
Arthropod-Borne Flaviviruses in Pregnancy
 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
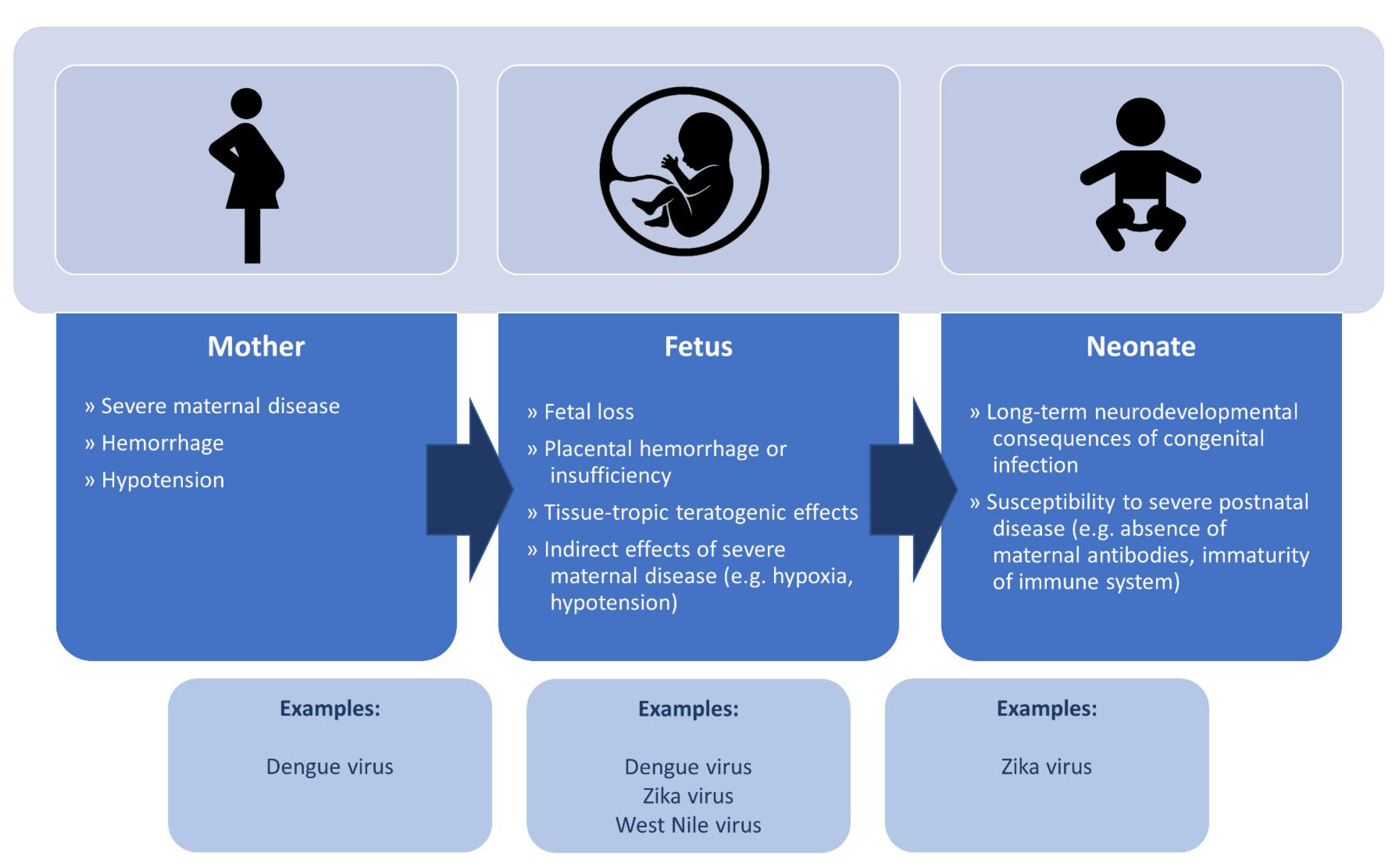
2. Overarching Pathophysiology
3. Zika Virus
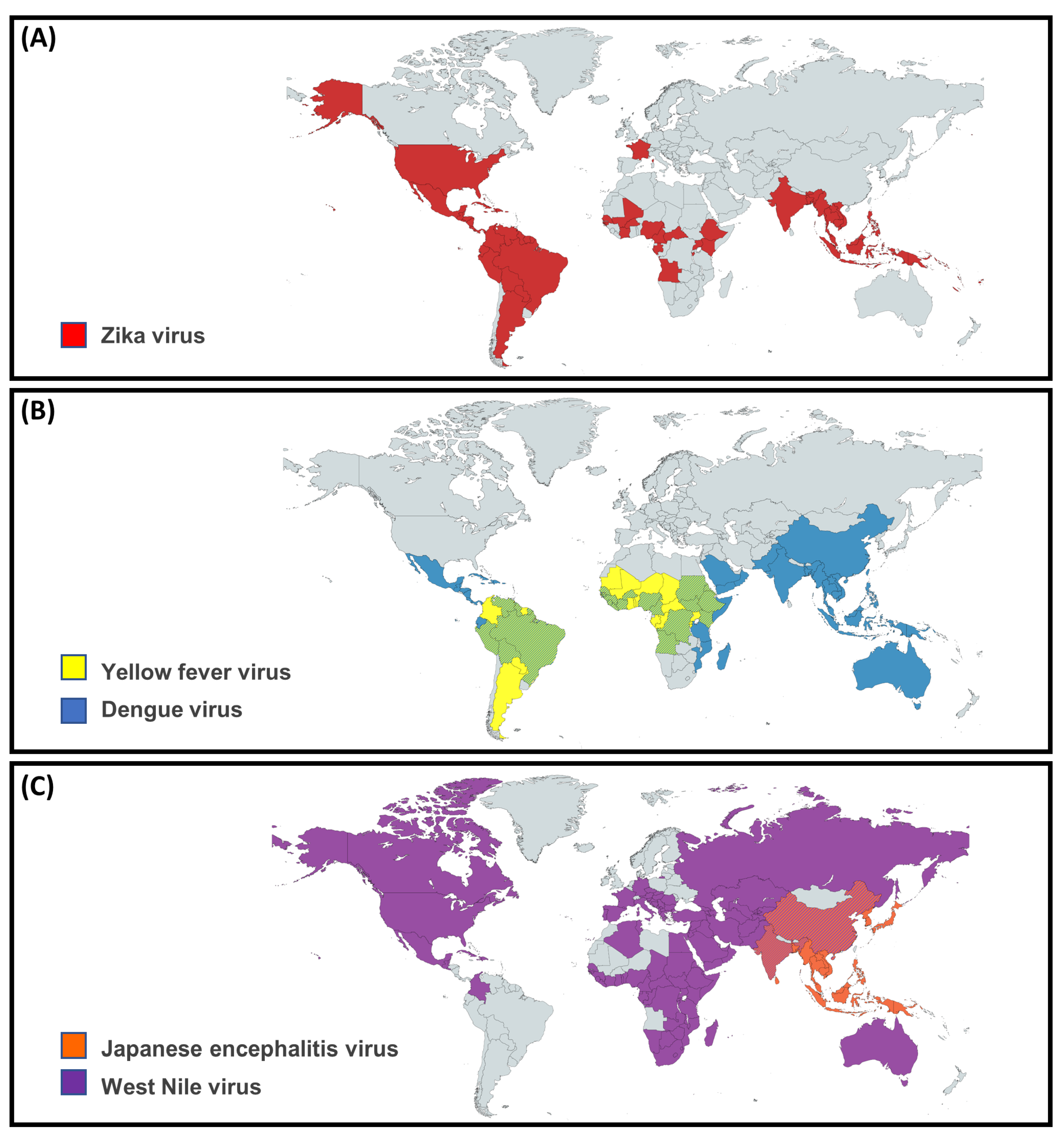
3.1. Epidemiology
3.2. Pathogenesis
3.3. Clinical Features
3.3.1. Pregnant Women
3.3.2. Fetus
3.3.3. Neonate
4. Dengue Virus
4.1. Epidemiology
4.2. Pathogenesis
4.3. Clinical Features
4.3.1. Pregnant Women
4.3.2. Fetus
4.3.3. Neonate
5. Japanese Encephalitis Virus
5.1. Epidemiology
5.2. Pathogenesis
5.3. Clinical Features
5.3.1. Pregnant Women
5.3.2. Fetus
5.3.3. Neonate
6. West Nile Virus
6.1. Epidemiology
6.2. Pathogenesis
6.3. Clinical Features
6.3.1. Pregnant Women
6.3.2. Fetus
6.3.3. Neonate
7. Yellow Fever Virus
7.1. Epidemiology
7.2. Pathogenesis
7.3. Clinical Features
7.3.1. Pregnant Women
7.3.2. Fetus
7.3.3. Neonate
8. Other Flaviviruses in Pregnancy
9. Laboratory Diagnosis
9.1. Serology
9.2. Molecular Testing
9.3. Other Modalities
9.4. Integration of Diagnostic Results
10. Treatment of Flavivirus Infections
Specific Considerations in Dengue Virus Infection
11. Prevention of and Vaccination for Flavivirus Infections
11.1. Behavioural, Environmental, and Infection Control Strategies
11.2. Vaccination for Flaviviruses
12. Arthropod Vectors Responsible for Flavivirus Transmission
13. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CI | confidence interval |
| CNS | central nervous system |
| CSF | cerebrospinal fluid |
| CZS | congenital Zika syndrome |
| DENV | Dengue virus |
| DHF | dengue hemorrhagic fever |
| DNA | deoxyribonucleic acid |
| DSS | dengue shock syndrome |
| GBS | Guillain-Barré syndrome |
| JE | Japanese encephalitis |
| JEV | Japanese encephalitis virus |
| LAMP | loop-mediated isothermal amplification |
| NAAT | nucleic acid amplification test |
| NS1 | non-structural antigen 1 |
| OR | odds ratio |
| PC3 | physical containment level 3 |
| RNA | ribonucleic acid |
| WNV | West Nile virus |
| YFV | Yellow fever virus |
| ZIKV | Zika virus |
References
- Pierson, T.C.; Diamond, M.S. The continued threat of emerging flaviviruses. Nat. Microbiol. 2020, 5, 796–812. [Google Scholar] [CrossRef] [PubMed]
- Chong, H.Y.; Leow, C.Y.; Abdul Majeed, A.B.; Leow, C.H. Flavivirus infection—A review of immunopathogenesis, immunological response, and immunodiagnosis. Virus Res. 2019, 274, 197770. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. West Nile Virus Infections in EU/EEA and EU-Neighbouring Countries; ECDC: Solna, Sweden, 1 February 2022; Available online: https://gis.ecdc.europa.eu/portal/apps/opsdashboard/index.html#/305f1b279cb04dddac3d4833b770a620 (accessed on 4 February 2022).
- World Health Organization. Countries and Territories with Current or Previous Zika Virus Transmission; World Health Organization: Geneva, Switzerland, February 2022; Available online: https://cdn.who.int/media/docs/default-source/documents/emergencies/zika/map-of-countries_with_zika_transmission_feb2022.pdf?sfvrsn=802a352a_5 (accessed on 4 February 2022).
- Howard-Jones, A.R.; Pham, D.; Jeoffreys, N.; Eden, J.-S.; Hueston, L.; Kesson, A.M.; Nagendra, V.; Samarasekara, H.; Newton, P.; Chen, S.C.A.; et al. Emerging Genotype IV Japanese Encephalitis Virus Outbreak in New South Wales, Australia. Viruses 2022, 14, 1853. [Google Scholar] [CrossRef] [PubMed]
- Endy, T.P.; Chunsuttiwat, S.; Nisalak, A.; Libraty, D.H.; Green, S.; Rothman, A.L.; Vaughn, D.W.; Ennis, F.A. Epidemiology of inapparent and symptomatic acute dengue virus infection: A prospective study of primary school children in Kamphaeng Phet, Thailand. Am. J. Epidemiol. 2002, 156, 40–51. [Google Scholar] [CrossRef]
- Mostashari, F.; Bunning, M.L.; Kitsutani, P.T.; Singer, D.A.; Nash, D.; Cooper, M.J.; Katz, N.; Liljebjelke, K.A.; Biggerstaff, B.J.; Fine, A.D.; et al. Epidemic West Nile encephalitis, New York, 1999: Results of a household-based seroepidemiological survey. Lancet 2001, 358, 261–264. [Google Scholar] [CrossRef]
- Rosen, L. The natural history of Japanese encephalitis virus. Annu. Rev. Microbiol. 1986, 40, 395–414. [Google Scholar] [CrossRef]
- Chan-Tack, K.M.; Forrest, G. West Nile virus meningoencephalitis and acute flaccid paralysis after infliximab treatment. J. Rheumatol. 2006, 33, 191–192. [Google Scholar]
- Charlier, C.; Beaudoin, M.C.; Couderc, T.; Lortholary, O.; Lecuit, M. Arboviruses and pregnancy: Maternal, fetal, and neonatal effects. Lancet Child Adolesc. Health 2017, 1, 134–146. [Google Scholar] [CrossRef]
- Pardigon, N. Pathophysiological mechanisms of Flavivirus infection of the central nervous system. Transfus. Clin. Biol. 2017, 24, 96–100. [Google Scholar] [CrossRef]
- Osuna-Ramos, J.F.; Reyes-Ruiz, J.M.; Del Ángel, R.M. The Role of Host Cholesterol During Flavivirus Infection. Front. Cell. Infect. Microbiol. 2018, 8, 388. [Google Scholar] [CrossRef]
- Peruzzu, D.; Amendola, A.; Venturi, G.; de Turris, V.; Marsili, G.; Fortuna, C.; Fecchi, K.; Gagliardi, M.C. Zika Virus Exploits Lipid Rafts to Infect Host Cells. Viruses 2022, 14, 2059. [Google Scholar] [CrossRef] [PubMed]
- Platt, D.J.; Smith, A.M.; Arora, N.; Diamond, M.S.; Coyne, C.B.; Miner, J.J. Zika virus-related neurotropic flaviviruses infect human placental explants and cause fetal demise in mice. Sci. Transl. Med. 2018, 10, eaao7090. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Zika Travel Information; Centers for Disease Control and Prevention (CDC): Atlanta, GA, USA, 2022. Available online: https://wwwnc.cdc.gov/travel/page/zika-information (accessed on 4 February 2022).
- Bogoch, I.I.; Brady, O.J.; Kraemer, M.U.G.; German, M.; Creatore, M.I.; Kulkarni, M.A.; Brownstein, J.S.; Mekaru, S.R.; Hay, S.I.; Groot, E.; et al. Anticipating the international spread of Zika virus from Brazil. Lancet 2016, 387, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Centeno-Tablante, E.; Medina-Rivera, M.; Finkelstein, J.L.; Herman, H.S.; Rayco-Solon, P.; Garcia-Casal, M.N.; Rogers, L.; Ghezzi-Kopel, K.; Zambrano Leal, M.P.; Andrade Velasquez, J.K.; et al. Update on the Transmission of Zika Virus Through Breast Milk and Breastfeeding: A Systematic Review of the Evidence. Viruses 2021, 13, 123. [Google Scholar] [CrossRef]
- Venturi, G.; Zammarchi, L.; Fortuna, C.; Remoli, M.E.; Benedetti, E.; Fiorentini, C.; Trotta, M.; Rizzo, C.; Mantella, A.; Rezza, G.; et al. An autochthonous case of Zika due to possible sexual transmission, Florence, Italy, 2014. Eurosurveillance 2016, 21, 30148. [Google Scholar] [CrossRef]
- Musso, D.; Nhan, T.; Robin, E.; Roche, C.; Bierlaire, D.; Zisou, K.; Shan Yan, A.; Cao-Lormeau, V.M.; Broult, J. Potential for Zika virus transmission through blood transfusion demonstrated during an outbreak in French Polynesia, November 2013 to February 2014. Eurosurveillance 2014, 19, 20761. [Google Scholar] [CrossRef]
- Brasil, P.; Calvet, G.A.; Siqueira, A.M.; Wakimoto, M.; de Sequeira, P.C.; Nobre, A.; Quintana Mde, S.; Mendonça, M.C.; Lupi, O.; de Souza, R.V.; et al. Zika Virus Outbreak in Rio de Janeiro, Brazil: Clinical Characterization, Epidemiological and Virological Aspects. PLoS Negl. Trop. Dis. 2016, 10, e0004636. [Google Scholar] [CrossRef]
- Edupuganti, S.; Natrajan, M.S.; Rouphael, N.; Lai, L.; Xu, Y.; Feldhammer, M.; Hill, C.; Patel, S.M.; Johnson, S.J.; Bower, M.; et al. Biphasic Zika Illness with Rash and Joint Pain. Open Forum Infect. Dis. 2017, 4, ofx133. [Google Scholar] [CrossRef]
- Parra, B.; Lizarazo, J.; Jiménez-Arango, J.A.; Zea-Vera, A.F.; González-Manrique, G.; Vargas, J.; Angarita, J.A.; Zuñiga, G.; Lopez-Gonzalez, R.; Beltran, C.L.; et al. Guillain-Barré Syndrome Associated with Zika Virus Infection in Colombia. N. Engl. J. Med. 2016, 375, 1513–1523. [Google Scholar] [CrossRef]
- da Silva, I.R.F.; Frontera, J.A.; Bispo de Filippis, A.M.; Nascimento, O. Neurologic Complications Associated with the Zika Virus in Brazilian Adults. JAMA Neurol. 2017, 74, 1190–1198. [Google Scholar] [CrossRef]
- Flamand, C.; Fritzell, C.; Matheus, S.; Dueymes, M.; Carles, G.; Favre, A.; Enfissi, A.; Adde, A.; Demar, M.; Kazanji, M.; et al. The proportion of asymptomatic infections and spectrum of disease among pregnant women infected by Zika virus: Systematic monitoring in French Guiana, 2016. Eurosurveillance 2017, 22, 1190–1198. [Google Scholar] [CrossRef] [PubMed]
- Brasil, P.; Pereira, J.P., Jr.; Moreira, M.E.; Ribeiro Nogueira, R.M.; Damasceno, L.; Wakimoto, M.; Rabello, R.S.; Valderramos, S.G.; Halai, U.A.; Salles, T.S.; et al. Zika Virus Infection in Pregnant Women in Rio de Janeiro. N. Engl. J. Med. 2016, 375, 2321–2334. [Google Scholar] [CrossRef] [PubMed]
- Brasil, P.; Vasconcelos, Z.; Kerin, T.; Gabaglia, C.R.; Ribeiro, I.P.; Bonaldo, M.C.; Damasceno, L.; Pone, M.V.; Pone, S.; Zin, A.; et al. Zika virus vertical transmission in children with confirmed antenatal exposure. Nat. Commun. 2020, 11, 3510. [Google Scholar] [CrossRef] [PubMed]
- Pomar, L.; Vouga, M.; Lambert, V.; Pomar, C.; Hcini, N.; Jolivet, A.; Benoist, G.; Rousset, D.; Matheus, S.; Malinger, G.; et al. Maternal-fetal transmission and adverse perinatal outcomes in pregnant women infected with Zika virus: Prospective cohort study in French Guiana. BMJ 2018, 363, k4431. [Google Scholar] [CrossRef]
- Alvarado-Domenech, L.I.; Rivera-Amill, V.; Appleton, A.A.; Rosario-Villafañe, V.; Repollet-Carrer, I.; Borges-Rodríguez, M.; Pérez-Rodríguez, N.M.; Olivieri-Ramos, O.; González, M.; González-Montalvo, C.; et al. Early Childhood Neurodevelopmental Outcomes in Children with Prenatal Zika Virus Exposure: A Cohort Study in Puerto Rico. J. Pediatr. 2022, 247, 38–45. [Google Scholar] [CrossRef]
- Ventura, C.V.; Ventura, L.O. Ophthalmologic Manifestations Associated with Zika Virus Infection. Pediatrics 2018, 141 (Suppl. 2), S161–S166. [Google Scholar] [CrossRef]
- Vouga, M.; Baud, D. Imaging of congenital Zika virus infection: The route to identification of prognostic factors. Prenat. Diagn. 2016, 36, 799–811. [Google Scholar] [CrossRef]
- Freitas, D.A.; Souza-Santos, R.; Carvalho, L.M.A.; Barros, W.B.; Neves, L.M.; Brasil, P.; Wakimoto, M.D. Congenital Zika syndrome: A systematic review. PLoS ONE 2020, 15, e0242367. [Google Scholar] [CrossRef]
- Hcini, N.; Kugbe, Y.; Rafalimanana, Z.H.L.; Lambert, V.; Mathieu, M.; Carles, G.; Baud, D.; Panchaud, A.; Pomar, L. Association between confirmed congenital Zika infection at birth and outcomes up to 3 years of life. Nat. Commun. 2021, 12, 3270. [Google Scholar] [CrossRef]
- Pimentel, R.; Khosla, S.; Rondon, J.; Peña, F.; Sullivan, G.; Perez, M.; Mehta, S.D.; Brito, M.O. Birth Defects and Long-Term Neurodevelopmental Abnormalities in Infants Born During the Zika Virus Epidemic in the Dominican Republic. Ann. Glob. Health 2021, 87, 4. [Google Scholar] [CrossRef]
- Australasian Society for Infectious Diseases. Management of Perinatal Infections, 3rd ed.; Palasanthiran, P., Starr, M., Jones, C., Giles, M., Eds.; Australasian Society for Infectious Diseases: Sydney, Australia, 2022. [Google Scholar]
- Maness, N.J.; Schouest, B.; Singapuri, A.; Dennis, M.; Gilbert, M.H.; Bohm, R.P.; Schiro, F.; Aye, P.P.; Baker, K.; Van Rompay, K.K.A.; et al. Postnatal Zika virus infection of nonhuman primate infants born to mothers infected with homologous Brazilian Zika virus. Sci. Rep. 2019, 9, 12802. [Google Scholar] [CrossRef] [PubMed]
- Healy, C.M.; Rench, M.A.; Swaim, L.S.; Timmins, A.; Vyas, A.; Sangi-Haghpeykar, H.; Ng, N.; Rajam, G.; Havers, F.; Schiffer, J.; et al. Kinetics of maternal pertussis-specific antibodies in infants of mothers vaccinated with tetanus, diphtheria and acellular pertussis (Tdap) during pregnancy. Vaccine 2020, 38, 5955–5961. [Google Scholar] [CrossRef] [PubMed]
- Nyiro, J.U.; Sande, C.; Mutunga, M.; Kiyuka, P.K.; Munywoki, P.K.; Scott, J.A.; Nokes, D.J. Quantifying maternally derived respiratory syncytial virus specific neutralising antibodies in a birth cohort from coastal Kenya. Vaccine 2015, 33, 1797–1801. [Google Scholar] [CrossRef] [PubMed]
- Bentes, A.A.; Crispim, A.P.C.; Marinho, P.E.S.; Viegas, E.C.C.; Loutfi, K.S.; Guedes, I.; Araujo, S.T.; Alvarenga, A.M.; Campos, E.S.L.M.; Santos, M.A.; et al. Neurologic Manifestations of Noncongenital Zika Virus in Children. J. Pediatr. 2021, 237, 298–301. [Google Scholar] [CrossRef] [PubMed]
- Shepard, D.S.; Undurraga, E.A.; Halasa, Y.A.; Stanaway, J.D. The global economic burden of dengue: A systematic analysis. Lancet Infect. Dis. 2016, 16, 935–941. [Google Scholar] [CrossRef]
- Hales, S.; de Wet, N.; Maindonald, J.; Woodward, A. Potential effect of population and climate changes on global distribution of dengue fever: An empirical model. Lancet 2002, 360, 830–834. [Google Scholar] [CrossRef]
- Guzman, M.G.; Harris, E. Dengue. Lancet 2015, 385, 453–465. [Google Scholar] [CrossRef]
- Henchal, E.A.; Putnak, J.R. The dengue viruses. Clin. Microbiol. Rev. 1990, 3, 376–396. [Google Scholar] [CrossRef]
- Simmons, C.P.; Farrar, J.J.; van Vinh Chau, N.; Wills, B. Dengue. N. Engl. J. Med. 2012, 366, 1423–1432. [Google Scholar] [CrossRef]
- Kuno, G. Review of the Factors Modulating Dengue Transmission. Epidemiol. Rev. 1995, 17, 321–335. [Google Scholar] [CrossRef]
- Vaughn, D.W.; Green, S.; Kalayanarooj, S.; Innis, B.L.; Nimmannitya, S.; Suntayakorn, S.; Rothman, A.L.; Ennis, F.A.; Nisalak, A. Dengue in the Early Febrile Phase: Viremia and Antibody Responses. J. Infect. Dis. 1997, 176, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Halstead, S.B. Dengue Antibody-Dependent Enhancement: Knowns and Unknowns. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Katzelnick, L.C.; Gresh, L.; Halloran, M.E.; Mercado, J.C.; Kuan, G.; Gordon, A.; Balmaseda, A.; Harris, E. Antibody-dependent enhancement of severe dengue disease in humans. Science 2017, 358, 929–932. [Google Scholar] [CrossRef] [PubMed]
- Libraty, D.H.; Endy, T.P.; Houng, H.; Shu, H.; Green, S.; Kalayanarooj, S.; Suntayakorn, S.; Chansiriwongs, W.; Vaughn, D.W.; Nisalak, A.; et al. Differing Influences of Virus Burden and Immune Activation on Disease Severity in Secondary Dengue-3 Virus Infections. J. Infect. Dis. 2002, 185, 1213–1221. [Google Scholar] [CrossRef] [PubMed]
- Rothman, A.L. Immunity to dengue virus: A tale of original antigenic sin and tropical cytokine storms. Nat. Rev. Immunol. 2011, 11, 532–543. [Google Scholar] [CrossRef] [PubMed]
- Endy, T.P.; Srikiatkhachorn, A.; Jarman, R.G.; Mammen, M.P.; Vaughn, D.W.; Kalanarooj, S.; Nisalak, A.; Gibbons, R.V. Analysis of Repeat Hospital Admissions for Dengue to Estimate the Frequency of Third or Fourth Dengue Infections Resulting in Admissions and Dengue Hemorrhagic Fever, and Serotype Sequences. Am. J. Trop. Med. Hyg. 2007, 77, 910–913. [Google Scholar] [CrossRef]
- Martina, B.E.E.; Koraka, P.; Osterhaus, A.D.M.E. Dengue Virus Pathogenesis: An Integrated View. Clin. Microbiol. Rev. 2009, 22, 564–581. [Google Scholar] [CrossRef]
- Beatty, P.R.; Puerta-Guardo, H.; Killingbeck, S.S.; Glasner, D.R.; Hopkins, K.; Harris, E. Dengue virus NS1 triggers endothelial permeability and vascular leak that is prevented by NS1 vaccination. Sci. Transl. Med. 2015, 7, 304ra141. [Google Scholar] [CrossRef]
- Nakao, S.; Lai, C.J.; Young, N.S. Dengue virus, a flavivirus, propagates in human bone marrow progenitors and hematopoietic cell lines. Blood 1989, 74, 1235–1240. [Google Scholar] [CrossRef]
- Machado, C.R.; Machado, E.S.; Rohloff, R.D.; Azevedo, M.; Campos, D.P.; de Oliveira, R.B.; Brasil, P. Is Pregnancy Associated with Severe Dengue? A Review of Data from the Rio de Janeiro Surveillance Information System. PLoS Negl. Trop. Dis. 2013, 7, e2217. [Google Scholar] [CrossRef] [PubMed]
- Rathore, S.S.; Oberoi, S.; Hilliard, J.; Raja, R.; Ahmed, N.K.; Vishwakarma, Y.; Iqbal, K.; Kumari, C.; Velasquez-Botero, F.; Nieto-Salazar, M.A.; et al. Maternal and foetal-neonatal outcomes of dengue virus infection during pregnancy. Trop. Med. Int. Health 2022, 27, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Basurko, C.; Everhard, S.; Matheus, S.; Restrepo, M.; Hildéral, H.; Lambert, V.; Boukhari, R.; Duvernois, J.-P.; Favre, A.; Valmy, L.; et al. A prospective matched study on symptomatic dengue in pregnancy. PLoS ONE 2018, 13, e0202005. [Google Scholar] [CrossRef] [PubMed]
- Brar, R.; Sikka, P.; Suri, V.; Singh, M.P.; Suri, V.; Mohindra, R.; Biswal, M. Maternal and fetal outcomes of dengue fever in pregnancy: A large prospective and descriptive observational study. Arch. Gynecol. Obstet. 2021, 304, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Paixão, E.S.; Teixeira, M.G.; Costa, M.d.C.N.; Rodrigues, L.C. Dengue during pregnancy and adverse fetal outcomes: A systematic review and meta-analysis. Lancet Infect. Dis. 2016, 16, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Nunes, P.; Nogueira, R.; Coelho, J.; Rodrigues, F.; Salomão, N.; José, C.; de Carvalho, J.; Rabelo, K.; de Azeredo, E.; Basílio-de-Oliveira, R.; et al. A Stillborn Multiple Organs’ Investigation from a Maternal DENV-4 Infection: Histopathological and Inflammatory Mediators Characterization. Viruses 2019, 11, 319. [Google Scholar] [CrossRef]
- Ribeiro, C.F.; Lopes, V.G.S.; Brasil, P.; Pires, A.R.C.; Rohloff, R.; Nogueira, R.M.R. Dengue infection in pregnancy and its impact on the placenta. Int. J. Infect. Dis. 2017, 55, 109–112. [Google Scholar] [CrossRef]
- Paixão, E.S.; Costa, M.d.C.N.; Teixeira, M.G.; Harron, K.; de Almeida, M.F.; Barreto, M.L.; Rodrigues, L.C. Symptomatic dengue infection during pregnancy and the risk of stillbirth in Brazil, 2006–2012: A matched case-control study. Lancet Infect. Dis. 2017, 17, 957–964. [Google Scholar] [CrossRef]
- Sharma, J.B.; Gulati, N. Potential relationship between dengue fever and neural tube defects in a Northern District of India. Int. J. Gynecol. Obstet. 1992, 39, 291–295. [Google Scholar] [CrossRef]
- Ribeiro, C.F.; Lopes, V.G.S.; Brasil, P.; Coelho, J.; Muniz, A.G.; Nogueira, R.M.R. Perinatal Transmission of Dengue: A Report of 7 Cases. J. Pediatr. 2013, 163, 1514–1516. [Google Scholar] [CrossRef]
- Tan, P.C.; Rajasingam, G.; Devi, S.; Omar, S.Z. Dengue infection in pregnancy: Prevalence, vertical transmission, and pregnancy outcome. Obstet. Gynecol. 2008, 111, 1111–1117. [Google Scholar] [CrossRef]
- Arragain, L.; Dupont-Rouzeyrol, M.; O’Connor, O.; Sigur, N.; Grangeon, J.-P.; Huguon, E.; Dechanet, C.; Cazorla, C.; Gourinat, A.-C.; Descloux, E. Vertical Transmission of Dengue Virus in the Peripartum Period and Viral Kinetics in Newborns and Breast Milk: New Data. J. Pediatr. Infect. Dis. Soc. 2016, 6, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Chaturvedi, U.C. Dengue in infants: An overview. FEMS Immunol. Med. Microbiol. 2010, 59, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Nimmanitya, S.; Kliks, S.C.; Burke, D.S.; Nisalak, A. Evidence That Maternal Dengue Antibodies Are Important in the Development of Dengue Hemorrhagic Fever in Infants. Am. J. Trop. Med. Hyg. 1988, 38, 411–419. [Google Scholar] [CrossRef]
- Simmons, C.P.; Chau, T.N.B.; Thuy, T.T.; Tuan, N.M.; Hoang, D.M.; Thien, N.T.; Lien, L.B.; Quy, N.T.; Hieu, N.T.; Hien, T.T.; et al. Maternal Antibody and Viral Factors in the Pathogenesis of Dengue Virus in Infants. J. Infect. Dis. 2007, 196, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Tsarev, S.A.; Sanders, M.L.; Vaughn, D.W.; Innis, B.L. Phylogenetic analysis suggests only one serotype of Japanese encephalitis virus. Vaccine 2000, 18, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Auerswald, H.; Maquart, P.-O.; Chevalier, V.; Boyer, S. Mosquito Vector Competence for Japanese Encephalitis Virus. Viruses 2021, 13, 1154. [Google Scholar] [CrossRef] [PubMed]
- Solomon, T.; Ni, H.; Beasley, D.W.C.; Ekkelenkamp, M.; Cardosa, M.J.; Barrett, A.D.T. Origin and Evolution of Japanese Encephalitis Virus in Southeast Asia. J. Virol. 2003, 77, 3091–3098. [Google Scholar] [CrossRef]
- van den Hurk, A.F.; Skinner, E.; Ritchie, S.A.; Mackenzie, J.S. The Emergence of Japanese Encephalitis Virus in Australia in 2022: Existing Knowledge of Mosquito Vectors. Viruses 2022, 14, 1208. [Google Scholar] [CrossRef]
- Kallas, E.G.; Precioso, A.R.; Palacios, R.; Thomé, B.; Braga, P.E.; Vanni, T.; Campos, L.M.A.; Ferrari, L.; Mondini, G.; da Graça Salomão, M.; et al. Safety and immunogenicity of the tetravalent, live-attenuated dengue vaccine Butantan-DV in adults in Brazil: A two-step, double-blind, randomised placebo-controlled phase 2 trial. Lancet Infect. Dis. 2020, 20, 839–850. [Google Scholar] [CrossRef]
- Rivera, L.; Biswal, S.; Sáez-Llorens, X.; Reynales, H.; López-Medina, E.; Borja-Tabora, C.; Bravo, L.; Sirivichayakul, C.; Kosalaraksa, P.; Martinez Vargas, L.; et al. Three-year Efficacy and Safety of Takeda’s Dengue Vaccine Candidate (TAK-003). Clin. Infect. Dis. 2022, 75, 107–117. [Google Scholar] [CrossRef]
- Whitehead, S.S. Development of TV003/TV005, a single dose, highly immunogenic live attenuated dengue vaccine; what makes this vaccine different from the Sanofi-Pasteur CYD™ vaccine? Expert Rev. Vaccines 2016, 15, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Gould, E.A.; Solomon, T. Pathogenic flaviviruses. Lancet 2008, 371, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Skipetrova, A.; Wartel, T.A.; Gailhardou, S. Dengue vaccination during pregnancy–An overview of clinical trials data. Vaccine 2018, 36, 3345–3350. [Google Scholar] [CrossRef]
- Campbell, G.; Hills, S.; Fischer, M.; Jacobson, J.; Hoke, C.; Hombach, J.; Marfin, A.; Solomon, T.; Tsai, T.; Tsui, V.; et al. Estimated global incidence of Japanese encephalitis. Bull. World Health Organ. 2011, 89, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, J.S.; Williams, D.T.; van den Hurk, A.F.; Smith, D.W.; Currie, B.J. Japanese Encephalitis Virus: The Emergence of Genotype IV in Australia and Its Potential Endemicity. Viruses 2022, 14, 2480. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Sharma, P.; Shukla, K.K.; Misra, S.; Nyati, K.K. Japanese encephalitis virus: Associated immune response and recent progress in vaccine development. Microb. Pathog. 2019, 136, 103678. [Google Scholar] [CrossRef] [PubMed]
- Le Flohic, G.; Porphyre, V.; Barbazan, P.; Gonzalez, J.-P. Review of Climate, Landscape, and Viral Genetics as Drivers of the Japanese Encephalitis Virus Ecology. PLoS Negl. Trop. Dis. 2013, 7, e2208. [Google Scholar] [CrossRef] [PubMed]
- Burns, K.F. Congenital Japanese B Encephalitis Infection of Swine. Exp. Biol. Med. 1950, 75, 621–625. [Google Scholar] [CrossRef]
- Turtle, L.; Solomon, T. Japanese encephalitis—The prospects for new treatments. Nat. Rev. Neurol. 2018, 14, 298–313. [Google Scholar] [CrossRef]
- Sharma, K.B.; Vrati, S.; Kalia, M. Pathobiology of Japanese encephalitis virus infection. Mol. Asp. Med. 2021, 81, 100994. [Google Scholar] [CrossRef]
- Solomon, T. Flavivirus Encephalitis. N. Engl. J. Med. 2004, 351, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Solomon, T.; Winter, P.M. Neurovirulence and host factors in flavivirus encephalitis—Evidence from clinical epidemiology. In Emergence and Control of Zoonotic Viral Encephalitides; Springer: Vienna, Austria, 2004; pp. 161–170. [Google Scholar]
- Mathur, A.; Arora, K.L.; Chaturvedi, U.C. Congenital infection of mice with Japanese encephalitis virus. Infect. Immun. 1981, 34, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Mathur, A.; Arora, K.L.; Chaturvedi, U.C. Transplacental Japanese Encephalitis Virus (JEV) Infection in Mice During Consecutive Pregnancies. J. Gen. Virol. 1982, 59, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Turtle, L.; Easton, A.; Defres, S.; Ellul, M.; Bovill, B.; Hoyle, J.; Jung, A.; Lewthwaite, P.; Solomon, T. ‘More than devastating’—Patient experiences and neurological sequelae of Japanese encephalitis. J. Travel Med. 2019, 26, taz064. [Google Scholar] [CrossRef]
- Verma, R.; Junewar, V.; Praharaj, H.N. Unusual association of eclamptic encephalopathy and Japanese encephalitis. Case Rep. 2012, 2012, bcr0220125840. [Google Scholar] [CrossRef]
- Chaturvedi, U.C.; Mathur, A.; Chandra, A.; Das, S.K.; Tandon, H.O.; Singh, U.K. Transplacental Infection with Japanese Encephalitis Virus. J. Infect. Dis. 1980, 141, 712–715. [Google Scholar] [CrossRef]
- Mathur, A.; Tandon, H.O.; Mathur, K.R.; Sarkari, N.B.; Singh, U.K.; Chaturvedi, U.C. Japanese encephalitis virus infection during pregnancy. Indian J. Med. Res. 1985, 81, 9–12. [Google Scholar]
- Zhu, A.; Petrakis, N.; Gaber, M.; Mason, D.; Clifford, V.; Kelly, J. A case of Japanese encephalitis in a Victorian infant. Med. J. Aust. 2022, 217, 79–80. [Google Scholar] [CrossRef]
- Chancey, C.; Grinev, A.; Volkova, E.; Rios, M. The global ecology and epidemiology of West Nile virus. BioMed Res. Int. 2015, 2015, 376230. [Google Scholar] [CrossRef]
- Petersen, L.R. Epidemiology of West Nile Virus in the United States: Implications for Arbovirology and Public Health. J. Med. Entomol. 2019, 56, 1456–1462. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). West Nile Virus: Final Cumulative Maps and Data for 1999–2020; Centers for Disease Control and Prevention (CDC): Washington, WA, USA, 17 December 2021. Available online: https://www.cdc.gov/westnile/statsmaps/cumMapsData.html (accessed on 4 February 2022).
- Gray, T.J.; Burrow, J.N.; Markey, P.G.; Whelan, P.I.; Jackson, J.; Smith, D.W.; Currie, B.J. West nile virus (Kunjin subtype) disease in the northern territory of Australia--a case of encephalitis and review of all reported cases. Am. J. Trop. Med. Hyg. 2011, 85, 952–956. [Google Scholar] [CrossRef] [PubMed]
- Komar, N.; Langevin, S.; Hinten, S.; Nemeth, N.; Edwards, E.; Hettler, D.; Davis, B.; Bowen, R.; Bunning, M. Experimental infection of North American birds with the New York 1999 strain of West Nile virus. Emerg. Infect. Dis. 2003, 9, 311–322. [Google Scholar] [CrossRef]
- Ludlow, M.; Kortekaas, J.; Herden, C.; Hoffmann, B.; Tappe, D.; Trebst, C.; Griffin, D.E.; Brindle, H.E.; Solomon, T.; Brown, A.S.; et al. Neurotropic virus infections as the cause of immediate and delayed neuropathology. Acta Neuropathol. 2016, 131, 159–184. [Google Scholar] [CrossRef]
- Sejvar, J.J. The long-term outcomes of human West Nile virus infection. Clin. Infect. Dis. 2007, 44, 1617–1624. [Google Scholar] [CrossRef]
- Bruno, J.; Rabito, F.J., Jr.; Dildy, G.A., 3rd. West nile virus meningoencephalitis during pregnancy. J. La. State Med. Soc. 2004, 156, 204–205. [Google Scholar] [PubMed]
- Stewart, R.D.; Bryant, S.N.; Sheffield, J.S. West nile virus infection in pregnancy. Case Rep. Infect. Dis. 2013, 2013, 351872. [Google Scholar] [CrossRef]
- Jamieson, D.J.; Jernigan, D.B.; Ellis, J.E.; Treadwell, T.A. Emerging infections and pregnancy: West Nile virus, monkeypox, severe acute respiratory syndrome, and bioterrorism. Clin. Perinatol. 2005, 32, 765–776. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, D.R.; Kuhn, S.; Kniss, K.L.; Hinckley, A.F.; Rasmussen, S.A.; Pape, W.J.; Kightlinger, L.K.; Beecham, B.D.; Miller, T.K.; Neitzel, D.F.; et al. Birth outcomes following West Nile Virus infection of pregnant women in the United States: 2003–2004. Pediatrics 2006, 117, e537–e545. [Google Scholar] [CrossRef]
- Alpert, S.G.; Fergerson, J.; Noël, L.P. Intrauterine West Nile virus: Ocular and systemic findings. Am. J. Ophthalmol. 2003, 136, 733–735. [Google Scholar] [CrossRef]
- Lindsey, N.P.; Hayes, E.B.; Staples, J.E.; Fischer, M. West Nile virus disease in children, United States, 1999–2007. Pediatrics 2009, 123, e1084–e1089. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Possible West Nile virus transmission to an infant through breast-feeding--Michigan, 2002. Morb. Mortal. Wkly. Rep. 2002, 51, 877–878. [Google Scholar]
- de Oliveira Figueiredo, P.; Stoffella-Dutra, A.G.; Barbosa Costa, G.; Silva de Oliveira, J.; Dourado Amaral, C.; Duarte Santos, J.; Soares Rocha, K.L.; Araújo Júnior, J.P.; Lacerda Nogueira, M.; Zazá Borges, M.A.; et al. Re-Emergence of Yellow Fever in Brazil during 2016–2019: Challenges, Lessons Learned, and Perspectives. Viruses 2020, 12, 1233. [Google Scholar] [CrossRef] [PubMed]
- Douam, F.; Ploss, A. Yellow Fever Virus: Knowledge Gaps Impeding the Fight Against an Old Foe. Trends Microbiol. 2018, 26, 913–928. [Google Scholar] [CrossRef] [PubMed]
- Monath, T.P.; Vasconcelos, P.F. Yellow fever. J. Clin. Virol. 2015, 64, 160–173. [Google Scholar] [CrossRef]
- World Health Organization. Vaccines and vaccination against Yellow fever. WHO position paper–June 2013. Wkly. Epidemiol. Rec. 2013, 88, 269–283. [Google Scholar]
- Gardner, C.L.; Ryman, K.D. Yellow fever: A reemerging threat. Clin. Lab. Med. 2010, 30, 237–260. [Google Scholar] [CrossRef]
- Staples, J.E.; Monath, T.P. Yellow fever: 100 years of discovery. JAMA 2008, 300, 960–962. [Google Scholar] [CrossRef]
- Quaresma, J.A.; Pagliari, C.; Medeiros, D.B.; Duarte, M.I.; Vasconcelos, P.F. Immunity and immune response, pathology and pathologic changes: Progress and challenges in the immunopathology of Yellow fever. Rev. Med. Virol. 2013, 23, 305–318. [Google Scholar] [CrossRef]
- Lindenbach, B.D.; Murray, C.L.; Thiel, H.J.; Rice, C.M. Flaviviridae. In Fields Virology; Knipe, D.M., Howley, P.M., Eds.; Lippincott-Raven: Philadelphia, PA, USA, 2013; pp. 747–794. [Google Scholar]
- Rudolph, K.E.; Lessler, J.; Moloney, R.M.; Kmush, B.; Cummings, D.A. Incubation periods of mosquito-borne viral infections: A systematic review. Am. J. Trop. Med. Hyg. 2014, 90, 882–891. [Google Scholar] [CrossRef]
- Domingo, C.; Charrel, R.N.; Schmidt-Chanasit, J.; Zeller, H.; Reusken, C. Yellow fever in the diagnostics laboratory. Emerg. Microbes Infect. 2018, 7, 1–15. [Google Scholar] [CrossRef]
- Tuboi, S.H.; Costa, Z.G.; da Costa Vasconcelos, P.F.; Hatch, D. Clinical and epidemiological characteristics of Yellow fever in Brazil: Analysis of reported cases 1998–2002. Trans. R. Soc. Trop. Med. Hyg. 2007, 101, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Krubiner, C.B.; Schwartz, D.A. Viral Hemorrhagic Fevers in Pregnant Women and the Vaccine Landscape: Comparisons between Yellow Fever, Ebola, and Lassa Fever. Curr. Trop. Med. Rep. 2019, 6, 186–196. [Google Scholar] [CrossRef]
- Bentlin, M.R.; de Barros Almeida, R.A.; Coelho, K.I.; Ribeiro, A.F.; Siciliano, M.M.; Suzuki, A.; Fortaleza, C.M. Perinatal transmission of Yellow fever, Brazil, 2009. Emerg. Infect. Dis. 2011, 17, 1779–1780. [Google Scholar] [CrossRef] [PubMed]
- Diniz, L.M.O.; Romanelli, R.M.C.; de Carvalho, A.L.; Teixeira, D.C.; de Carvalho, L.F.A.; Ferreira Cury, V.; Filho, M.P.L.; Perígolo, G.; Heringer, T.P. Perinatal Yellow Fever: A Case Report. Pediatr. Infect. Dis. J. 2019, 38, 300–301. [Google Scholar] [CrossRef] [PubMed]
- Hunsperger, E.A.; Muñoz-Jordán, J.; Beltran, M.; Colón, C.; Carrión, J.; Vazquez, J.; Acosta, L.N.; Medina-Izquierdo, J.F.; Horiuchi, K.; Biggerstaff, B.J.; et al. Performance of Dengue Diagnostic Tests in a Single-Specimen Diagnostic Algorithm. J. Infect. Dis. 2016, 214, 836–844. [Google Scholar] [CrossRef]
- Communicable Diseases Network Australia (CDNA). Australian National Notifiable Diseases Case Definition: West Nile/Kunjin Virus Infection; Communicable Diseases Network Australia (CDNA): Canberra, Australia, 2010. Available online: https://www.health.gov.au/sites/default/files/documents/2022/06/west-nile-kunjin-virus-infection-surveillance-case-definition.pdf (accessed on 4 February 2022).
- Gibney, K.B.; Edupuganti, S.; Panella, A.J.; Kosoy, O.I.; Delorey, M.J.; Lanciotti, R.S.; Mulligan, M.J.; Fischer, M.; Staples, J.E. Detection of anti-Yellow fever virus immunoglobulin M antibodies at 3-4 years following Yellow fever vaccination. Am. J. Trop. Med. Hyg. 2012, 87, 1112–1115. [Google Scholar] [CrossRef]
- Pham, D.; Howard-Jones, A.R.; Hueston, L.; Jeoffreys, N.; Doggett, S.; Rockett, R.J.; Eden, J.-S.; Sintchenko, V.; Chen, S.C.; O’Sullivan, M.V.; et al. Emergence of Japanese encephalitis in Australia: A diagnostic perspective. Pathology 2022, 54, 669–677. [Google Scholar] [CrossRef]
- Houghton-Triviño, N.; Montaña, D.; Castellanos, J. Dengue-Yellow fever sera cross-reactivity; challenges for diagnosis. Rev. Salud Publica 2008, 10, 299–307. [Google Scholar] [CrossRef]
- Changal, K.H.; Raina, A.H.; Raina, A.; Raina, M.; Bashir, R.; Latief, M.; Mir, T.; Changal, Q.H. Differentiating secondary from primary dengue using IgG to IgM ratio in early dengue: An observational hospital based clinico-serological study from North India. BMC Infect. Dis. 2016, 16, 715. [Google Scholar] [CrossRef]
- Matheus, S.V.; Deparis, X.; Labeau, B.; Lelarge, J.; Morvan, J.; Dussart, P. Discrimination between Primary and Secondary Dengue Virus Infection by an Immunoglobulin G Avidity Test Using a Single Acute-Phase Serum Sample. J. Clin. Microbiol. 2005, 43, 2793–2797. [Google Scholar] [CrossRef]
- Nguyen, T.H.T.; Clapham, H.E.; Phung, K.L.; Nguyen, T.K.; Dinh, T.T.; Nguyen, T.H.Q.; Tran, V.N.; Whitehead, S.; Simmons, C.; Wolbers, M.; et al. Methods to discriminate primary from secondary dengue during acute symptomatic infection. BMC Infect. Dis. 2018, 18, 375. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). West Nile Virus: Diagnostic Testing; Centers for Disease Control and Prevention (CDC): Atlanta, GA, USA, 2021. Available online: https://www.cdc.gov/westnile/healthcareproviders/healthCareProviders-Diagnostic.html (accessed on 4 February 2022).
- Domingo, C.; Patel, P.; Yillah, J.; Weidmann, M.; Méndez, J.A.; Nakouné, E.R.; Niedrig, M. Advanced Yellow fever virus genome detection in point-of-care facilities and reference laboratories. J. Clin. Microbiol. 2012, 50, 4054–4060. [Google Scholar] [CrossRef] [PubMed]
- Barzon, L.; Percivalle, E.; Pacenti, M.; Rovida, F.; Zavattoni, M.; Del Bravo, P.; Cattelan, A.M.; Palù, G.; Baldanti, F. Virus and Antibody Dynamics in Travelers with Acute Zika Virus Infection. Clin. Infect. Dis. 2018, 66, 1173–1180. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Liao, Y.; Ke, X.; Zhou, J.; Chen, Y.; Gao, L.; Chen, Q.; Yu, S. Comparison of reverse transcription loop-mediated isothermal amplification, conventional PCR and real-time PCR assays for Japanese encephalitis virus. Mol. Biol. Rep. 2011, 38, 4063–4070. [Google Scholar] [CrossRef] [PubMed]
- Maamary, J.; Maddocks, S.; Barnett, Y.; Wong, S.; Rodriguez, M.; Hueston, L.; Jeoffreys, N.; Eden, J.-S.; Dwyer, D.E.; Floyd, T.; et al. First detection of locally acquired Japanese Encephalitis Virus in New South Wales, Australia using clinical metagenomics. Emerg. Infect. Dis. 2023, in press.
- Public Health Laboratory Network. Flavivirus: Laboratory Case Definition; Public Health Laboratory Network: Canberra, Australia, 2022. Available online: https://www.health.gov.au/sites/default/files/documents/2022/07/flavivirus-laboratory-case-definition.pdf (accessed on 4 February 2022).
- Communicable Diseases Network Australia (CDNA). CDNA National Guidelines for Public Health Units: Zika Virus Infection; Communicable Diseases Network Australia (CDNA): Canberra, Australia, 16 December 2016. Available online: https://www.health.gov.au/sites/default/files/documents/2020/02/zika-virus-cdna-national-guidelines-for-public-health-units.pdf (accessed on 4 February 2022).
- Tricou, V.; Minh, N.N.; Farrar, J.; Tran, H.T.; Simmons, C.P. Kinetics of Viremia and NS1 Antigenemia Are Shaped by Immune Status and Virus Serotype in Adults with Dengue. PLoS Negl. Trop. Dis. 2011, 5, e1309. [Google Scholar] [CrossRef]
- Guzman, M.G.; Jaenisch, T.; Gaczkowski, R.; Ty Hang, V.T.; Sekaran, S.D.; Kroeger, A.; Vazquez, S.; Ruiz, D.; Martinez, E.; Mercado, J.C.; et al. Multi-Country Evaluation of the Sensitivity and Specificity of Two Commercially-Available NS1 ELISA Assays for Dengue Diagnosis. PloS Negl. Trop. Dis. 2010, 4, e811. [Google Scholar] [CrossRef]
- Tiroumourougane, S.V. Japanese viral encephalitis. Postgrad. Med. J. 2002, 78, 205–215. [Google Scholar] [CrossRef]
- Chowers, M.Y.; Lang, R.; Nassar, F.; Ben-David, D.; Giladi, M.; Rubinshtein, E.; Itzhaki, A.; Mishal, J.; Siegman-Igra, Y.; Kitzes, R.; et al. Clinical characteristics of the West Nile fever outbreak, Israel, 2000. Emerg. Infect. Dis. 2001, 7, 675–678. [Google Scholar] [CrossRef]
- Popescu, C.P.; Florescu, S.A.; Hasbun, R.; Harxhi, A.; Evendar, R.; Kahraman, H.; Neuberger, A.; Codreanu, D.; Zaharia, M.F.; Tosun, S.; et al. Prediction of unfavorable outcomes in West Nile virus neuroinvasive infection-Result of a multinational ID-IRI study. J. Clin. Virol. 2020, 122, 104213. [Google Scholar] [CrossRef]
- World Health Organization. Dengue: Guidelines for Diagnosis, Treatment, Prevention and Control; World Health Organization: Geneva, Switzerland, 2009; Available online: https://apps.who.int/iris/bitstream/handle/10665/44188/9789241547871_eng.pdf (accessed on 4 February 2022).
- Monath, T.P. Treatment of Yellow fever. Antivir. Res. 2008, 78, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Gnann, J.W., Jr.; Agrawal, A.; Hart, J.; Buitrago, M.; Carson, P.; Hanfelt-Goade, D.; Tyler, K.; Spotkov, J.; Freifeld, A.; Moore, T.; et al. Lack of Efficacy of High-Titered Immunoglobulin in Patients with West Nile Virus Central Nervous System Disease. Emerg. Infect. Dis. 2019, 25, 2064–2073. [Google Scholar] [CrossRef] [PubMed]
- Murray, K.O.; Baraniuk, S.; Resnick, M.; Arafat, R.; Kilborn, C.; Shallenberger, R.; York, T.L.; Martinez, D.; Malkoff, M.; Elgawley, N.; et al. Clinical investigation of hospitalized human cases of West Nile virus infection in Houston, Texas, 2002–2004. Vector Borne Zoonotic Dis. 2008, 8, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Doyle, M.P.; Genualdi, J.R.; Bailey, A.L.; Kose, N.; Gainza, C.; Rodriguez, J.; Reeder, K.M.; Nelson, C.A.; Jethva, P.N.; Sutton, R.E.; et al. Isolation of a Potently Neutralizing and Protective Human Monoclonal Antibody Targeting Yellow Fever Virus. mBio 2022, 13, e00512-22. [Google Scholar] [CrossRef] [PubMed]
- Kai, Y.; Lilan, X.; Yaoming, L. Monoclonal Antibody That Inhibits Cleavage Activity of Japanese Encephalitis Virus NS3. Monoclon. Antibodies Immunodiagn. Immunother. 2021, 40, 28–32. [Google Scholar] [CrossRef]
- Lu, J.; Chen, L.; Du, P.; Guo, J.; Wang, X.; Jiang, Y.; Yu, Y.; Wang, R.; Yang, Z. A human monoclonal antibody to neutralize all four serotypes of dengue virus derived from patients at the convalescent phase of infection. Virology 2022, 576, 74–82. [Google Scholar] [CrossRef]
- Ozawa, T.; Masaki, H.; Takasaki, T.; Aoyama, I.; Yumisashi, T.; Yamanaka, A.; Konishi, E.; Ohnuki, Y.; Muraguchi, A.; Kishi, H. Human monoclonal antibodies against West Nile virus from Japanese encephalitis-vaccinated volunteers. Antivir. Res. 2018, 154, 58–65. [Google Scholar] [CrossRef]
- Sootichote, R.; Puangmanee, W.; Benjathummarak, S.; Kowaboot, S.; Yamanaka, A.; Boonnak, K.; Ampawong, S.; Chatchen, S.; Ramasoota, P.; Pitaksajjakul, P. Potential Protective Effect of Dengue NS1 Human Monoclonal Antibodies against Dengue and Zika Virus Infections. Biomedicines 2023, 11, 227. [Google Scholar] [CrossRef]
- Christie, S.; Chan, V.; Mollayeva, T.; Colantonio, A. Systematic review of rehabilitation intervention outcomes of adult and paediatric patients with infectious encephalitis. BMJ Open 2018, 8, e015928. [Google Scholar] [CrossRef]
- Rajapakse, S.; de Silva, N.L.; Weeratunga, P.; Rodrigo, C.; Fernando, S.D. Prophylactic and therapeutic interventions for bleeding in dengue: A systematic review. Trans. R. Soc. Trop. Med. Hyg. 2017, 111, 433–439. [Google Scholar] [CrossRef]
- Escobar, M.F.; Mora, B.L.; Cedano, J.A.; Loaiza, S.; Rosso, F. Comprehensive treatment in severe dengue during preterm and term labor: Could tocolysis be useful? J. Matern.-Fetal Neonatal Med. 2020, 33, 2445–2450. [Google Scholar] [CrossRef] [PubMed]
- Oladapo, O.T.; Okusanya, B.O.; Abalos, E.; Gallos, I.D.; Papadopoulou, A. Intravenous versus intramuscular prophylactic oxytocin for reducing blood loss in the third stage of labour. Cochrane Database Syst. Rev. 2020, 2020. [Google Scholar] [CrossRef]
- Gabiane, G.; Yen, P.S.; Failloux, A.B. Aedes mosquitoes in the emerging threat of urban Yellow fever transmission. Rev. Med. Virol. 2022, 32, e2333. [Google Scholar] [CrossRef] [PubMed]
- Gujral, I.B.; Zielinski-Gutierrez, E.C.; LeBailly, A.; Nasci, R. Behavioral risks for West Nile virus disease, northern Colorado, 2003. Emerg. Infect. Dis. 2007, 13, 419–425. [Google Scholar] [CrossRef]
- Han, L.L.; Popovici, F.; Alexander, J.P., Jr.; Laurentia, V.; Tengelsen, L.A.; Cernescu, C.; Gary, H.E., Jr.; Ion-Nedelcu, N.; Campbell, G.L.; Tsai, T.F. Risk factors for West Nile virus infection and meningoencephalitis, Romania, 1996. J. Infect. Dis. 1999, 179, 230–233. [Google Scholar] [CrossRef]
- Kabilan, L. Control of Japanese encephalitis in India: A reality. Indian J. Pediatr. 2004, 71, 707–712. [Google Scholar] [CrossRef]
- Monath, T.P. Yellow fever vaccine. Expert Rev. Vaccines 2005, 4, 553–574. [Google Scholar] [CrossRef]
- Petersen, E.E.; Meaney-Delman, D.; Neblett-Fanfair, R.; Havers, F.; Oduyebo, T.; Hills, S.L.; Rabe, I.B.; Lambert, A.; Abercrombie, J.; Martin, S.W.; et al. Update: Interim Guidance for Preconception Counseling and Prevention of Sexual Transmission of Zika Virus for Persons with Possible Zika Virus Exposure-United States, September 2016. Morb. Mortal. Wkly. Rep. 2016, 65, 1077–1081. [Google Scholar] [CrossRef]
- World Health Organization. Prevention of Potential Sexual Transmission of Zika Virus: Interim Guidance; World Health Organization: Geneva, Switzerland, 6 September 2016; Available online: http://apps.who.int/iris/bitstream/10665/204421/1/WHO_ZIKV_MOC_16.1_eng.pdf?ua=1 (accessed on 4 February 2022).
- Heydari, N.; Larsen, D.A.; Neira, M.; Beltrán Ayala, E.; Fernandez, P.; Adrian, J.; Rochford, R.; Stewart-Ibarra, A.M. Household Dengue Prevention Interventions, Expenditures, and Barriers to Aedes aegypti Control in Machala, Ecuador. Int. J. Environ. Res. Public Health 2017, 14, 196. [Google Scholar] [CrossRef]
- Hassan, T.; Bashir, R.; Abdelrahman, D.; Madni, H.; M El Hussein, A.; Elkidir, I.; Enan, K. Transmission of Yellow fever vaccine virus from breast feeding mothers to their infants: Reporting of Yellow fever virus (YFV) RNA detection in milk specimens. F1000Research 2022, 11, 76. [Google Scholar] [CrossRef]
- Kuhn, S.; Twele-Montecinos, L.; MacDonald, J.; Webster, P.; Law, B. Case report: Probable transmission of vaccine strain of Yellow fever virus to an infant via breast milk. Can. Med. Assoc. J. 2011, 183, E243–E245. [Google Scholar] [CrossRef] [PubMed]
- Traiber, C.; Coelho-Amaral, P.; Ritter, V.R.; Winge, A. Infant meningoencephalitis caused by Yellow fever vaccine virus transmitted via breastmilk. J. Pediatr. 2011, 87, 269–272. [Google Scholar] [CrossRef] [PubMed]
- Cavalcanti, D.P.; Salomão, M.A.; Lopez-Camelo, J.; Pessoto, M.A. Early exposure to Yellow fever vaccine during pregnancy. Trop. Med. Int. Health 2007, 12, 833–837. [Google Scholar] [CrossRef] [PubMed]
- D’Acremont, V.; Tremblay, S.; Genton, B. Impact of vaccines given during pregnancy on the offspring of women consulting a travel clinic: A longitudinal study. J. Travel Med. 2008, 15, 77–81. [Google Scholar] [CrossRef]
- Nasidi, A.; Monath, T.P.; Vandenberg, J.; Tomori, O.; Calisher, C.H.; Hurtgen, X.; Munube, G.R.; Sorungbe, A.O.; Okafor, G.C.; Wali, S. Yellow fever vaccination and pregnancy: A four-year prospective study. Trans. R. Soc. Trop. Med. Hyg. 1993, 87, 337–339. [Google Scholar] [CrossRef]
- Robert, E.; Vial, T.; Schaefer, C.; Arnon, J.; Reuvers, M. Exposure to Yellow fever vaccine in early pregnancy. Vaccine 1999, 17, 283–285. [Google Scholar] [CrossRef]
- Staples, J.E.; Gershman, M.; Fischer, M. Yellow fever vaccine: Recommendations of the Advisory Committee on Immunization Practices (ACIP). MMWR Recomm. Rep. 2010, 59, 1–27. [Google Scholar]
- Suzano, C.E.; Amaral, E.; Sato, H.K.; Papaiordanou, P.M. The effects of Yellow fever immunization (17DD) inadvertently used in early pregnancy during a mass campaign in Brazil. Vaccine 2006, 24, 1421–1426. [Google Scholar] [CrossRef]
- Heffelfinger, J.D.; Li, X.; Batmunkh, N.; Grabovac, V.; Diorditsa, S.; Liyanage, J.B.; Pattamadilok, S.; Bahl, S.; Vannice, K.S.; Hyde, T.B.; et al. Japanese Encephalitis Surveillance and Immunization—Asia and Western Pacific Regions, 2016. Morb. Mortal. Wkly. Rep. 2017, 66, 579–583. [Google Scholar] [CrossRef]
- Li, X.; Ma, S.-J.; Liu, X.; Jiang, L.-N.; Zhou, J.-H.; Xiong, Y.-Q.; Ding, H.; Chen, Q. Immunogenicity and safety of currently available Japanese encephalitis vaccines: A systematic review. Hum. Vaccines Immunother. 2014, 10, 3579–3593. [Google Scholar] [CrossRef]
- Schiøler, K.L.; Samuel, M.; Wai, K.L. Vaccines for preventing Japanese encephalitis. Cochrane Database Syst. Rev. 2007. [Google Scholar] [CrossRef] [PubMed]
- Eckels, K.; Yongxin, Y.; Dubois, D.; Marchette, N.; Trent, D.; Johnson, A. Japanese encephalitis virus live-attenuated vaccine, Chinese strain SA14-14-2; adaptation to primary canine kidney cell cultures and preparation of a vaccine for human use. Vaccine 1988, 6, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Appaiahgari, M.B.; Vrati, S. IMOJEV®: A Yellow fever virus-based novel Japanese encephalitis vaccine. Expert Rev. Vaccines 2010, 9, 1371–1384. [Google Scholar] [CrossRef] [PubMed]
- Appaiahgari, M.B.; Vrati, S. Clinical development of IMOJEV®—A recombinant Japanese encephalitis chimeric vaccine (JE-CV). Expert Opin. Biol. Ther. 2012, 12, 1251–1263. [Google Scholar] [CrossRef]
- Erra, E.O.; Kantele, A. The Vero cell-derived, inactivated, SA 14-14-2 strain-based vaccine (Ixiaro) for prevention of Japanese encephalitis. Expert Rev. Vaccines 2015, 14, 1167–1179. [Google Scholar] [CrossRef] [PubMed]
- Jelinek, T. IXIARO® updated: Overview of clinical trials and developments with the inactivated vaccine against Japanese encephalitis. Expert Rev. Vaccines 2013, 12, 859–869. [Google Scholar] [CrossRef]
- Khodr, Z.G.; Hall, C.; Chang, R.N.; Bukowinski, A.T.; Gumbs, G.R.; Conlin, A.M.S. Japanese encephalitis vaccination in pregnancy among U.S. active duty military women. Vaccine 2020, 38, 4529–4535. [Google Scholar] [CrossRef]
- Essink, B.; Chu, L.; Seger, W.; Barranco, E.; Le Cam, N.; Bennett, H.; Faughnan, V.; Pajon, R.; Paila, Y.D.; Bollman, B.; et al. The safety and immunogenicity of two Zika virus mRNA vaccine candidates in healthy flavivirus baseline seropositive and seronegative adults: The results of two randomised, placebo-controlled, dose-ranging, phase 1 clinical trials. Lancet Infect. Dis. 2023. [Google Scholar] [CrossRef]
- Yeasmin, M.; Molla, M.M.A.; Masud, H.; Saif-Ur-Rahman, K.M. Safety and immunogenicity of Zika virus vaccine: A systematic review of clinical trials. Rev. Med. Virol. 2022, 33, e2385. [Google Scholar] [CrossRef]
- Kaiser, J.A.; Barrett, A.D.T. Twenty Years of Progress Toward West Nile Virus Vaccine Development. Viruses 2019, 11, 823. [Google Scholar] [CrossRef]
- McArthur, M.A.; Sztein, M.B.; Edelman, R. Dengue vaccines: Recent developments, ongoing challenges and current candidates. Expert Rev. Vaccines 2013, 12, 933–953. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.J.; Yoon, I.-K. A review of Dengvaxia®: Development to deployment. Hum. Vaccines Immunother. 2019, 15, 2295–2314. [Google Scholar] [CrossRef]
- Sridhar, S.; Luedtke, A.; Langevin, E.; Zhu, M.; Bonaparte, M.; Machabert, T.; Savarino, S.; Zambrano, B.; Moureau, A.; Khromava, A.; et al. Effect of Dengue Serostatus on Dengue Vaccine Safety and Efficacy. N. Engl. J. Med. 2018, 379, 327–340. [Google Scholar] [CrossRef] [PubMed]
- Larson, H.J.; Hartigan-Go, K.; de Figueiredo, A. Vaccine confidence plummets in the Philippines following dengue vaccine scare: Why it matters to pandemic preparedness. Hum. Vaccines Immunother. 2019, 15, 625–627. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.H.; Kinney, R.M.; Livengood, J.A.; Bolling, B.; Arguello, J.J.; Luy, B.E.; Silengo, S.J.; Boroughs, K.L.; Stovall, J.L.; Kalanidhi, A.P.; et al. Genetic and Phenotypic Characterization of Manufacturing Seeds for a Tetravalent Dengue Vaccine (DENVax). PLoS Negl. Trop. Dis. 2013, 7, e2243. [Google Scholar] [CrossRef]
- Higa, Y. Dengue Vectors and their Spatial Distribution. Trop. Med. Health 2011, 39, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Abreu, F.V.S.; Ribeiro, I.P.; Ferreira-de-Brito, A.; Santos, A.; Miranda, R.M.; Bonelly, I.S.; Neves, M.; Bersot, M.I.; Santos, T.P.D.; Gomes, M.Q.; et al. Haemagogus leucocelaenus and Haemagogus janthinomys are the primary vectors in the major Yellow fever outbreak in Brazil, 2016–2018. Emerg. Microbes Infect. 2019, 8, 218–231. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.R.; Webb, C.E.; Higgs, S.; van den Hurk, A.F. Japanese Encephalitis Virus Emergence in Australia: Public Health Importance and Implications for Future Surveillance. Vector Borne Zoonotic Dis. 2022, 22, 529–534. [Google Scholar] [CrossRef]
- Williams, P.C.; Bartlett, A.W.; Howard-Jones, A.; McMullan, B.; Khatami, A.; Britton, P.N.; Marais, B.J. Impact of climate change and biodiversity collapse on the global emergence and spread of infectious diseases. J. Paediatr. Child Health 2021, 57, 1811–1818. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Virus | Zika Virus (ZIKV) | Dengue (DENV) | Japanese Encephalitis Virus (JEV) | West Nile Virus (WNV) | Yellow Fever Virus (YFV) |
|---|---|---|---|---|---|
| Incubation time | 2–14 days | 5–7 days | 5–15 days | 2–14 days | 3–6 days |
| Maternal clinical features | Asymptomatic (majority) Fever, pruritic rash (palms and soles involved), arthralgia, headache GBS or (rarely) meningoencephalitis | Significant increase in severe disease and maternal mortality in pregnancy | Asymptomatic or mild disease (majority) Meningoencephalitis in <1% | Asymptomatic (majority) Fever, headache, malaise, nausea Meningitis, encephalitis, acute flaccid paralysis (rare) | Asymptomatic (majority) Nonspecific febrile illness Severe (15% patients)—jaundice, multi-organ failure, coagulopathy |
| Features of congenital and neonatal infection | Severe congenital anomalies (26–65%) including ventriculomegaly, microcephaly, intracranial calcifications, ophthalmological anomalies, and neuro-developmental abnormalities Fetal loss (7–14%) | Significant association with prematurity, low birth weight, miscarriage or foetal deathCongenital malformations and neonatal transmission described, but uncommon | Miscarriage described up to 22 weeks of pregnancy; minimal data on neonatal disease | Rare congenital abnormalities (~4%): lissencephaly, meningoencephalitis, cerebral atrophy, chorioretinitis, coarctation of the aorta | Minimal data |
| Primary vector(s) | Mosquito (Aedes aegypti) | Mosquitos, primarily Aedes spp. | Mosquitos, primarily Culex spp. | Wide range of mosquitos including Culex spp. and Aedes spp. Ticks implicated in vitro but no in vivo transmission determined | Mosquito (Aedes spp., Haemogogus spp.) |
| Pathophysiology (unique features) | Tropism for nervous tissue (especially neural progenitor cells) and placental cells | Severe infection with dengue hemorrhagic fever or dengue shock syndrome, more common in secondary infections | Perivascular transport across the blood-brain barrier into CNS; transplacental infection well described in animal models | Neuronal apoptosis with parenchymal inflammation predominantly affecting subcortical structures; glial cell damage | Apoptosis of mid-zone hepatocytes |
| Diagnostic tools | Antibody detection NAAT Viral culture | Antibody detection NS1 antigen NAAT Viral culture | Antibody detection NAAT, LAMP Viral culture | Antibody detection NAAT Viral culture | Antibody detection NAAT Viral culture |
| Treatment options | Supportive care | Supportive care Avoid non-steroidal anti-inflammatory medications Blood products and vitamin K may be required | Supportive care | Supportive care | Supportive care |
| Vaccination/prevention | Vector control/reducing mosquito exposure Avoid unprotected sex following exposure or acute infection | Vector control/reducing mosquito exposure Vaccine available, however significant concern in DENV-naïve patients due to the risk of precipitating severe secondary infection; live vaccine contraindicated in pregnancy | Vector control/reducing mosquito exposure Multiple vaccines available; IXIARO/JESPECT can be used in pregnancy | Vector control/reducing mosquito exposure | Vector control/reducing mosquito exposure Live attenuated vaccine contraindicated in pregnancy (unless high-risk/severe epidemic) and contraindicated in breastfeeding and infants <6 months |
| Virus | Assay | Sample Types | Advantages | Disadvantages | Comments |
|---|---|---|---|---|---|
| Zika virus (ZKV) | ZKV IgM ZKV IgG | Serum | IgM detectable within the first few days of illness onset, persists for several weeks to months | Cross-reaction with other flaviviruses requires careful interpretation. | Acute and convalescent titers (at 2–4 weeks from illness onset) should be compared. |
| NAAT | Urine, blood, placental tissue, fetal tissue, amniotic fluid, semen, genital tract secretions, saliva | Highly sensitive | Requires fresh tissue (not formalin-fixed). | Not currently used as a first-line assay. Reserved for acutely unwell patients, for confirmatory testing, or in the diagnosis of congenital Zika syndrome (CZS). | |
| Dengue virus (DENV) | NS1 antigen | Serum | Highly sensitive in the early phase of illness (primary and secondary infections). Highly specific for DENV. | Does not differentiate between dengue serotypes. | NS1 antigenemia is shorter in secondary than in primary infection |
| DENV IgM DENV IgG | Serum | IgM detectable within the first few days of primary infection, and persists for 2–3 months. Rapid rise in DENV IgG in secondary infection | Cross-reaction with other flaviviruses requires careful interpretation. Vaccine-induced IgM can persist for years. | Acute and convalescent titers (at 2–4 weeks from illness onset) should be compared. IgM/IgG ratio may be used to differentiate primary (high ratio) from secondary (low ratio) infections. | |
| DENV IgG avidity | Serum | Can help differentiate primary from secondary disease | Only available in specialist centres. | High avidity suggests secondary infection. | |
| NAAT | Blood, urine | Highly specific | Low sensitivity outside the first week of illness. Expensive, not routinely available. | Not currently used as a first-line assay. Reserved for acutely unwell patients or confirmatory testing. | |
| Japanese encephalitis virus (JEV) | JEV IgM JEV IgG | Serum, CSF (IgM) Serum (IgG) | IgM detectable within the first few days of illness onset | Cross-reaction with other flaviviruses requires careful interpretation. Vaccine-induced IgM can persist for years. | Acute and convalescent titers (at 2–4 weeks from illness onset) should be compared. |
| NAAT | CSF, blood, urine, brain tissue | Highly specific for JEV | Low sensitivity outside first few days of illness. Invasive sampling for cerebral tissue. | Early tissue sampling enhances diagnostic yield. | |
| West Nile virus (WNV) | WNV IgM WNV IgG | Serum, CSF (IgM) Serum (IgG) | IgM detectable within the first few days of illness onset | Cross-reaction with other flaviviruses requires careful interpretation. | Acute and convalescent titers (at 2–4 weeks from illness onset) should be compared. |
| NAAT | Blood, urine, CSF | Highly specific | Low sensitivity outside the first week of illness. Expensive, not routinely available. | Not currently used as a first-line assay. Reserved for acutely unwell patients or for confirmatory testing. | |
| Yellow fever virus (YFV) | YFV IgM YFV IgG | Serum | IgM detectable within the first few days of illness onset | Cross-reaction with other flaviviruses requires careful interpretation. Vaccine-induced IgM can persist for years. | Acute and convalescent titers (at 2–4 weeks from illness onset) should be compared. |
| NAAT | Blood, urine | Highly specific | Low sensitivity outside the first week of illness. Expensive, not routinely available. | Not currently used as a first-line assay. Reserved for acutely unwell patients or for confirmatory testing. | |
| All | Pan-flavivirus IgM Pan-flavivirus IgG | Serum | Can be used as a screening assay | Does not differentiate between flaviviruses. | A positive result should be further delineated with specific IgM and IgG for each Flavivirus of interest to enable accurate diagnosis. Acute and convalescent titers (at 2–4 weeks from illness onset) should be compared. |
| Viral culture | CSF, tissue, blood, urine, placental tissue (ZKV), fetal tissues (ZKV) | High specificity; provides viral isolate for further sequencing | Lower sensitivity than NAAT or serology. Requires specialist laboratory with PC3 facilities. | Staff handling viral cultures should be vaccinated when possible. | |
| Histopathology, immunohistochemistry | Tissue | Provides detailed structural information. Immunohistochemistry provides specificity for select viral antigens | Histopathological changes are largely non-specific between flaviviruses. | Requires specialist histopathologist expertise. | |
| Next-generation metagenomic sequencing | Brain tissue, CSF | High specificity Pathogen-agnostic testing | High level of technical expertise is required. High cost. Available in specialist centres only. | Not appropriate for testing samples from non-sterile sites (for example respiratory secretions) or with anticipated low viral loads (for example, serum or urine). |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Howard-Jones, A.R.; Pham, D.; Sparks, R.; Maddocks, S.; Dwyer, D.E.; Kok, J.; Basile, K. Arthropod-Borne Flaviviruses in Pregnancy. Microorganisms 2023, 11, 433. https://doi.org/10.3390/microorganisms11020433
Howard-Jones AR, Pham D, Sparks R, Maddocks S, Dwyer DE, Kok J, Basile K. Arthropod-Borne Flaviviruses in Pregnancy. Microorganisms. 2023; 11(2):433. https://doi.org/10.3390/microorganisms11020433
Chicago/Turabian StyleHoward-Jones, Annaleise R., David Pham, Rebecca Sparks, Susan Maddocks, Dominic E. Dwyer, Jen Kok, and Kerri Basile. 2023. "Arthropod-Borne Flaviviruses in Pregnancy" Microorganisms 11, no. 2: 433. https://doi.org/10.3390/microorganisms11020433
APA StyleHoward-Jones, A. R., Pham, D., Sparks, R., Maddocks, S., Dwyer, D. E., Kok, J., & Basile, K. (2023). Arthropod-Borne Flaviviruses in Pregnancy. Microorganisms, 11(2), 433. https://doi.org/10.3390/microorganisms11020433