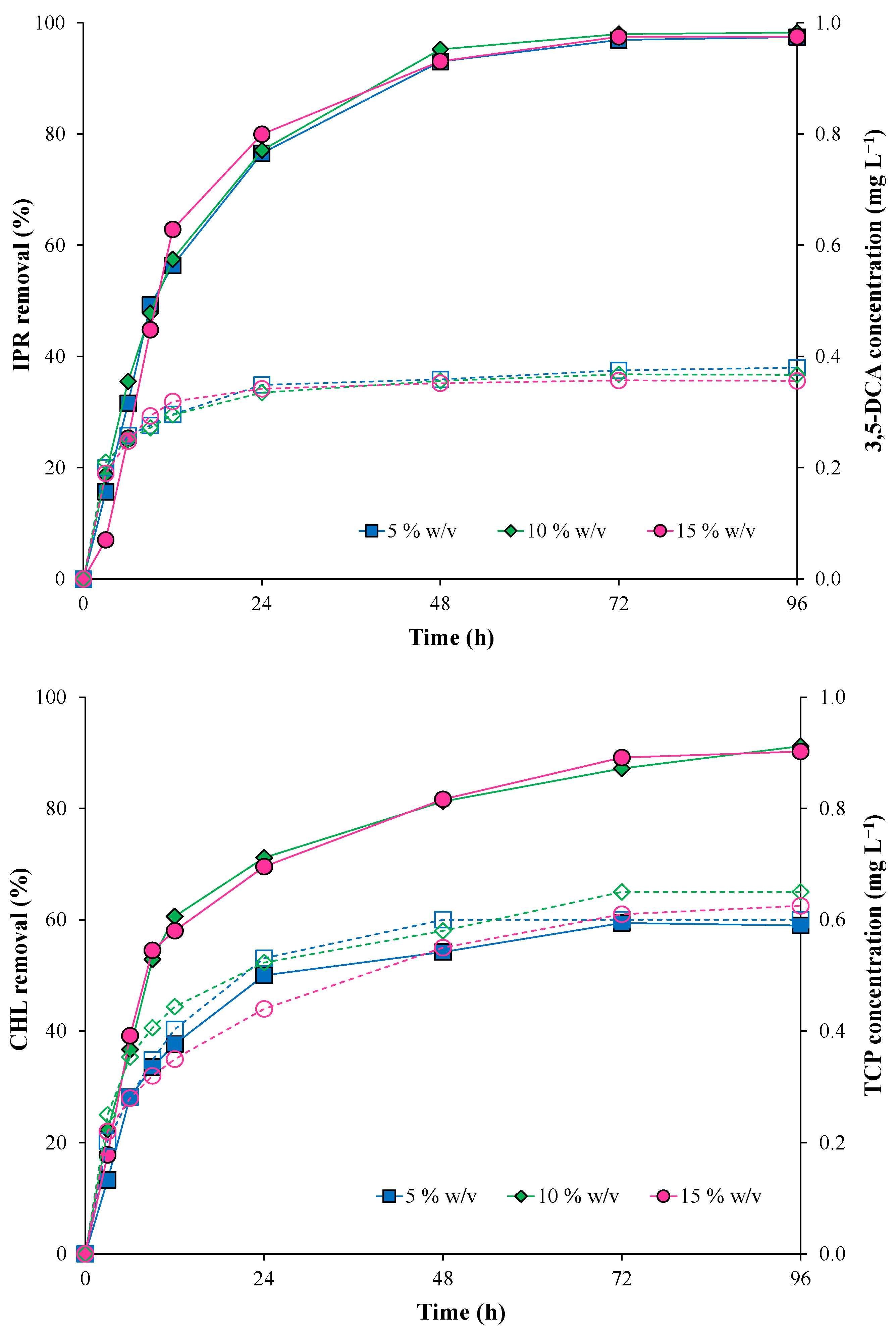
2.5. Pesticide Degradation by the Immobilized Bacterial Consortium in Batch Mode
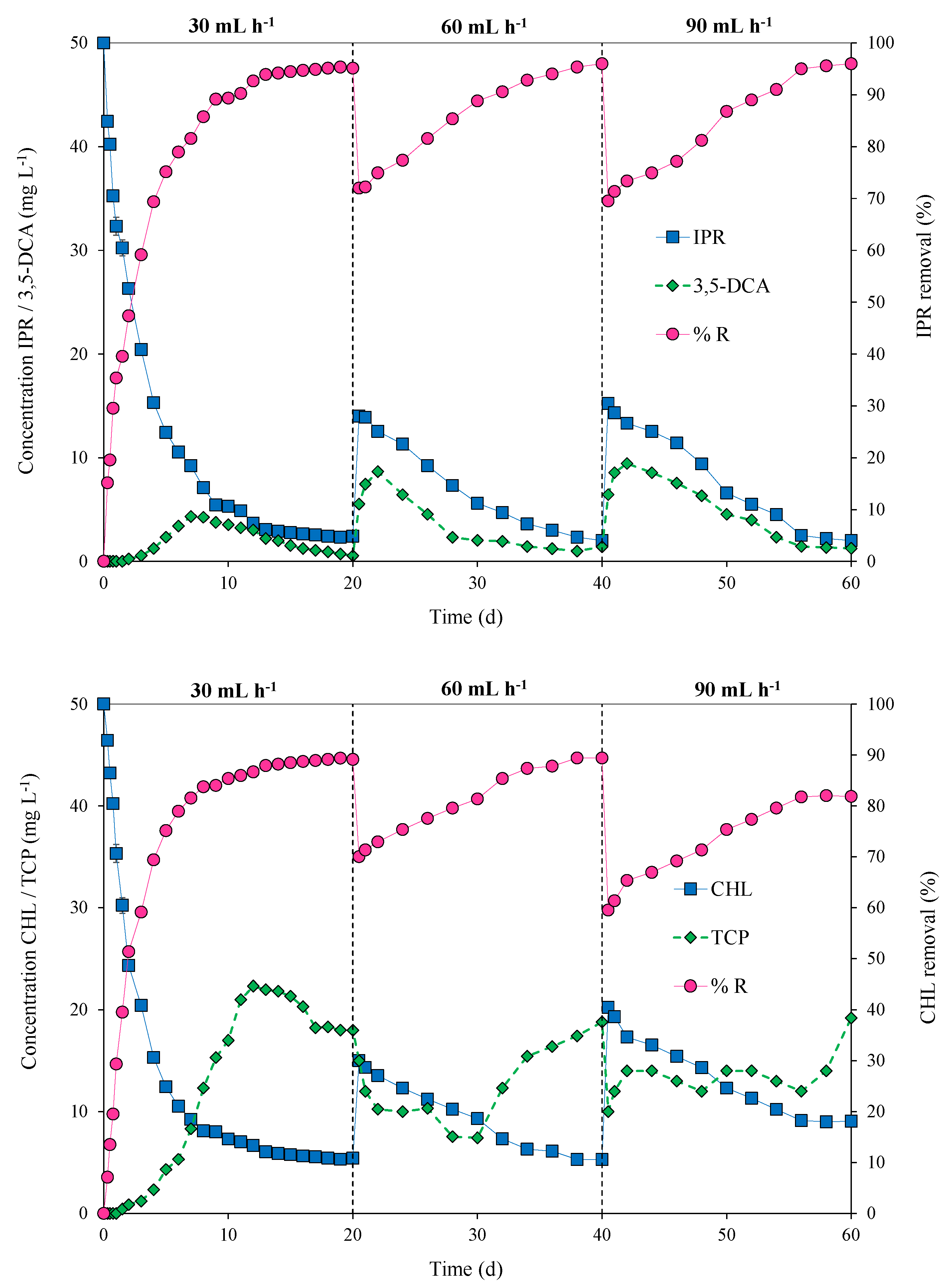
2.6. Pesticide Degradation by Immobilized Bacterial Consortium in Continuous Mode
2.7. Analyses of Pesticides and Metabolites
2.8. Kinetics and Statistical Analysis
3. Results
3.1. Pesticide Removal by Individual Bacterial Strains and Free Bacterial Consortium
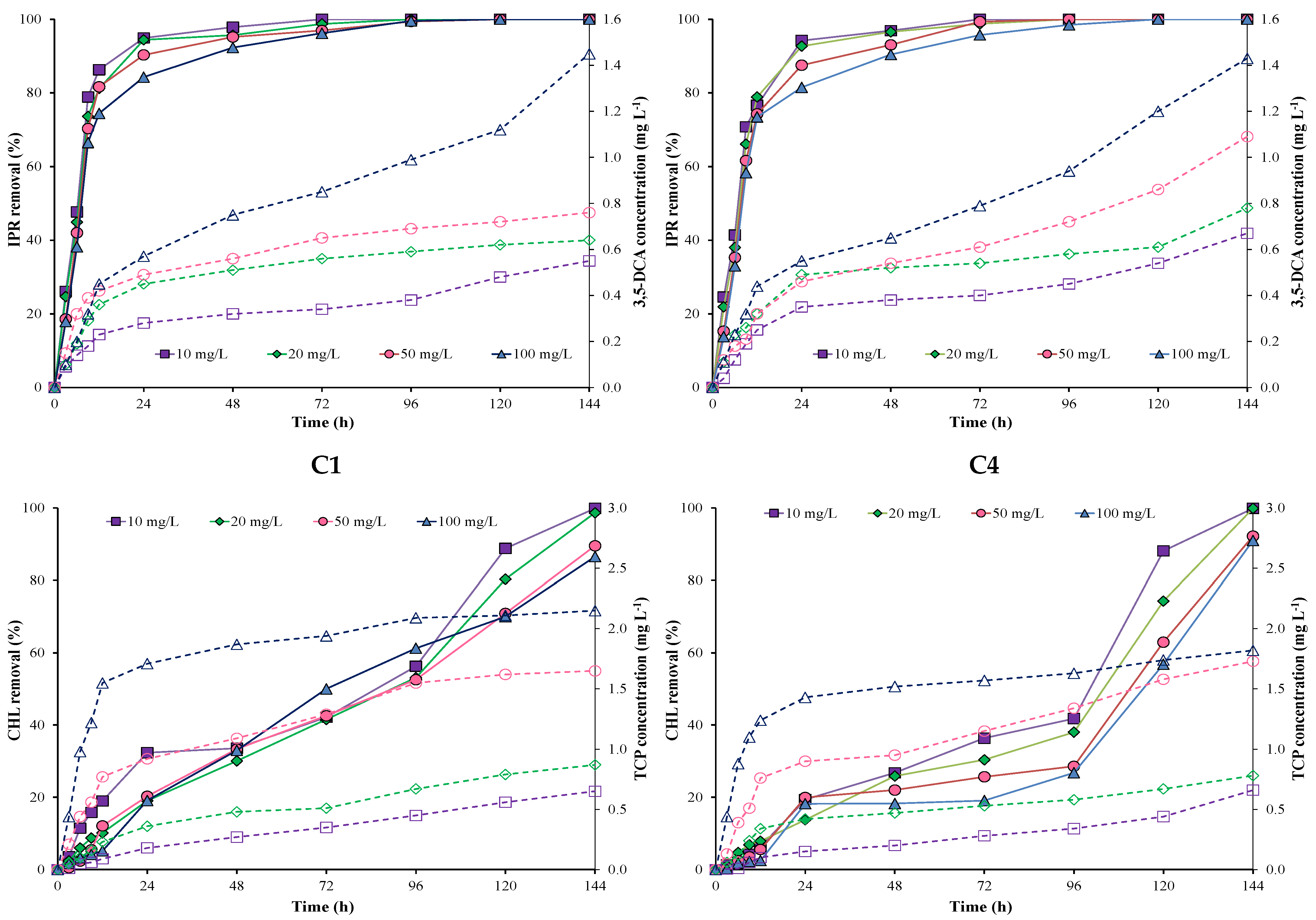
3.2. Pesticide Removal by Immobilized Bacterial Consortium
3.3. Pesticide Degradation via the Immobilized Bacterial Consortium in a Packed-Bed Bioreactor
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sabzevari, S.; Hofman, J. A worldwide review of currently used pesticides’ monitoring in agricultural soils. Sci. Total Environ. 2022, 812, 152344. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, G.; Atreya, K.; Scheepers, P.T.J.; Geissen, V. Concentration and distribution of pesticide residues in soil: Non-dietary human health risk assessment. Chemosphere 2020, 253, 126594. [Google Scholar] [CrossRef] [PubMed]
- Papazlatani, C.V.; Karas, P.A.; Lampronikou, E.; Karpouzas, D.G. Using biobeds for the treatment of fungicide-contaminated effluents from various agro-food processing industries: Microbiome responses and mobile genetic element dynamics. Sci. Total Environ. 2022, 823, 153744. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.L.; Zhang, J.; Liu, X.Z.; Tian, L.; Xiong, Z.S.; Xiong, X.; Chen, P.; Wu, D.S.; Zou, J.P. Degradation of pesticide wastewater with simultaneous resource recovery via ozonation coupled with anaerobic biochemical technology. Chemosphere 2022, 300, 134520. [Google Scholar] [CrossRef] [PubMed]
- Jatoi, A.S.; Hashmi, Z.; Adriyani, R.; Yuniarto, A.; Mazari, S.A.; Akhter, F.; Mubarak, N.M. Recent trends and future challenges of pesticide removal techniques—A comprehensive review. J. Environ. Chem. Eng. 2021, 9, 105571. [Google Scholar] [CrossRef]
- Castillo, M.D.P.; Torstensson, L.; Stenström, J. Biobeds for environmental protection from pesticide use—A review. J. Agric. Food Chem. 2008, 56, 6206–6219. [Google Scholar] [CrossRef]
- Karanasios, E.; Tsiropoulos, N.G.; Karpouzas, D.G. On-farm biopurification systems for the depuration of pesticide wastewaters: Recent biotechnological advances and future perspectives. Biodegradation 2012, 23, 787–802. [Google Scholar] [CrossRef]
- Diez, M.; Leiva, B.; Gallardo, F. Novel insights in biopurification system for dissipation of a pesticide mixture in repeated applications. Environ. Sci. Pollut. Res. 2018, 25, 21440–21450. [Google Scholar] [CrossRef]
- Diez, M.C.; Schalchli, H.; Elgueta, S.; Salgado, E.; Millahueque, N.; Rubilar, O.; Tortella, G.R.; Briceño, G. Rhizosphere effect on pesticide degradation in biobeds under different hydraulic loads. J. Soil Sci. Plant Nutr. 2015, 15, 410–421. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Rodríguez, C.E.; Cambronero-Heinrichs, J.C.; Castro-Gutiérrez, V.; Tortella, G.R. Biopurification Systems: Current advances and future prospects of on-farm biodegradation of pesticides. Handb. Environ. Chem. 2022, 113, 287–315. [Google Scholar] [CrossRef]
- Madrigal-Zúñiga, K.; Ruiz-Hidalgo, K.; Chin-Pampillo, J.S.; Masís-Mora, M.; Castro-Gutiérrez, V.; Rodríguez-Rodríguez, C.E. Fungal bioaugmentation of two rice husk-based biomixtures for the removal of carbofuran in on-farm biopurification systems. Biol. Fertil. Soils 2016, 52, 243–250. [Google Scholar] [CrossRef]
- Murillo-Zamora, S.; Castro-Gutiérrez, V.; Masís-Mora, M.; Lizano-Fallas, V.; Rodríguez-Rodríguez, C.E. Elimination of fungicides in biopurification systems: Effect of fungal bioaugmentation on removal performance and microbial community structure. Chemosphere 2017, 186, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Saez, J.M.; Bigliardo, A.L.; Raimondo, E.E.; Briceño, G.E.; Polti, M.A.; Benimeli, C.S. Lindane dissipation in a biomixture: Effect of soil properties and bioaugmentation. Ecotoxicol. Environ. Saf. 2018, 156, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Briceño, G.; Lamilla, C.; Leiva, B.; Levio, M.; Donoso-Piñol, P.; Schalchli, H.; Gallardo, F.; Diez, M.C. Pesticide-Tolerant bacteria isolated from a biopurification system to remove commonly used pesticides to protect water resources. PLoS ONE 2020, 15, e0234865. [Google Scholar] [CrossRef] [PubMed]
- Gangola, S.; Bhatt, P.; Kumar, A.J.; Bhandari, G.; Joshi, S.; Punetha, A.; Bhatt, K.; Rene, E.R. Biotechnological tools to elucidate the mechanism of pesticide degradation in the environment. Chemosphere 2022, 296, 133916. [Google Scholar] [CrossRef]
- Briceño, G.; Fuentes, M.S.; Palma, G.; Jorquera, M.A.; Amoroso, M.J.; Diez, M.C. Chlorpyrifos biodegradation and 3,5,6-trichloro-2-pyridinol production by actinobacteria isolated from soil. Int. Biodeterior. Biodegrad. 2012, 73, 1–7. [Google Scholar] [CrossRef]
- Briceño, G.; Schalchli, H.; Mutis, A.; Benimeli, C.S.; Palma, G.; Tortella, G.R.; Diez, M.C. Use of pure and mixed culture of diazinon-degrading Streptomyces to remove other organophosphorus pesticides. Int. Biodeterior. Biodegrad. 2016, 114, 193–201. [Google Scholar] [CrossRef]
- Campos, M.; Karas, P.S.; Perruchon, C.; Papadopoulou, E.S.; Christou, V.; Menkissoglou-Spiroudi, U.; Diez, M.C.; Karpouzas, D.G. Novel insights into the metabolic pathway of iprodione by soil bacteria. Environ. Sci. Pollut. Res. 2017, 24, 152–163. [Google Scholar] [CrossRef]
- John, E.M.; Sreekumar, J.; Jisha, M.S. Optimization of chlorpyrifos degradation by assembled bacterial consortium using response surface methodology. Soil Sediment Contam. 2016, 25, 668–682. [Google Scholar] [CrossRef]
- Fuentes, M.S.; Raimondo, E.E.; Amoroso, M.J.; Benimeli, C.S. Removal of a mixture of pesticides by a Streptomyces consortium: Influence of different soil systems. Chemosphere 2017, 173, 359–367. [Google Scholar] [CrossRef]
- Uniyal, S.; Sharma, R.K.; Kondakal, V. New insights into the biodegradation of chlorpyrifos by a novel bacterial consortium: Process optimization using general factorial experimental design. Ecotoxicol. Environ. Saf. 2021, 209, 111799. [Google Scholar] [CrossRef] [PubMed]
- Góngora-Echeverría, V.R.; García-Escalante, R.; Rojas-Herrera, R.; Giácoman-Vallejos, G.; Ponce-Caballero, C. Pesticide bioremediation in liquid media using a microbial consortium and bacteria-pure strains isolated from a biomixture used in agricultural areas. Ecotoxicol. Environ. Saf. 2020, 200, 110734. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, C.M. Efficiency of degrading packed bed reactors. In Proceedings of the 2016 WISA: International Conference on Information Security Applications, Jeju Islands, Republic of Korea, 25–27 August 2016. [Google Scholar]
- Marín-Benito, J.M.; Brown, C.D.; Herrero-Hernández, E.; Arienzo, M.; Sánchez-Martín, M.J.; Rodríguez-Cruz, M.S. Use of raw or incubated organic wastes as amendments in reducing pesticide leaching through soil columns. Sci. Total Environ. 2013, 463–464, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Deokar, S.K.; Mandavgane, S.A. Estimation of packed-bed parameters and prediction of breakthrough curves for adsorptive removal of 2,4-dichlorophenoxyacetic acid using rice husk ash. J. Environ. Chem. Eng. 2015, 3, 1827–1836. [Google Scholar] [CrossRef]
- Adhikari, S.; Chattopadhyay, P.; Ray, L. Continuous removal of malathion by immobilised biomass of Bacillus species S14 using a packed bed column reactor. Chem. Speciat. Bioavailab. 2012, 24, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Levio-Raiman, M.; Briceño, G.; Leiva, B.; López, S.; Schalchli, H.; Lamilla, C.; Bornhardt, C.; Diez, M.C. Treatment of pesticide-contaminated water using a selected fungal consortium: Study in a batch and packed-bed bioreactor. Agronomy 2021, 11, 743. [Google Scholar] [CrossRef]
- Geed, S.R.; Kureel, M.K.; Giri, B.S.; Singh, R.S.; Rai, B.N. Performance evaluation of Malathion biodegradation in batch and continuous packed bed bioreactor (PBBR). Bioresour. Technol. 2017, 227, 56–65. [Google Scholar] [CrossRef]
- Aswathi, A.; Pandey, A.; Madhavan, A.; Sukumaran, R.K. Chlorpyrifos induced proteome remodelling of Pseudomonas nitroreducens AR-3 potentially aid efficient degradation of the pesticide. Environ. Technol. Innov. 2021, 21, 101307. [Google Scholar] [CrossRef]
- Morillo, E.; Villaverde, J. Advanced technologies for the remediation of pesticide-contaminated soils. Sci. Total Environ. 2017, 586, 576–597. [Google Scholar] [CrossRef] [Green Version]
- Aswathi, A.; Pandey, A.; Sukumaran, R.K. Rapid degradation of the organophosphate pesticide—Chlorpyrifos by a novel strain of Pseudomonas nitroreducens AR-3. Bioresour. Technol. 2019, 292, 122025. [Google Scholar] [CrossRef]
- Carles, L.; Martin-Laurent, F.; Devers, M.; Spor, A.; Rouard, N.; Beguet, J.; Besse-Hoggan, P.; Batisson, I. Potential of preventive bioremediation to reduce environmental contamination by pesticides in an agricultural context: A case study with the herbicide 2,4-D. J. Hazard. Mater. 2021, 416, 125740. [Google Scholar] [CrossRef] [PubMed]
- Campos, M.; Perruchon, C.; Vasilieiadis, S.; Menkissoglu-Spiroudi, U.; Karpouzas, D.G.; Diez, M.C. Isolation and characterization of bacteria from acidic pristine soil environment able to transform iprodione and 3,5-dichloraniline. Int. Biodeterior. Biodegrad. 2015, 104, 201–211. [Google Scholar] [CrossRef]
- Castillo Diaz, J.M.; Delgado-Moreno, L.; Núñez, R.; Nogales, R.; Romero, E. Enhancing pesticide degradation using indigenous microorganisms isolated under high pesticide load in bioremediation systems with vermicomposts. Bioresour. Technol. 2016, 214, 234–241. [Google Scholar] [CrossRef]
- Mercadier, C.; Vega, D.; Bastide, J. Iprodione degradation by isolated soil microorganisms. FEMS Microbiol. Ecol. 1997, 23, 207–215. [Google Scholar] [CrossRef]
- Festa, S.; Coppotelli, B.M.; Morelli, I.S. Comparative bioaugmentation with a consortium and a single strain in a phenanthrene-contaminated soil: Impact on the bacterial community and biodegradation. Appl. Soil Ecol. 2016, 98, 8–19. [Google Scholar] [CrossRef]
- Yang, Z.; Jiang, W.; Wang, X.; Cheng, T.; Zhang, D.; Wang, H.; Qiu, J.; Cao, L.; Wang, X.; Hong, Q. An amidase gene, ipaH, is responsible for the initial step in the iprodione degradation pathway of Paenarthrobacter sp. strain YJN-5. Appl. Environ. Microbiol. 2018, 84, e01150-18. [Google Scholar] [CrossRef] [Green Version]
- Akbar, S.; Sultan, S. Soil bacteria showing a potential of chlorpyrifos degradation and plant growth enhancement. Braz. J. Microbiol. 2016, 47, 563–570. [Google Scholar] [CrossRef] [Green Version]
- Rayu, S.; Nielsen, U.N.; Nazaries, L.; Singh, B.K. Isolation and molecular characterization of novel chlorpyrifos and 3,5,6-trichloro-2-pyridinol-degrading bacteria from sugarcane farm soils. Front. Microbiol. 2017, 8, 518. [Google Scholar] [CrossRef] [Green Version]
- Chishti, Z.; Hussain, S.; Arshad, K.R.; Khalid, A.; Arshad, M. Microbial degradation of chlorpyrifos in liquid media and soil. J. Environ. Manag. 2013, 114, 372–380. [Google Scholar] [CrossRef]
- Bose, S.; Kumar, P.S.; Vo, D.V.N. A review on the microbial degradation of chlorpyrifos and its metabolite TCP. Chemosphere 2021, 283, 131447. [Google Scholar] [CrossRef]
- Briceño, G.; Vergara, K.; Schalchli, H.; Palma, G.; Tortella, G.; Fuentes, M.S.; Diez, M.C. Organophosphorus pesticide mixture removal from environmental matrices by a soil Streptomyces mixed culture. Environ. Sci. Pollut. Res. 2018, 25, 21296–21307. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Pang, S.; Zhou, Z.; Wu, X.; Li, J.; Huang, Y.; Zhang, W.; Lei, Q.; Bhatt, P.; Mishra, S.; et al. Novel pathway of acephate degradation by the microbial consortium ZQ01 and its potential for environmental bioremediation. J. Hazard. Mater. 2022, 426, 127841. [Google Scholar] [CrossRef] [PubMed]
- Abraham, J.; Silambarasan, S.; Logeswari, P. Simultaneous degradation of organophosphorus and organochlorine pesticides by bacterial consortium. J. Taiwan Inst. Chem. Eng. 2014, 45, 2590–2596. [Google Scholar] [CrossRef]
- Xu, C.; Yu, H. Insights into constructing a stable and efficient microbial consortium. Chin. J. Chem. Eng. 2021, 30, 112–120. [Google Scholar] [CrossRef]
- Banerjee, A.; Ghoshal, A.K. Biodegradation of phenol by calcium-alginate immobilized Bacillus cereus in a packed bed reactor and determination of the mass transfer correlation. J. Environ. Chem. Eng. 2016, 4, 1523–1529. [Google Scholar] [CrossRef]
- Tallur, P.N.; Mulla, S.I.; Megadi, V.B.; Talwar, M.P.; Ninnekar, H.Z. Biodegradation of cypermethrin by immobilized cells of micrococcus sp. Strain CPN 1. Braz. J. Microbiol. 2015, 46, 667–672. [Google Scholar] [CrossRef] [Green Version]
- Mohanty, S.S.; Jena, H.M. Degradation kinetics and mechanistic study on herbicide bioremediation using hyper butachlor-tolerant Pseudomonas putida G3. Process Saf. Environ. Prot. 2019, 125, 172–181. [Google Scholar] [CrossRef]
- Briceño, G.; Fuentes, M.S.; Rubilar, O.; Jorquera, M.; Tortella, G.; Palma, G.; Amoroso, M.J.; Diez, M.C. Removal of the insecticide diazinon from liquid media by free and immobilized Streptomyces sp. isolated from agricultural soil. J. Basic Microbiol. 2015, 55, 293–302. [Google Scholar] [CrossRef]
- Zilouei, H.; Guieysse, B.; Mattiasson, B. Biological degradation of chlorophenols in packed-bed bioreactors using mixed bacterial consortia. Process Biochem. 2006, 41, 1083–1089. [Google Scholar] [CrossRef]
- Yadav, M.; Srivastva, N.; Singh, R.S.; Upadhyay, S.N.; Dubey, S.K. Biodegradation of chlorpyrifos by Pseudomonas sp. in a continuous packed bed bioreactor. Bioresour. Technol. 2014, 165, 265–269. [Google Scholar] [CrossRef]
- Yadav, M.; Srivastva, N.; Shukla, A.K.; Singh, R.S.; Upadhyay, S.N.; Dubey, S.K. Efficacy of Aspergillus sp. for degradation of chlorpyrifos in batch and continuous aerated packed bed bioreactors. Appl. Biochem. Biotechnol. 2015, 175, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhang, W.; Pang, S.; Chen, J.; Bhatt, P.; Mishra, S.; Chen, S. Insights into the microbial degradation and catalytic mechanisms of chlorpyrifos. Environ. Res. 2021, 194, 110660. [Google Scholar] [CrossRef] [PubMed]
- Khalid, S.; Hashmi, I. Biotreatment of chlorpyrifos in a bench scale bioreactor using Psychrobacter alimentarius T14. Environ. Technol. 2016, 37, 316–325. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Iprodione | Chlorpyrifos |
|---|---|---|
| Chemical class | Dicarboximide | Organophosphate |
| Molecular formula | C9H11Cl3NO3PS | C13H13Cl2N3O3 |
| Chemical structure |  |  |
| Water solubility (mg L−1) | 6.80 | 1.05 |
| Molecular weight (g mol−1) | 330 | 351 |
| T1/2 (d) in soils | 36 | 50 |
| T1/2 (d) in biomixture * | 15.5 | 19.8 |
| GUS | 2.7 | 0.17 |
| Kow (Log P) | 3.0 | 4.7 |
| Koc | 700 | 8100 |
| C1 | C4 | Free Bacterial Consortium | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Pesticide | Concentration (mg L−1) | k (h−1) | T1/2 (h) | R2 | k (h−1) | T1/2 (h) | R2 | k (h−1) | T1/2 (h) | R2 |
| IPR | 10 | 0.11 ± 0.01 | 6.27 ± 0.10 a | 0.998 | 0.12 ± 0.03 | 12.40 ± 0.02 a | 0.985 | 0.29 ± 0.00 | 8.81 ± 0.11 a | 0.960 |
| 20 | 0.11 ± 0.01 | 6.40 ± 0.20 a | 0.978 | 0.11± 0.02 | 12.93 ± 0.01 a | 0.994 | 0.28 ± 0.01 | 8.74 ± 0.21 a | 0.997 | |
| 50 | 0.16 ± 0.02 | 4.29 ± 0.20 b | 0.999 | 0.09 ± 0.01 | 8.49 ± 0.05 b | 0.995 | 0.25 ± 0.02 | 8.63 ± 0.01 a | 0.965 | |
| 100 | 0.10 ± 0.01 | 7.11 ± 0.10 a | 0.987 | 0.07 ± 0.01 | 12.77 ± 0.02 a | 0.982 | 0.20 ± 0.00 | 8.71 ± 0.02 a | 0.991 | |
| CHL | 10 | 0.01 ± 0.001 | 112.56 ± 2.40 a | 0.981 | 0.01 ± 0.001 | 145.68 ± 2.64 a | 0.992 | 0.17 ± 0.01 | 10.56 ± 1.88 a | 0.975 |
| 20 | 0.01 ± 0.001 | 109.92 ± 2.88 a | 0.983 | 0.02 ± 0.001 | 170.16 ± 5.52 a | 0.990 | 0.20 ± 0.01 | 10.08 ± 1.42 a | 0.991 | |
| 50 | 0.02 ± 0.001 | 198.24 ± 3.60 b | 0.994 | 0.01 ± 0.001 | 259.42 ± 2.88 b | 0.998 | 0.16 ± 0.01 | 12.96 ± 1.79 a | 0.962 | |
| 100 | 0.01 ± 0.001 | 231.12 ± 2.64 b | 0.998 | 0.01 ± 0.001 | 277.44 ± 2.40 b | 0.991 | 0.17 ± 0.01 | 12.24 ± 1.98 a | 0.955 | |
| IPR | CHL | |||||
|---|---|---|---|---|---|---|
| Inoculum Concentration (% w/v) | k (h−1) | T1/2 (h) | R2 | k (h−1) | T1/2 (h) | R2 |
| 5 | 0.14 ± 0.02 | 11.78 ± 0.01 a | 0.997 | 0.09 ± 0.000 | 24.42 ± 0.02 a | 0.996 |
| 10 | 0.15 ± 0.01 | 11.51 ± 0.02 a | 0.998 | 0.10 ± 0.001 | 9.29 ± 0.10 b | 0.991 |
| 15 | 0.16 ± 0.03 | 10.78 ± 0.01 a | 0.995 | 0.12 ± 0.001 | 9.10 ± 0.05 b | 0.998 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Levío-Raimán, M.; Bornhardt, C.; Diez, M.C. Biodegradation of Iprodione and Chlorpyrifos Using an Immobilized Bacterial Consortium in a Packed-Bed Bioreactor. Microorganisms 2023, 11, 220. https://doi.org/10.3390/microorganisms11010220
Levío-Raimán M, Bornhardt C, Diez MC. Biodegradation of Iprodione and Chlorpyrifos Using an Immobilized Bacterial Consortium in a Packed-Bed Bioreactor. Microorganisms. 2023; 11(1):220. https://doi.org/10.3390/microorganisms11010220
Chicago/Turabian StyleLevío-Raimán, Marcela, Cristian Bornhardt, and M. Cristina Diez. 2023. "Biodegradation of Iprodione and Chlorpyrifos Using an Immobilized Bacterial Consortium in a Packed-Bed Bioreactor" Microorganisms 11, no. 1: 220. https://doi.org/10.3390/microorganisms11010220
APA StyleLevío-Raimán, M., Bornhardt, C., & Diez, M. C. (2023). Biodegradation of Iprodione and Chlorpyrifos Using an Immobilized Bacterial Consortium in a Packed-Bed Bioreactor. Microorganisms, 11(1), 220. https://doi.org/10.3390/microorganisms11010220