

Association of ISVsa3 with Multidrug Resistance in Salmonella enterica Isolates from Cattle (Bos taurus)
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.1.1. S. enterica Isolates from Midwestern U.S. Feedyards from 2000 to 2001
2.1.2. Bacterial Strains Used as Controls for Multiplex qPCR (mqPCR) Assay for Detection of AMR Genes Associated with ICEPmu1 and ICEMh1
2.1.3. Isolates from Nebraska Veterinary Diagnostic Center from 2011 to 2020
2.2. Culture of Bacterial Strains and DNA Preparation
2.3. mqPCR
2.4. Endpoint PCR
2.5. WGS
2.6. Sequencing Quality Control and Genome Assembly
2.7. GenBank Accessions
2.8. Bioinformatic and Statistical Analyses
3. Results
3.1. Frequency of Antimicrobial Resistance (AMR) Genes Typically Associated with ICEMh1 and ICEMh1-like Integrative Conjugative Clements
3.2. Frequency of Salmonella Genomic Island 1 (SGI1) and SGI1 Variants
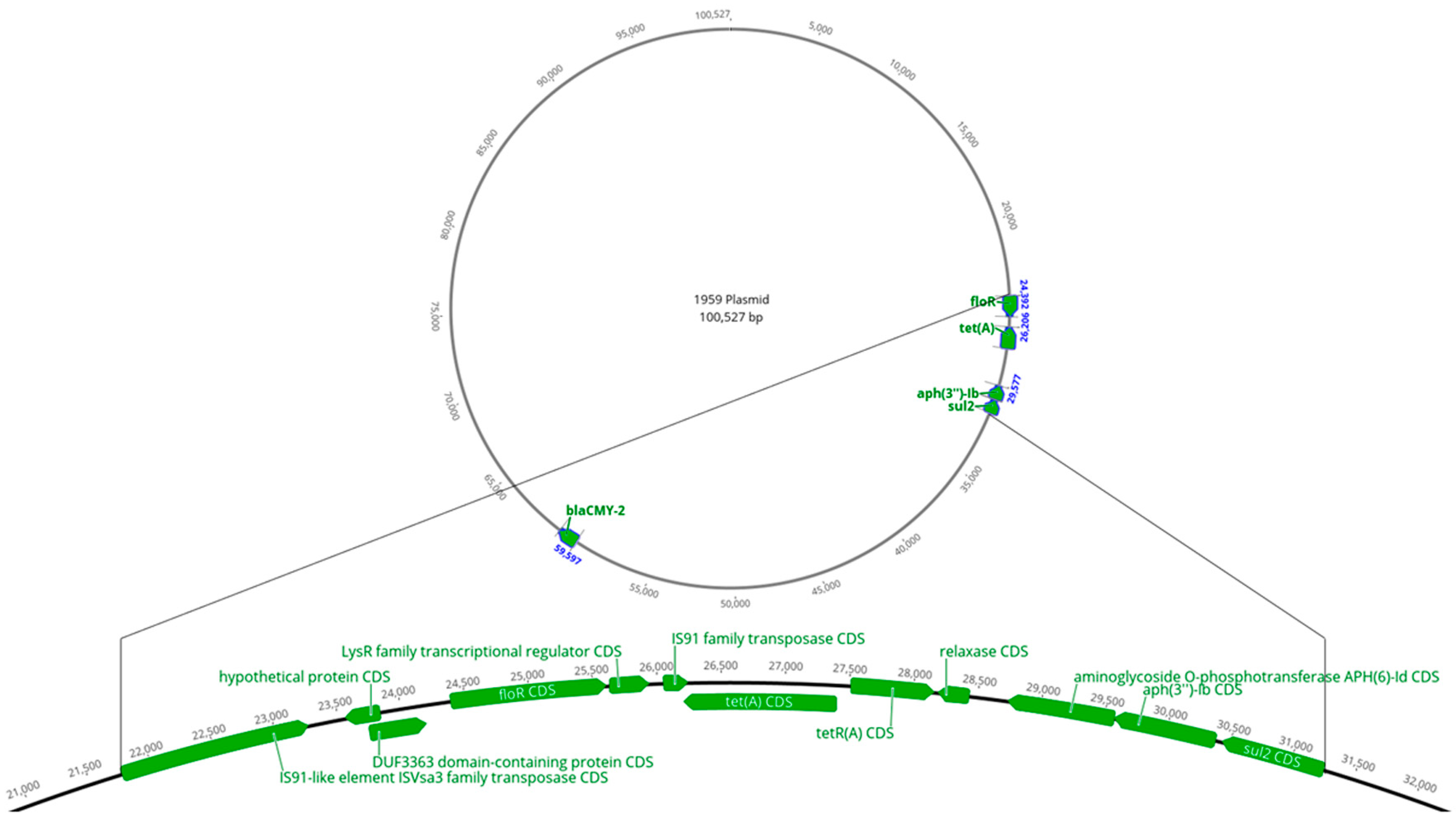
3.3. Other Mobile Genetic Elements, Their Genomic Locations, and MDR Association
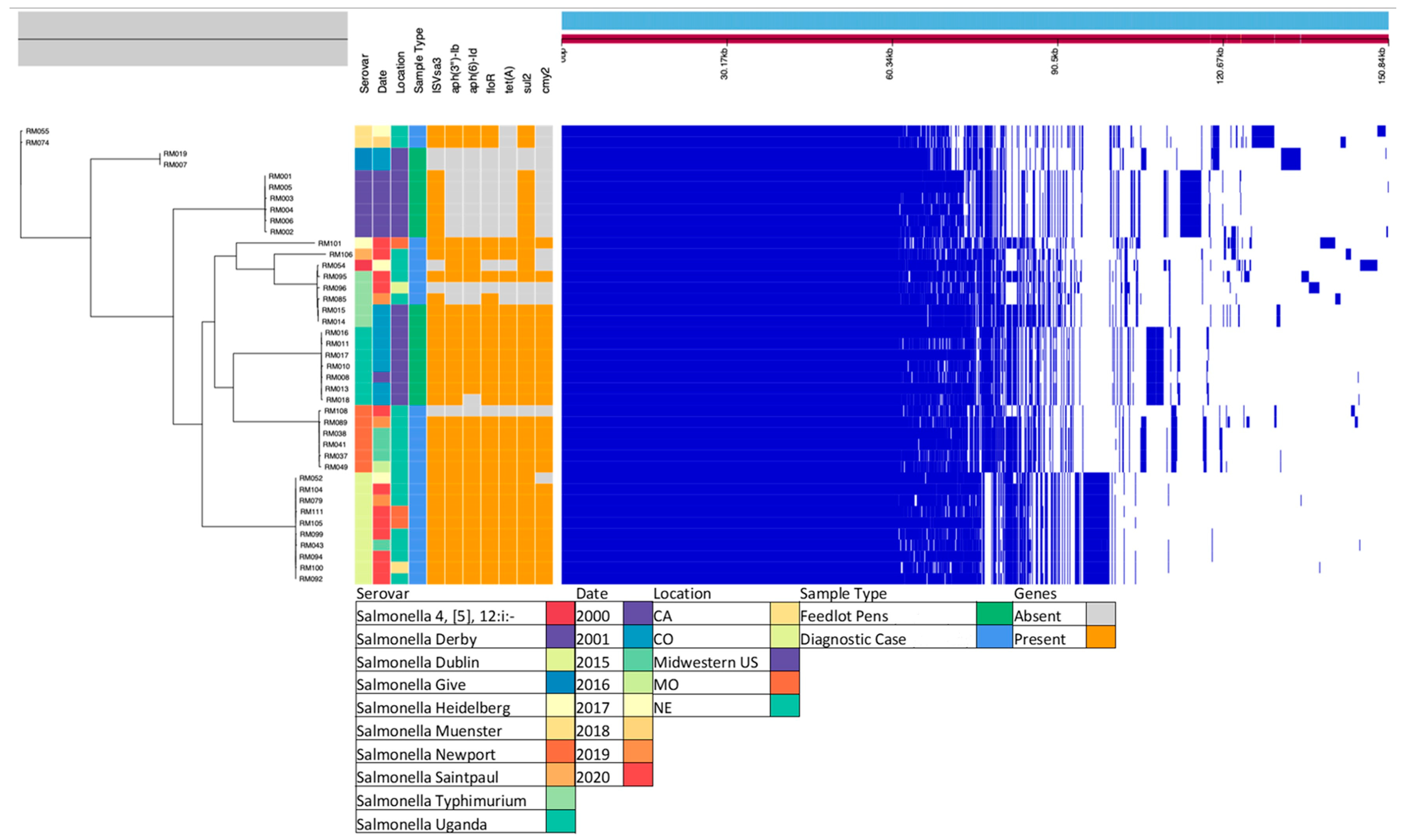
3.4. Core Gene Alignment and Metadata
3.5. ISVsa3 and Associated AMR Genetic Segment Alignment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CDC. Antibiotic Resistance Threats in the United States; U.S. Department of Health and Human Services, CDC: Atlanta, GA, USA, 2019. [Google Scholar]
- Richardson, L.C.; Bazaco, M.C.; Parker, C.C.; Dewey-Mattia, D.; Golden, N.; Jones, K.; Klontz, K.; Travis, C.; Kufel, J.Z.; Cole, D. An updated scheme for categorizing foods implicated in foodborne disease outbreaks: A tri-agency collaboration. Foodborne Pathog. Dis. 2017, 14, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Scharff, R.L. Food attribution and economic cost estimates for meat- and poultry-related illnesses. J. Food Prot. 2020, 83, 959–967. [Google Scholar] [CrossRef] [PubMed]
- DuPont, H.L. Clinical practice. Bacterial diarrhea. N. Engl. J. Med. 2009, 361, 1560–1569. [Google Scholar] [CrossRef] [PubMed]
- Shane, A.L.; Mody, R.K.; Crump, J.A.; Tarr, P.I.; Steiner, T.S.; Kotloff, K.; Langley, J.M.; Wanke, C.; Warren, C.A.; Cheng, A.C.; et al. 2017 Infectious Diseases Society of America clinical practice guidelines for the diagnosis and management of infectious diarrhea. Clin. Infect. Dis. 2017, 65, e45–e80. [Google Scholar] [CrossRef] [Green Version]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Burkhardt, J.E.; Walterspiel, J.N.; Schaad, U.B. Quinolone arthropathy in animals versus children. Clin. Infect. Dis. 1997, 25, 1196–1204. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.H.; Kim, E.Y.; Kim, Y.J. Systemic use of fluoroquinolone in children. Korean, J. Pediatr. 2013, 56, 196–201. [Google Scholar] [CrossRef]
- Medalla, F.; Hoekstra, R.M.; Whichard, J.M.; Barzilay, E.J.; Chiller, T.M.; Joyce, K.; Rickert, R.; Krueger, A.; Stuart, A.; Griffin, P.M. Increase in resistance to ceftriaxone and nonsusceptibility to ciprofloxacin and decrease in multidrug resistance among Salmonella strains, United States, 1996–2009. Foodborne Pathog. Dis. 2013, 10, 302–309. [Google Scholar] [CrossRef]
- Varma, J.K.; Molbak, K.; Barrett, T.J.; Beebe, J.L.; Jones, T.F.; Rabatsky-Her, T.; Smith, K.E.; Vugia, D.J.; Chang, H.G.; Angulo, F.J. Antimicrobial-resistant non-typhoidal Salmonella is associated with excess bloodstream infections and hospitalizations. J. Infect. Dis. 2005, 191, 554–561. [Google Scholar] [CrossRef] [Green Version]
- Barrow, P.A.; Jones, M.A.; Thomson, N. Salmonella. In Pathogenesis of Bacterial Infections in Animals; Wiley: Hoboken, NJ, USA, 2010; pp. 231–265. [Google Scholar] [CrossRef]
- Gutema, F.D.; Agga, G.E.; Abdi, R.D.; De Zutter, L.; Duchateau, L.; Gabriël, S. Prevalence and serotype diversity of Salmonella in apparently healthy cattle: Systematic review and meta-analysis of published studies, 2000–2017. Front. Vet. Sci. 2019, 6, 102. [Google Scholar] [CrossRef]
- Otto, S.J.G.; Ponich, K.L.; Cassis, R.; Goertz, C.; Peters, D.; Checkley, S.L. Antimicrobial resistance of bovine Salmonella enterica ssp. enterica isolates from the Alberta Agriculture and Forestry Disease Investigation Program (2006–2014). Can. Vet. J. 2018, 59, 1195–1201. [Google Scholar] [PubMed]
- Threlfall, E.J. Epidemic Salmonella typhimurium DT 104—A truly international multiresistant clone. J. Antimicrob. Chemother. 2000, 46, 7–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poppe, C.; Martin, L.; Muckle, A.; Archambault, M.; McEwen, S.; Weir, E. Characterization of antimicrobial resistance of Salmonella Newport isolated from animals, the environment, and animal food products in Canada. Can. J. Vet. Res. 2006, 70, 105–114. [Google Scholar]
- Fritz, H.M.; Pereira, R.V.; Toohey-Kurth, K.; Marshall, E.; Tucker, J.; Clothier, K.A. Salmonella enterica serovar Dublin from cattle in California from 1993–2019: Antimicrobial resistance trends of clinical relevance. Antibiotics 2022, 11, 1110. [Google Scholar] [CrossRef] [PubMed]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S. Mobile genetic elements associated with antimicrobial resistance. Clin. Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Li, X.; Xie, Y.; Bi, D.; Sun, J.; Li, J.; Tai, C.; Deng, Z.; Ou, H.Y. ICEberg 2.0: An updated database of bacterial integrative and conjugative elements. Nucleic Acids Res. 2019, 47, D660–D665. [Google Scholar] [CrossRef]
- Hall, R.M. Salmonella genomic islands and antibiotic resistance in Salmonella enterica. Future Microbiol. 2010, 5, 1525–1538. [Google Scholar] [CrossRef]
- Cameron, A.; Zaheer, R.; McAllister, T.A. Emerging variants of the integrative and conjugant element ICEMh1 in livestock pathogens: Structural insights, potential host range, and implications for bacterial fitness and antimicrobial therapy. Front. Microbiol. 2019, 10, 2608. [Google Scholar] [CrossRef]
- Snyder, E.; Credille, B.; Berghaus, R.; Giguère, S. Prevalence of multi drug antimicrobial resistance in Mannheimia haemolytica isolated from high-risk stocker cattle at arrival and two weeks after processing. J. Anim. Sci. 2017, 95, 1124–1131. [Google Scholar] [CrossRef]
- Snyder, E.R.; Alvarez-Narvaez, S.; Credille, B.C. Genetic characterization of susceptible and multi-drug resistant Mannheimia haemolytica isolated from high-risk stocker calves prior to and after antimicrobial metaphylaxis. Vet. Microbiol. 2019, 235, 110–117. [Google Scholar] [CrossRef]
- Snyder, E.; Credille, B. Mannheimia haemolytica and Pasteurella multocida in bovine respiratory disease: How are they changing in response to efforts to control them? Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 253–268. [Google Scholar] [CrossRef] [PubMed]
- Woolums, A.R.; Karisch, B.B.; Frye, J.G.; Epperson, W.; Smith, D.R.; Blanton, J., Jr.; Austin, F.; Kaplan, R.; Hiott, L.; Woodley, T.; et al. Multidrug resistant Mannheimia haemolytica isolated from high-risk beef stocker cattle after antimicrobial metaphylaxis and treatment for bovine respiratory disease. Vet. Microbiol. 2018, 221, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.R.; Moxley, R.A.; Clowser, S.L.; Folmer, J.D.; Hinkley, S.; Erickson, G.E.; Klopfenstein, T.J. Use of rope devices to describe and explain the feedlot ecology of Salmonella by time and place. Foodborne Pathog. Dis. 2005, 2, 61–69. [Google Scholar] [CrossRef]
- Alam, M.J.; Renter, D.; Taylor, E.; Mina, D.; Moxley, R.; Smith, D. Antimicrobial susceptibility profiles of Salmonella enterica serotypes recovered from pens of commercial feedlot cattle using different types of composite samples. Curr. Microbiol. 2009, 58, 354–359. [Google Scholar] [CrossRef]
- Dutta, E.; Loy, J.D.; Deal, C.A.; Wynn, E.L.; Clawson, M.L.; Clarke, J.; Wang, B. Development of a multiplex real-time PCR assay for predicting macrolide and tetracycline resistance associated with bacterial pathogens of bovine respiratory disease. Pathogens 2021, 10, 64. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. VET01. In Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated From Animals, 5th ed.; Clinical and Laboratory Standards Institute: Annapolis, MD, USA, 2018; p. 156. [Google Scholar]
- Clinical and Laboratory Standards Institute. VET01S. In Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated From Animals, 5th ed.; Clinical and Laboratory Standards Institute: Annapolis, MD, USA, 2020; p. 250. [Google Scholar]
- Sidjabat, H.E.; Paterson, D.L.; Adams-Haduch, J.M.; Ewan, L.; Pasculle, A.W.; Muto, C.A.; Tian, G.B.; Doi, Y. Molecular epidemiology of CTX-M-producing Escherichia coli isolates at a tertiary medical center in Western Pennsylvania. Antimicrob. Agents Chemother. 2009, 53, 4733–4739. [Google Scholar] [CrossRef] [Green Version]
- Vikram, A.L.; Miller, E.; Arthur, T.M.; Bosilevac, J.M.; Wheeler, T.L.; Schmidt, J.W. Similar levels of antimicrobial resistance in US food service ground beef products with and without a “raised without antibiotics” claim. J. Food Prot. 2018, 81, 2007–2018. [Google Scholar] [CrossRef] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinformatics 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar]
- Bortolaia, V.; Kaas, R.F.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.R.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadfield, J.; Croucher, N.J.; Goater, R.J.; Abudahab, K.; Aanensen, D.M.; Harris, S.R. Phandango: An interactive viewer for bacterial population genomics. Bioinformatics 2018, 34, 292–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MedCalc Software Ltd. Odds Ratio Calculator, Version 20.218. Available online: https://www.medcalc.org/calc/odds_ratio.php (accessed on 25 February 2023).
- Levent, G.; Schlochtermeier, A.; Ives, S.E.; Norman, K.N.; Lawhon, S.D.; Loneragan, G.H.; Anderson, R.C.; Vinasco, J.; Scott, H.M. Population dynamics of Salmonella enterica within beef cattle cohorts followed from single-dose metaphylactic antibiotic treatment until slaughter. Appl. Environ. Microbiol. 2019, 85, e01386-19. [Google Scholar] [CrossRef]
- Huguet, K.T.; Rivard, N.; Garneau, D.; Palanee, J.; Burrus, V. Replication of the Salmonella genomic island 1 (SGI1) triggered by helper IncC conjugative plasmids promotes incompatibility and plasmid loss. PLoS Genet. 2020, 16, e1008965. [Google Scholar] [CrossRef]
- Eyler, A.B.; M’ikanatha, M.M.; Xiaoli, L.; Dudley, E.G. Whole-genome sequencing reveals resistome of highly drug-resistant retail meat and human Salmonella Dublin. Zoonoses Public Health 2020, 67, 251–262. [Google Scholar] [CrossRef]
- He, T.; Wang, R.; Liu, D.; Walsh, T.R.; Zhang, R.; Yuan, L.; Yuebin, K.; Quanjiang, J.; Ruicheng, W.; Zhihai, L.; et al. Emergence of plasmid-mediated high-level tigecycline resistane in animals and humans. Nat. Microbiol. 2019, 4, 1450–1456. [Google Scholar] [CrossRef]
- Abdelhamed, H.; Ramachandran, R.; Ozdemir, O.; Waldbieser, G.; Lawrence, M.L. Characterization of a novel conjugative plasmid in Edwardsiella piscicida strain MS-18-199. Front. Cell. Infect. Microbiol. 2019, 9, 404. [Google Scholar] [CrossRef]
- Toleman, M.A.; Bennett, P.M.; Walsh, T.R. Common regions e.g., orf513 and antibiotic resistance: IS91-like elements evolving complex class 1 integrons. J. Antimicrob. Chemother. 2006, 58, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Toleman, M.A.; Bennett, P.M.; Walsh, T.R. ISCR elements: Novel gene-capturing systems of the 21st century? Microbiol. Mol. Biol. Rev. 2006, 70, 296–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toleman, M.A.; Walsh, T.R. ISCR elements are key players in IncA/C plasmid evolution. Antimicrob. Agents Chemother. 2010, 54, 3534. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Isolate | Date | Feedyard /Pen 1 | Sample 2 | Serotype | NARMS (2006) 3 | BOPO6/7F (2020) 4 | WGS 6 |
|---|---|---|---|---|---|---|---|
| RM001 | 19 September 2000 | 1/655c | Rope (R6) | Derby | FIS | ND 5 | + |
| RM002 | 19 September 2000 | 1/655c | Water (W2) | Derby | FIS | ND | + |
| RM003 | 30 October 2000 | 1/669 | Rope (R3) | Derby | FIS | ND | + |
| RM004 | 30 October 2000 | 1/669 | Rope (R6) | Derby | FIS | ND | + |
| RM005 | 30 October 2000 | 1/655b | Rope (R1) | Derby | FIS | ND | + |
| RM006 | 30 October 2000 | 1/655b | Rope (R3) | Derby | FIS | ND | + |
| RM007 | 17 September 2001 | 2/822b | Water (W2) | Give | CHL, FIS, KAN, NAL, STR, TET | Pan-susceptible | + |
| RM008 | 17 September 2001 | 3/154 | Feces (F1A) | Uganda | AMP, AUG2, AXO, CHL, FIS, FOX, STR, TET, XNL | AMP, FFN, SDM, TET, XNL | + |
| RM009 | 17 September 2001 | 3/154 | Feces (F1A) | Uganda | AMP, AUG2, AXO, CHL, FIS, FOX, STR, TET, XNL | AMP, FFN, SDM, TET, XNL | - |
| RM010 | 17 September 2001 | 3/154 | Feces (F1A) | Uganda | AMP, AUG2, AXO, CHL, FIS, FOX, STR, TET, XNL | AMP, FFN, SDM, TET, XNL | + |
| RM011 | 17 September 2001 | 3/157 | Rope (R1) | Uganda | AMP, AUG2, AXO, CHL, FIS, FOX, STR, TET, XNL | AMP, FFN, SDM, TET, XNL | + |
| RM012 | 17 September 2001 | 3/157 | Rope (R1) | Uganda | AMP, AUG2, AXO, CHL, FIS, FOX, STR, TET, XNL | AMP, FFN, SDM, TET, XNL | - |
| RM013 | 17 September 2001 | 3/157 | Rope (R1) | Uganda | AMP, AUG2, AXO, CHL, FIS, FOX, STR, TET, XNL | AMP, FFN, SDM, TET, XNL | + |
| RM014 | 17 September 2001 | 3/157 | Rope (R4) | Typhimurium | AMP, AUG2, AXO, CHL, FIS, STR, TET, XNL | AMP, FFN, SDM, TET, XNL | + |
| RM015 | 17 September 2001 | 3/157 | Rope (R4) | Typhimurium | AMP, AUG2, AXO, CHL, FIS, FOX, STR, TET, XNL | AMP, FFN, SDM, TET, XNL | + |
| RM016 | 17 September 2001 | 3/157 | Water (W2) | Uganda | AMP, AUG2, AXO, CHL, FIS, FOX, STR, TET, XNL | AMP, FFN, SDM, TET, XNL | + |
| RM017 | 17 September 2001 | 3/157 | Water (W2) | Uganda | AMP, AUG2, AXO, CHL, CIP, FIS, GEN, STR, TET, XNL | AMP, FFN, SDM, TET, XNL | + |
| RM018 | 17 September 2001 | 3/157 | Water (W2) | Uganda | AMP, AUG2, AXO, CHL, FIS, FOX, STR, TET, XNL | AMP, FFN, SDM, TET, XNL | + |
| RM019 | 23 October 2001 | 2/822b | Rope (R1) | Give | AMP, AUG2, AXO, CHL, FIS, STR, TET, XNL | Pan-susceptible | + |
| Isolate | Date | Signalment | History/Pathology | Sample 1 | Serotype 2 | AMR Phenotype 3 | WGS 4 |
|---|---|---|---|---|---|---|---|
| RM093 | 12 September 2019 | Cow | Diarrhea, hepatitis | Small and large intestines | Anatum | DANO, ENRO, FFN, SDM, OXY, XNL | - |
| RM043 | 20 May 2015 | Calf | Bloody diarrhea, pneumonia, septicemia | Pool of small intestine and lung | Dublin | CTET, FFN, OXY, SDM, XNL | + |
| RM052 | 4 April 2017 | No information | Pneumonia | Lung | Dublin | CTET, FFN, OXY, SDM | + |
| RM079 | 20 July 2018 | Dairy calf | Pneumonia | Lung | Dublin | CTET, FFN, OXY, SDM, XNL | + |
| RM092 | 26 July 2019 | Calf (9–10 week-old) | Pneumonia, septicemia | Lung | Dublin | ENRO, FFN, OXY, SDM, XNL | + |
| RM094 | 17 September 2019 | Dairy calf (1–4 week-old) | Septicemia, pneumonia | Liver, lung, small intestine | Dublin | DANO, ENRO, FFN, OXY, XNL | + |
| RM099 | 7 November 2019 | Calf | Diarrhea, pneumonia | Pooled lung, small and large intestines | Dublin | DANO, ENRO, FFN, OXY, XNL | + |
| RM100 | 15 January 2020 | No information | No information | Bacterial isolate | Dublin | DANO, ENRO, FFN, SDM, TET, XNL | + |
| RM104 | 3 April 2020 | Calf (1-week-old) | Septicemia, pneumonia | Feces | Dublin | FFN, OXY, SDM, XNL | + |
| RM105 | 7 April 2020 | Calf | Septicemia, pneumonia | Lung and small intestine | Dublin | FFN, OXY, SDM, XNL | + |
| RM111 | 11 June 2020 | No information | Diarrhea, pneumonia | Feces | Dublin | FFN, OXY, SDM, XNL | + |
| RM101 | 12 February 2020 | Calf (3–7-day-old) | Diarrhea, enteritis | Large intestine | Heidelberg | DANO, ENRO, OXY, SDM, XNL | + |
| RM055 | 11 May 2017 | No information | No information | Feces | Muenster | DANO, ENRO, FFN, SDM | + |
| RM074 | 16 May 2018 | No information | Diarrhea | Feces | Muenster | DANO, ENRO, FFN, SDM | + |
| RM037 | 10 April 2015 | No information | No information | Feces | Newport | CTET, FFN, OXY, SDM, XNL | + |
| RM038 | 14 April 2015 | Feedlot cattle | Bloody diarrhea | Feces | Newport | CTET, FFN, OXY, SDM, XNL | + |
| RM041 | 28 April 2015 | Feedlot cattle | Bloody diarrhea | Feces | Newport | CTET, FFN, OXY, SDM, XNL | + |
| RM049 | 15 October 2015 | Weaned calf | Bloody diarrhea | Feces | Newport | CTET, FFN, OXY, SDM, XNL | + |
| RM089 | 13 June 2019 | Calf | No information | Feces | Newport | FFN, OXY, SDM, XNL | + |
| RM106 | 14 April 2020 | Neonatal calf | Diarrhea, colitis | Large intestine | Saintpaul | FFN, OXY, SDM | + |
| RM086 | 5 April 2019 | Calf | Diarrhea | Feces | Species | DANO, ENRO, FFN, OXY, SDM | - |
| RM095 | 9 October 2019 | Bull (3-year-old) | Abomasitis, duodenitis, peritonitis (septicemia) | Liver, gallbladder, lymph node | Typhimurium | FFN, OXY, SDM, XNL | + |
| Number of AMR Genes 1 | Isolates Positive 2 | Plasmid 3 | IncC Plasmid 4 | blaCMY-2 Positive Plasmid | ||
|---|---|---|---|---|---|---|
| Isolates Positive 5 | Plasmid 6 | IncC Plasmid 7 | ||||
| 1 | 7/41 (17.0%) | 6/7 (85.7%) | 0/6 (0.0%) | 0/6 (0.0%) | NA | NA |
| 2 | 0/41 (0.0%) | NA | NA | NA | NA | NA |
| 3 | 0/41 (0.0%) | NA | NA | NA | NA | NA |
| 4 | 3/41 (7.3%) | 3/3 (100.0%) | 1/3 (33.3%) | 1/3 (33.3%) | 1/3 (33.3%) | 1/3 (33.3%) |
| 5 | 26/41 (63.4%) | 24/26 (90.0%) | 19/24 (79.2%) | 22/26 (84.6%) | 21/22 (95.4%) | 19/21 (90.5%) |
| Gene | Isolates Positive 1 | Plasmid 2 | IncC Plasmid 3 |
|---|---|---|---|
| ISVsa3 | 36/41 (87.8%) | 33/36 (91.7%) | 20/33 (60.6%) |
| floR | 30/36 (73.2%) | 27/30 (90.0%) | 20/27 (74.1%) |
| tet(A) | 27/36 (65.9%) | 25/27 (92.6%) | 20/25 (80.0%) |
| aph(3″)-Ib | 29/36 (70.7%) | 27/29 (93.1%) | 20/27 (74.1%) |
| aph(6)-Id | 28/36 (68.3%) 4 | 26/28 (92.9%) 4 | 19/26 (73.1%) 4 |
| sul2 | 35/36 (87.8%) | 33/36 (91.7%) | 19/33 (57.6%) |
| blaCMY-2 | 25/36 (61.0%) 5 | 23/25 (92.0%) 5 | 20/23 (87.0%) 5 |
| MDR Phenotype 1 | ||||
|---|---|---|---|---|
| ISVsa3 Genotype 2 | Positive | Negative | Total | |
| Positive | 31 | 6 | 37 | |
| Negative | 2 | 72 | 74 | |
| Total | 33 | 78 | 111 | |
| OR 3 | OR = [(31 × 72)/(6 × 2)]/12 = 2232/12 = 186.00 (CI = 35.55–973.15; z statistic = 6.190; p < 0.0001) | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lewis, G.L.; Fenton, R.J.; Moriyama, E.N.; Loy, J.D.; Moxley, R.A. Association of ISVsa3 with Multidrug Resistance in Salmonella enterica Isolates from Cattle (Bos taurus). Microorganisms 2023, 11, 631. https://doi.org/10.3390/microorganisms11030631
Lewis GL, Fenton RJ, Moriyama EN, Loy JD, Moxley RA. Association of ISVsa3 with Multidrug Resistance in Salmonella enterica Isolates from Cattle (Bos taurus). Microorganisms. 2023; 11(3):631. https://doi.org/10.3390/microorganisms11030631
Chicago/Turabian StyleLewis, Gentry L., Robert J. Fenton, Etsuko N. Moriyama, John Dustin Loy, and Rodney A. Moxley. 2023. "Association of ISVsa3 with Multidrug Resistance in Salmonella enterica Isolates from Cattle (Bos taurus)" Microorganisms 11, no. 3: 631. https://doi.org/10.3390/microorganisms11030631
APA StyleLewis, G. L., Fenton, R. J., Moriyama, E. N., Loy, J. D., & Moxley, R. A. (2023). Association of ISVsa3 with Multidrug Resistance in Salmonella enterica Isolates from Cattle (Bos taurus). Microorganisms, 11(3), 631. https://doi.org/10.3390/microorganisms11030631