
Field Use of Protective Bacteriophages against Pectinolytic Bacteria of Potato

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteriophages and Bacterial Strains
2.2. Laboratory Tests
2.3. Field Trial
2.4. Sequence Analyses
3. Results
3.1. Potato Cultivar Sensitivity Test
3.2. Virus Sequence Analysis
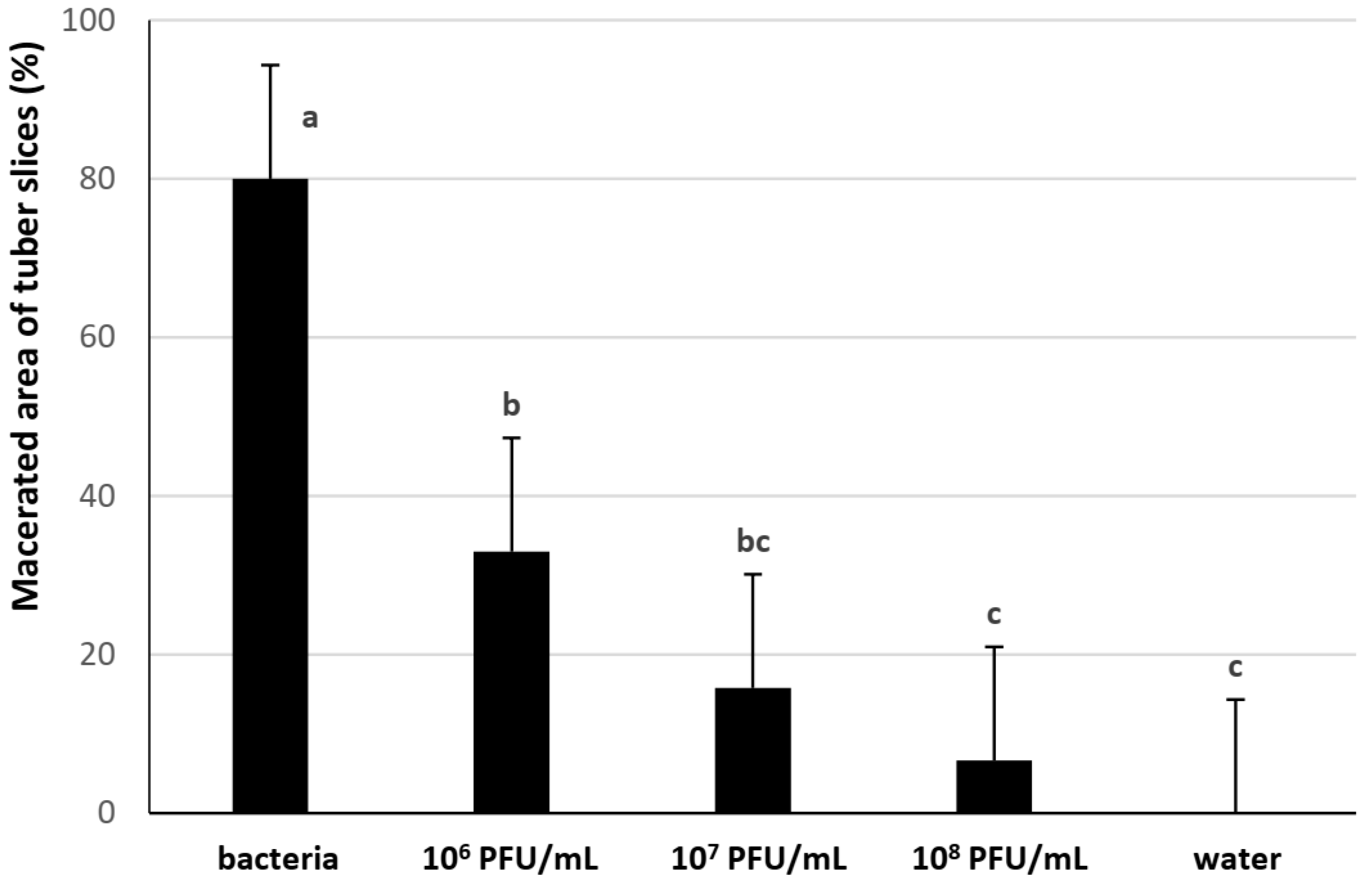
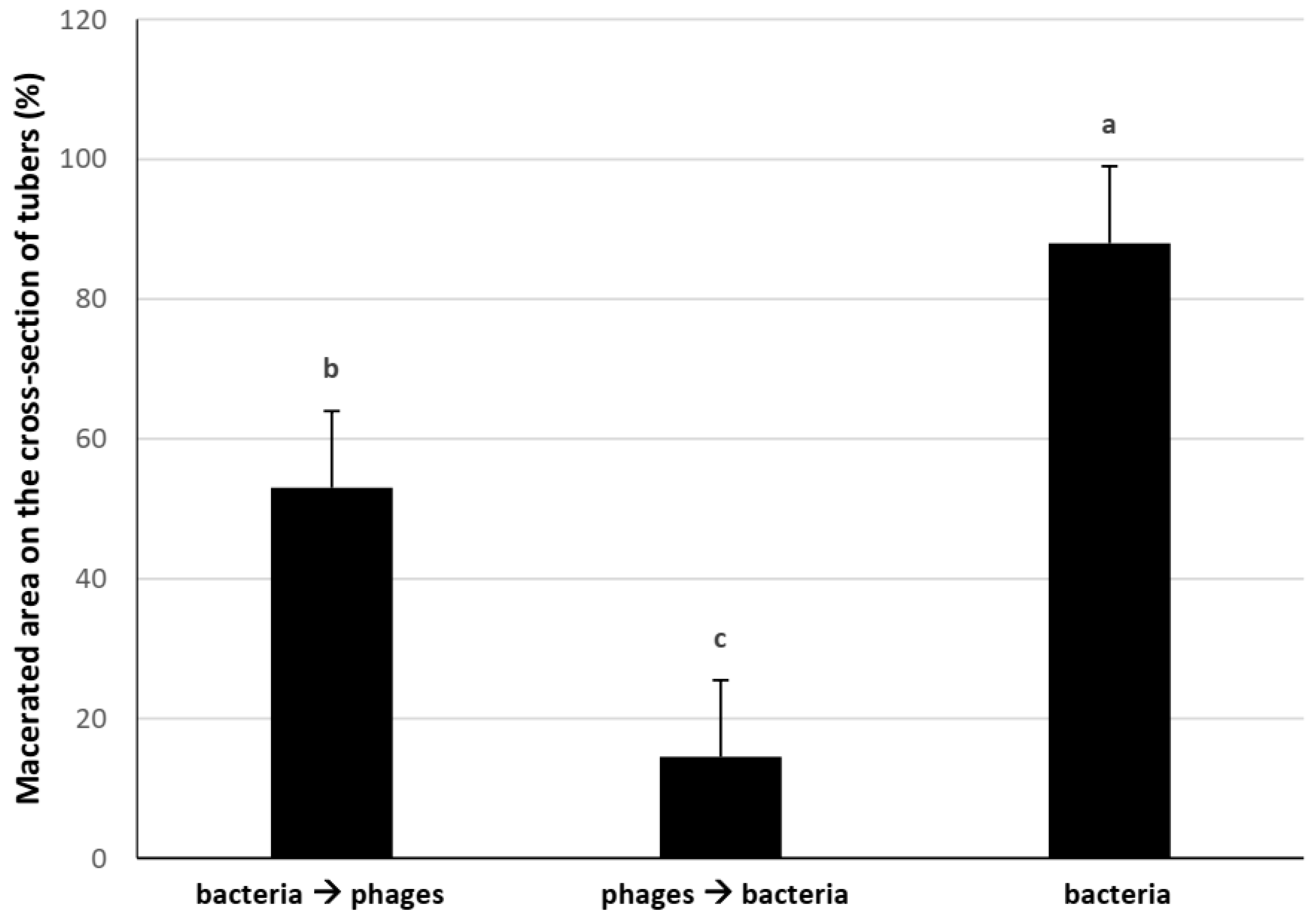
3.3. Laboratory Trial
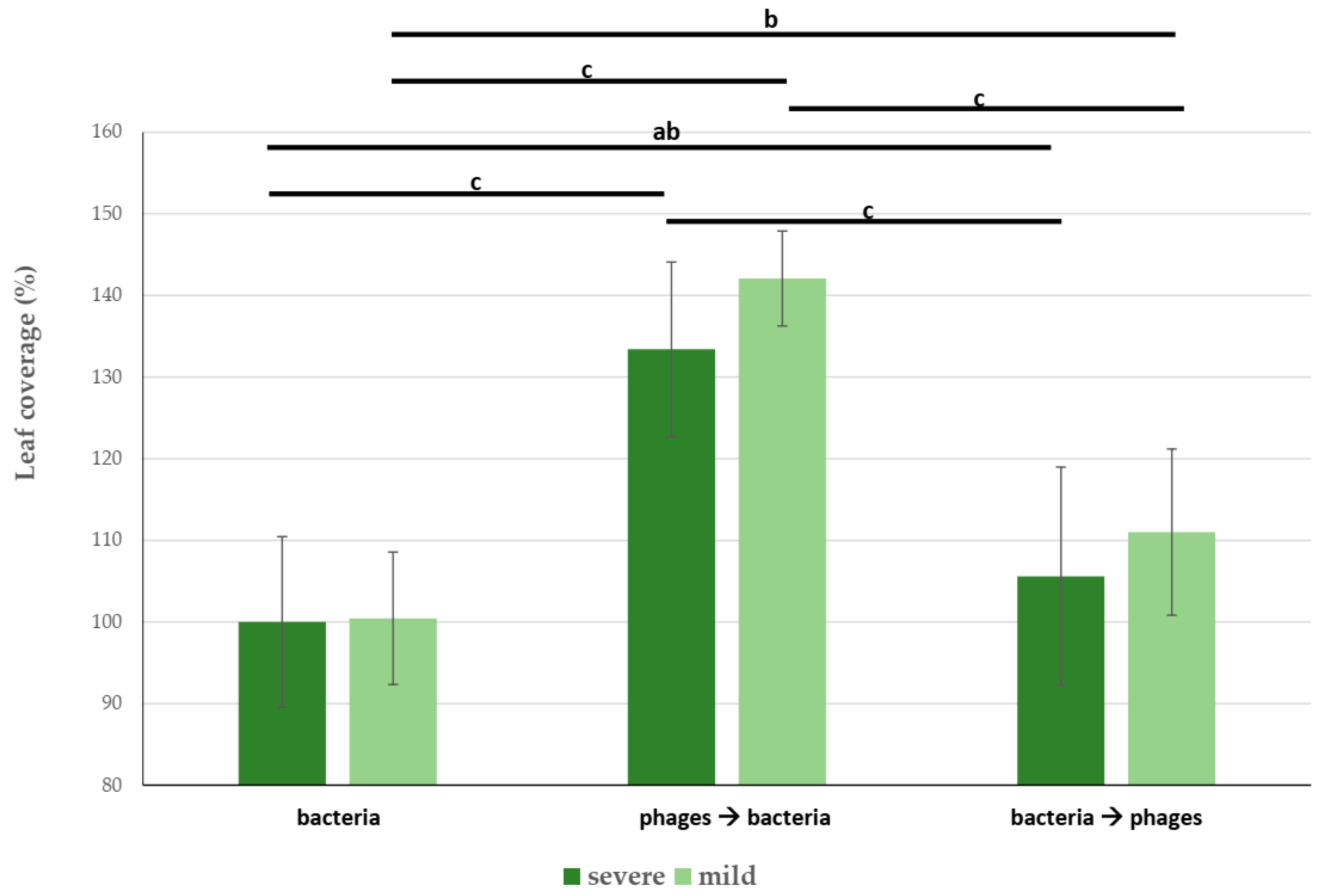
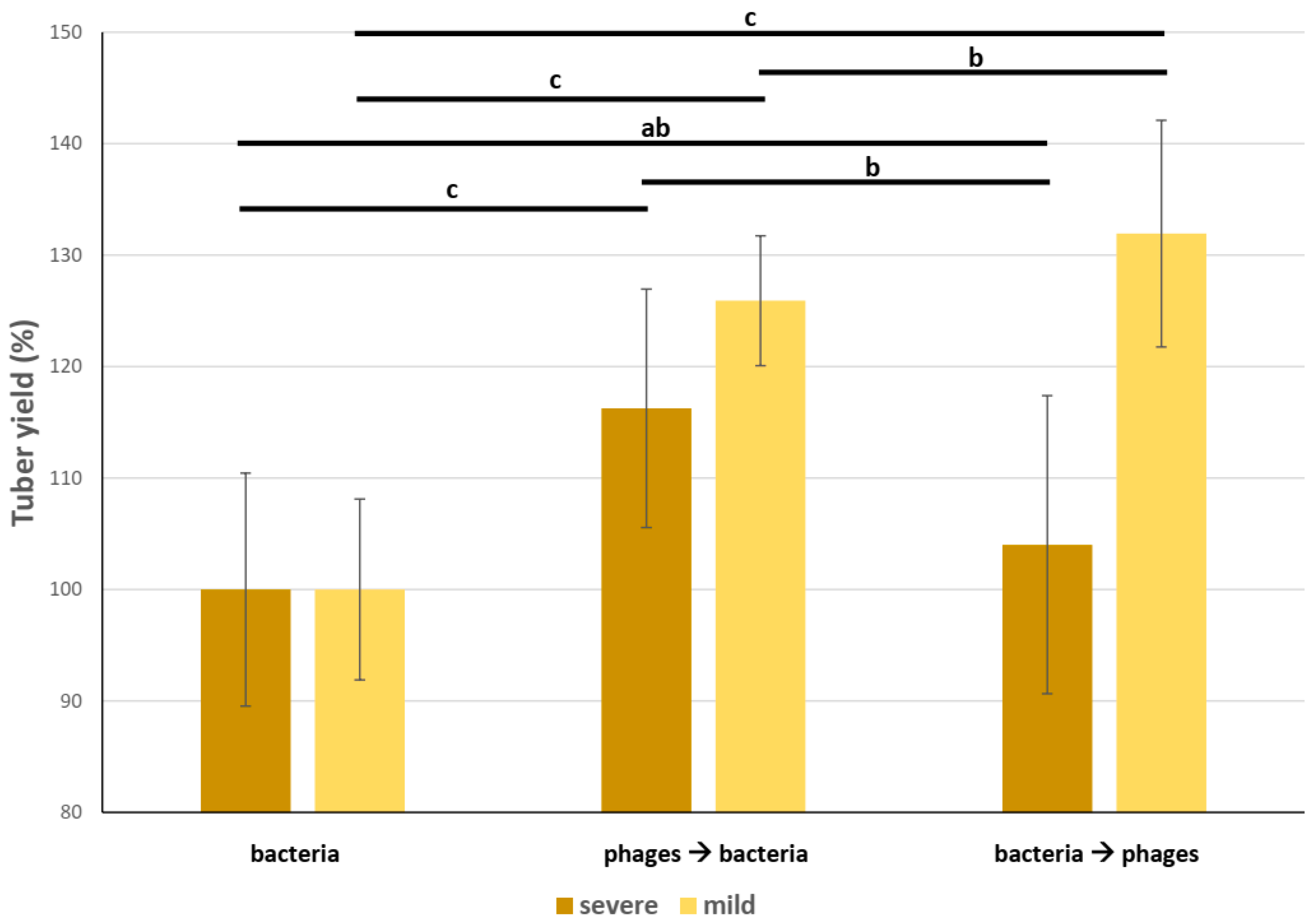
3.4. Field Trial
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van der Wolf, J.M.; Nijhuis, E.H.; Kowalewska, M.J.; Saddler, G.S.; Parkinson, N.; Elphinstone, J.G.; Pritchard, L.; Toth, I.K.; Lojkowska, E.; Potrykus, M.; et al. Dickeya solani sp. nov. a pectinolytic plant-pathogenic bacterium isolated from potato (Solanumtuberosum). Int. J. Syst. Evol. Microbiol. 2014, 64, 768–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology: Top 10 plant pathogenic bacteria. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [Green Version]
- Toth, I.K.; van der Wolf, J.M.; Saddler, G.; Lojkowska, E.; Hélias, V.; Pirhonen, M.; Tsror Lahkim, L.; Elphinstone, J.G. Dickeya species: An emerging problem for potato production in Europe: Dickeya spp. on potato in Europe. Plant Pathol. 2011, 60, 385–399. [Google Scholar] [CrossRef]
- Tsror, L.; Erlich, O.; Lebiush, S.; Hazanovsky, M.; Zig, U.; Slawiak, M.; Grabe, G.; van der Wolf, J.M.; van de Haar, J.J. Assessment of recent outbreaks of Dickeya sp. (syn. Erwinia chrysanthemi) slow wilt in potato crops in Israel. Eur. J. Plant. Pathol. 2009, 123, 311–320. [Google Scholar] [CrossRef]
- Laurila, J.; Hannukkala, A.; Nykyri, J.; Pasanen, M.; Hélias, V.; Garlant, L.; Pirhonen, M. Symptoms and yield reduction caused by Dickeya spp. strains isolated from potato and river water in Finland. Eur. J. Plant Pathol. 2010, 126, 249–262. [Google Scholar] [CrossRef]
- Beňo, F.; Horsáková, I.; Kmoch, M.; Petrzik, K.; Krátká, G.; Ševčík, R. Bacteriophages as a strategy to protect potato tubers against Dickeya dianthicola and Pectobacterium carotovorum soft rot. Microorganisms 2022, 10, 2369. [Google Scholar] [CrossRef]
- Vreugdenhil, D.; Bradshaw, J. (Eds.) Potato Biology and Biotechnology: Advances and Perspectives, 1st ed.; Elsevier: Oxford, UK; San Diego, CA, USA, 2007; ISBN 978-0-444-51018-1. [Google Scholar]
- Degefu, Y.; Potrykus, M.; Golanowska, M.; Virtanen, E.; Lojkowska, E. A new clade of Dickeya spp. plays a major role in potato blackleg outbreaks in north Finland: A new clade of Dickeya on potato in Finland. Ann. Appl. Biol. 2013, 162, 231–241. [Google Scholar] [CrossRef]
- Van der Wolf, J.; Krijger, M.; Mendes, O.; Kurm, V.; Gros, J. Natural infections of potato plants grown from minitubers with blackleg-causing soft rot Pectobacteriaceae. Microorganisms 2022, 10, 2504. [Google Scholar] [CrossRef]
- Moussa, H.B.; Claire, B.; Rochelle-Newall, E.; Fiorini, S.; Pédron, J.; Barny, M.-A. The diversity and abundance of soft rot pectobacteriaceae along the Durance River stream in the southeast of France revealed by multiple seasonal surveys. Phytopathology 2022, 112, 1676–1685. [Google Scholar] [CrossRef]
- Hugouvieux-Cotte-Pattat, N.; Brochier-Armanet, C.; Flandrois, J.-P.; Reverchon, S. Dickeya poaceiphila sp. nov., a plant-pathogenic bacterium isolated from sugar cane (Saccharum officinarum). Int. J. Syst. Evol. Microbiol. 2020, 70, 4508–4514. [Google Scholar] [CrossRef]
- Adriaenssens, E.M.; van Vaerenbergh, J.; Vandenheuvel, D.; Dunon, V.; Ceyssens, P.-J.; De Proft, M.; Kropinski, A.M.; Noben, J.P.; Maes, M.; Lavigne, R. T4-related bacteriophage LIMEstone isolates for the control of soft rot on potato caused by ‘Dickeya solani’. PLoS ONE 2012, 7, e33227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czajkowski, R.; Ozymko, Z.; Zwirowski, S.; Lojkowska, E. Complete genome sequence of a broad-host-range lytic Dickeya spp. bacteriophage ΦD5. Arch. Virol. 2014, 159, 3153–3155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Lofton, L.; Bamberg, J.; Swingle, B. Identification of resistance to Dickeya dianthicola soft rot in Solanum microdontum. Am. J. Potato Res. 2022, 9, 58–68. [Google Scholar] [CrossRef]
- Joshi, J.R.; Brown, K.; Charkowski, A.O.; Heuberger, A.L. Protease inhibitors from Solanum chacoense inhibit Pectobacterium virulence by reducing bacterial protease activity and motility. Mol. Plant Microbe. In. 2022, 35, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Austin, S.; Lojkowska, E.; Ehlenfeldt, M.K.; Kelman, A.; Helgeson, J.P. Fertile interspecific somatic hybrids of Solanum: A novel source of resistance to Erwinia soft rot. Phytopathology 1988, 78, 1216–1220. [Google Scholar] [CrossRef]
- Lebecka, R.; Śliwka, J.; Grupa-Urbańska, A.; Szajko, K.; Marzcewski, W. QTLs for potato tuber resistance to Dickeya solani are located on chromosomes II and IV. Plant Pathol. 2021, 70, 1745–1756. [Google Scholar] [CrossRef]
- Krzyzanowska, D.M.; Maciag, T.; Siwinska, J.; Krychowiak, M.; Jafra, S.; Czajkowski, R. Compatible mixture of bacterial antagonists developed to protect potato tubers from soft rot caused by Pectobacterium spp. and Dickeya spp. Plant Dis. 2019, 103, 1374–1382. [Google Scholar] [CrossRef]
- Jones, J.B.; Vallad, G.E.; Iriarte, F.B.; Obradović, A.; Wernsing, M.H.; Jackson, L.E.; Balogh, B.; Hong, J.C.; Momol, M.T. Considerations for using bacteriophages for plant disease control. Bacteriophage 2012, 2, e23857. [Google Scholar] [CrossRef] [Green Version]
- Buttimer, C.; McAuliffe, O.; Ross, R.P.; Hill, C.; O’Mahony, J.; Coffey, A. Bacteriophages and bacterial plant diseases. Front. Microbiol. 2017, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- Naureen, Z.; Dautaj, A.; Anpilogov, K.; Camilleri, G.; Dhuli, K.; Tanzi, B.; Maltese, P.E.; Cristofoli, F.; De Antoni, L.; Beccari, T.; et al. Bacteriophages presence in nature and their role in the natural selection of bacterial populations. Acta Biomed. 2020, 91, e2020024. [Google Scholar] [CrossRef]
- Czajkowski, R.; Ozymko, Z.; Lojkowska, E. Isolation and characterization of novel soilborne lytic bacteriophages infecting Dickeya spp. biovar 3 (‘D. Solani’). Plant Pathol. 2014, 63, 758–772. [Google Scholar] [CrossRef]
- Czajkowski, R.; Ozymko, Z.; de Jager, V.; Siwinska, J.; Smolarska, A.; Ossowicki, A.; Narajczyk, M.; Lojkowska, E. Genomic, proteomic and morphological characterization of two novel broad host lytic bacteriophages ΦPD10.3 and ΦPD23.1 infecting pectinolytic Pectobacterium spp. and Dickeya spp. PLoS ONE 2015, 10, e0119812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czajkowski, R.; Smolarska, A.; Ozymko, Z. The viability of lytic bacteriophage ΦD5 in potato-associated environments and its effect on Dickeya solani in potato (Solanum tuberosum L.) plants. PLoS ONE 2017, 12, e0183200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carstens, A.; Djurhuus, A.; Kot, W.; Jacobs-Sera, D.; Hatfull, G.; Hansen, L. Unlocking the potential of 46 new bacteriophages for biocontrol of Dickeya solani. Viruses 2018, 10, 621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrzik, K.; Vacek, J.; Brázdová, S.; Ševčík, R.; Koloniuk, I. Diversity of limestone bacteriophages infecting Dickeya solani isolated in the Czech Republic. Arch. Virol. 2021, 166, 1171–1175. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32(5), 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acid Res 2002, 130, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.P.; Varsani, A.; Roumagnac, P.; Botha, G.; Maslamoney, S.; Schwab, T.; Kelz, Z.; Kumar, V.; Murrell, B. RDP5: A computer program for analyzing recombination in, and removing signals of recombination from, nucleotide sequence datasets. Virus Evol. 2021, 7, veaa087. [Google Scholar] [CrossRef]
- Day, A.; Ahn, J.; Fang, X.; Salmond, G.P.C. Environmental bacteriophages of the emerging enterobacterial phytopathogen, Dickeya solani, show genomic conservation and capacity for horizontal gene transfer between their bacterial hosts. Front. Microbiol. 2017, 8, 1654. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.B.; Jackson, L.E.; Balogh, B.; Obradovic, A.; Iriarte, F.B.; Momol, M.T. Bacteriophages for plant disease control. Annu. Rev. Phytopathol. 2007, 45, 245–262. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar Sensitivity * | Bacteria Species | |
|---|---|---|
| Pectobacterium carotovorum | Dickeya solani | |
| 9.00–7.5 | - | - |
| 7.49–6.00 | Bella, Vysočina | Bella, Katy, Vysočina |
| 5.99–4.50 | ●DavidS, Katy, Magda, Mariannka, Monika, Primarosa, Suzan | ●DavidS, Dominika, Magda, Mariannka, Monika, Primarosa, ●VerneS, Suzan |
| 4.49–3.00 | Alice, Bohemia, Dominika, Jasmína, ●Jindra, Red Anna, ●VerneS, ●WestamylS | Alice, Bohemia, Jasmína, ●Jindra, Red Anna, ●WestamylS |
| 2.99–1.00 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrzik, K.; Vacek, J.; Kmoch, M.; Binderová, D.; Brázdová, S.; Lenz, O.; Ševčík, R. Field Use of Protective Bacteriophages against Pectinolytic Bacteria of Potato. Microorganisms 2023, 11, 620. https://doi.org/10.3390/microorganisms11030620
Petrzik K, Vacek J, Kmoch M, Binderová D, Brázdová S, Lenz O, Ševčík R. Field Use of Protective Bacteriophages against Pectinolytic Bacteria of Potato. Microorganisms. 2023; 11(3):620. https://doi.org/10.3390/microorganisms11030620
Chicago/Turabian StylePetrzik, Karel, Josef Vacek, Martin Kmoch, Denisa Binderová, Sára Brázdová, Ondřej Lenz, and Rudolf Ševčík. 2023. "Field Use of Protective Bacteriophages against Pectinolytic Bacteria of Potato" Microorganisms 11, no. 3: 620. https://doi.org/10.3390/microorganisms11030620
APA StylePetrzik, K., Vacek, J., Kmoch, M., Binderová, D., Brázdová, S., Lenz, O., & Ševčík, R. (2023). Field Use of Protective Bacteriophages against Pectinolytic Bacteria of Potato. Microorganisms, 11(3), 620. https://doi.org/10.3390/microorganisms11030620