


Total Osteopontin and Its Isoform OPN4 Are Differently Expressed in Respiratory Samples during Influenza A(H1N1)pdm09 Infection and Progression

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Respiratory Samples and Case Clinical Classification
2.2. Diagnosis of Influenza A(H1N1)pdm09
2.3. OPN Detection in the Respiratory Samples
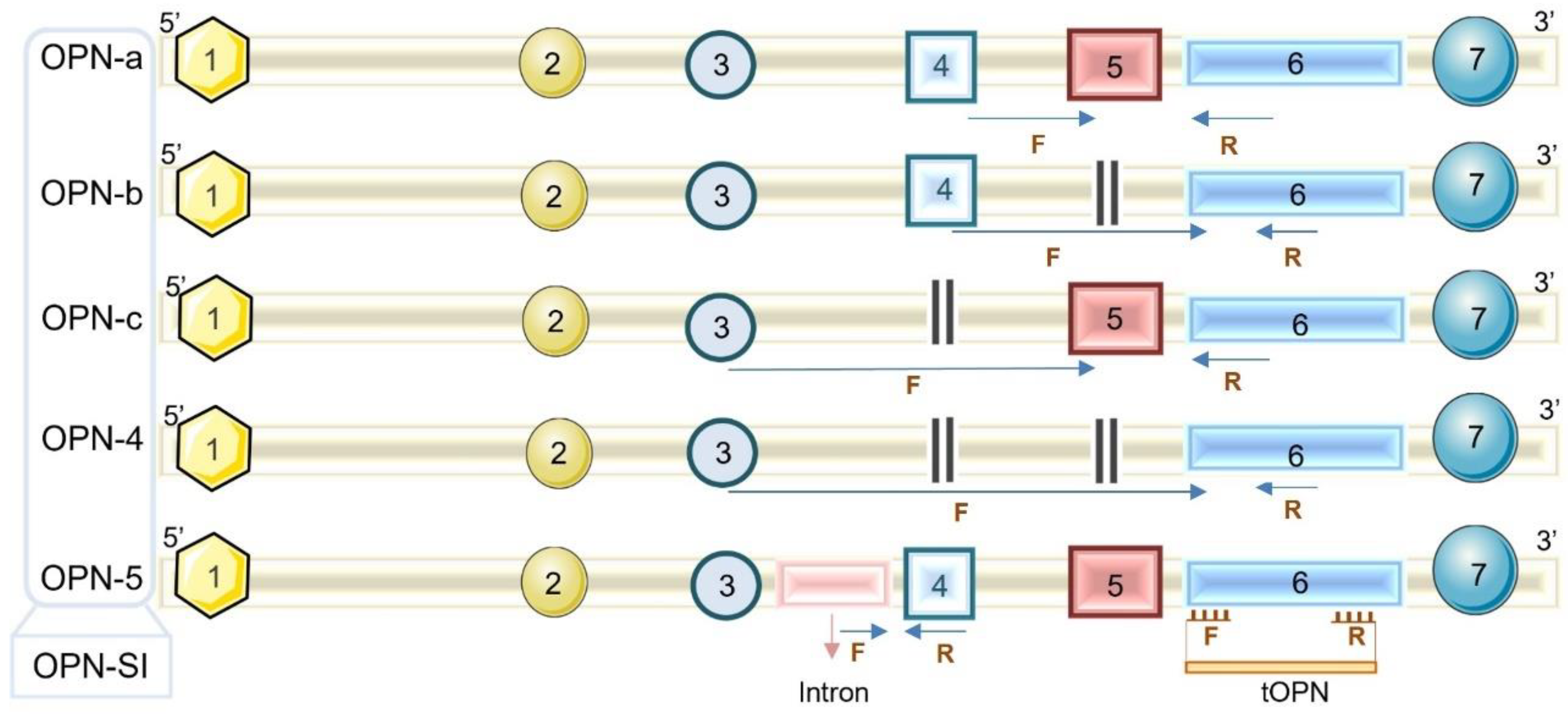
2.4. OPN Transcript Levels
2.5. Statistical Analysis
3. Results
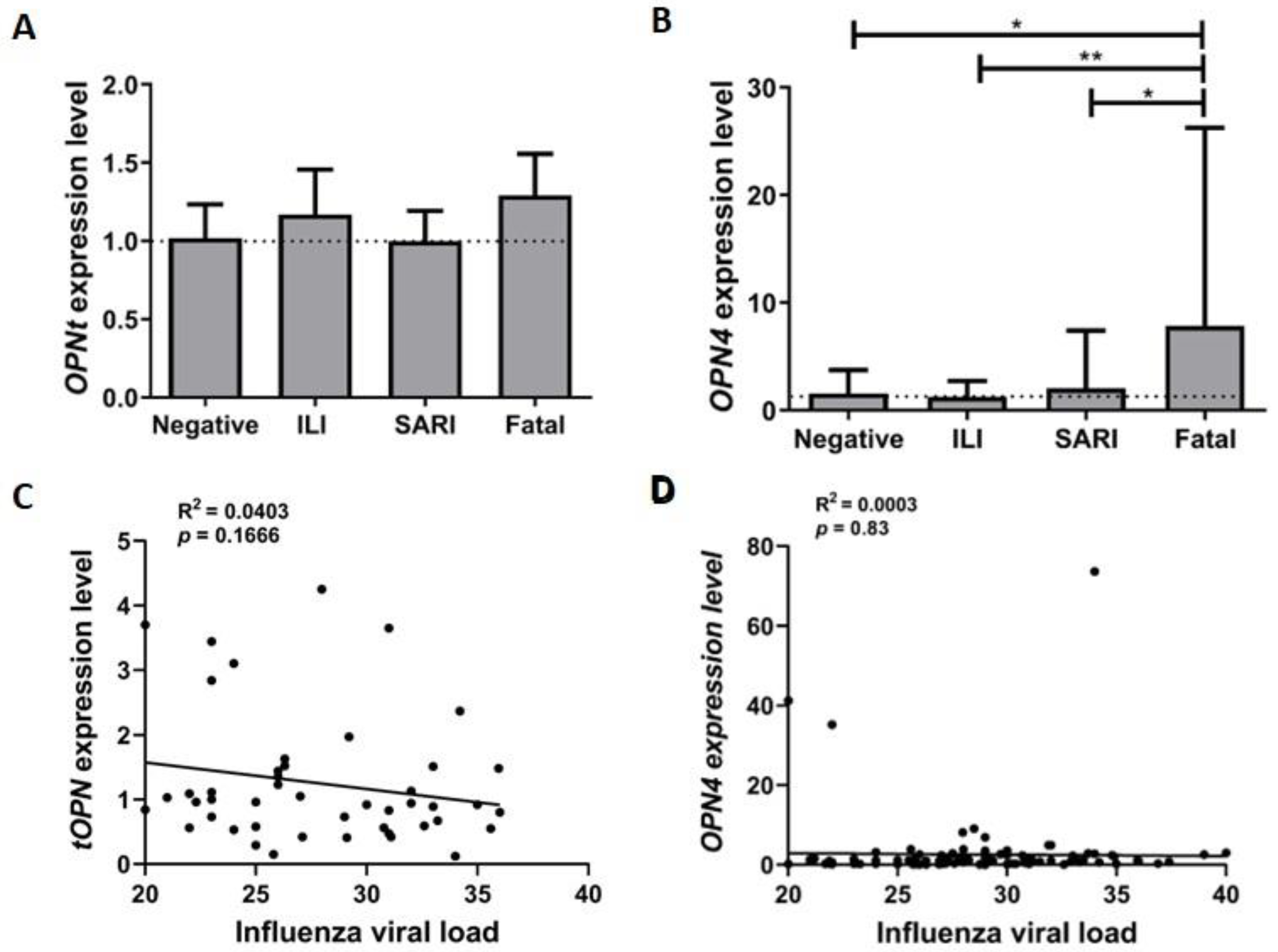
3.1. tOPN and OPN4 Transcripts Are more Prevalent in Respiratory Samples from Influenza A(H1N1)pdm09 Cases
3.2. OPN4 Expression Level Is Increased in Respiratory Samples from Fatal Influenza A(H1N1)pdm09 Patients
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Danielle Iuliano, A.; Roguski, K.M.; Chang, H.H.; Muscatello, D.J.; Palekar, R.; Tempia, S.; Cohen, C.; Michael Gran, J.; Schanzer, D.; Cowling, B.J.; et al. Estimates of global seasonal influenza-associated respiratory mortality: A modelling study HHS Public Access. Lancet 2018, 391, 1285–1300. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Global Influenza Strategy 2019–2030; World Health Organization: Geneva, Switzerland, 2019.
- World Helth Organization. Influenza (Seasonal). Available online: https://www.who.int/en/news-room/fact-sheets/detail/influenza-(seasonal) (accessed on 2 July 2019).
- Whitaker, J.A.; von Itzstein, M.S.; Poland, G.A. Strategies to maximize influenza vaccine impact in older adults. Vaccine 2018, 36, 5940–5948. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Goicoechea, S.; Kumar, S.; Pearce, C.M.; Durvasula, R.; Kempaiah, P.; Rathi, B. Poonam Oseltamivir analogs with potent anti-influenza virus activity. Drug Discov. Today 2020, 25, 1389–1402. [Google Scholar] [CrossRef]
- Capão, A.; Aguiar-Oliveira, M.L.; Caetano, B.C.; Neves, T.K.; Resende, P.C.; Almeida, W.A.F.; Miranda, M.D.; Martins-Filho, O.A.s.; Brown, D.; Siqueira, M.M.; et al. Analysis of Viral and Host Factors on Immunogenicity of 2018, 2019, and 2020 Southern Hemisphere Seasonal Trivalent Inactivated Influenza Vaccine in Adults in Brazil. Viruses 2022, 14, 1692. [Google Scholar] [CrossRef]
- Sousa, T.D.C.; Martins, J.S.C.C.; Miranda, M.D.; Garcia, C.C.; Resende, P.C.; Santos, C.A.D.; Debur, M.D.C.; Rodrigues, R.R.; Cavalcanti, A.C.; Gregianini, T.S.; et al. Low prevalence of influenza A strains with resistance markers in Brazil during 2017–2019 seasons. Front. Public Health 2022, 10, 944277. [Google Scholar] [CrossRef]
- International Committee on Taxonomy of Viruses (ICTV). Available online: https://talk.ictvonline.org/taxonomy/ (accessed on 7 January 2020).
- Dou, D.; Revol, R.; Östbye, H.; Wang, H.; Daniels, R. Influenza A virus cell entry, replication, virion assembly and movement. Front. Immunol. 2018, 9, 1–17. [Google Scholar] [CrossRef]
- Takashita, E.; Murakami, S.; Matsuzaki, Y.; Fujisaki, S.; Morita, H.; Nagata, S.; Katayama, M.; Mizuta, K.; Nishimura, H.; Watanabe, S.; et al. Antiviral Susceptibilities of Distinct Lineages of Influenza C and D Viruses. Viruses 2023, 15, 244. [Google Scholar] [CrossRef]
- Kim, Y.H.; Hong, K.J.; Kim, H.; Nam, J.H. Influenza vaccines: Past, present, and future. Rev. Med. Virol. 2022, 32, e2243. [Google Scholar] [CrossRef]
- Chen, Q.; Liu, Y.; Lu, A.; Ni, K.; Xiang, Z.; Wen, K.; Tu, W. Influenza virus infection exacerbates experimental autoimmune encephalomyelitis disease by promoting type I T cells infiltration into central nervous system. J. Autoimmun. 2017, 77, 1–10. [Google Scholar] [CrossRef]
- Writing Committee of the WHO Consultation on Clinical Aspects of Pandemic (H1N1)2009 Influenza Clinical Aspects of Pandemic 2009 Influenza. N. Engl. J. Med. 2010, 18, 1708–1719.
- Ilyicheva, T.; Durymanov, A.; Susloparov, I.; Kolosova, N.; Goncharova, N.; Svyatchenko, S.; Petrova, O.; Bondar, A.; Mikheev, V.; Ryzhikov, A. Fatal cases of seasonal influenza in Russia in 2015–2016. PLoS ONE 2016, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Matos, A.R.; Martins, J.S.; Maria de Lourdes, A.O.; Garcia, C.C.; Siqueira, M.M. Human CCR5Δ32 (rs333) polymorphism has no influence on severity and mortality of influenza A(H1N1)pdm09 infection in Brazilian patients from the post pandemic period. Infect. Genet. Evol. 2019, 67, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Martins, J.S.C.; Oliveira, M.L.A.; Garcia, C.C.; Siqueira, M.M.; Matos, A.R. Investigation of Human IFITM3 Polymorphisms rs34481144A and rs12252C and Risk for Influenza A(H1N1)pdm09 Severity in a Brazilian Cohort. Front. Cell. Infect. Microbiol. 2020, 10, 352. [Google Scholar] [CrossRef] [PubMed]
- Van Goethem, N.; Danwang, C.; Bossuyt, N.; Van Oyen, H.; Roosens, N.H.C.; Robert, A. A systematic review and meta-analysis of host genetic factors associated with influenza severity. BMC Genom. 2021, 22, 912. [Google Scholar] [CrossRef]
- Icer, M.A.; Gezmen-Karadag, M. The multiple functions and mechanisms of osteopontin. Clin. Biochem. 2018, 59, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Bastos, A.C.S.D.F.; Gomes, A.V.P.; Silva, G.R.; Emerenciano, M.; Ferreira, L.B.; Gimba, E.R.P. The Intracellular and Secreted Sides of Osteopontin and Their Putative Physiopathological Roles. Int. J. Mol. Sci. 2023, 24, 2429. [Google Scholar] [CrossRef]
- Bortell, N.; Flynn, C.; Conti, B.; Fox, H.S.; Marcondes, M.C.G. Osteopontin Impacts West Nile virus Pathogenesis and Resistance by Regulating Inflammasome Components and Cell Death in the Central Nervous System at Early Time Points. Mediators Inflamm. 2017, 2017, 1–12. [Google Scholar] [CrossRef]
- Sampayo-Escobar, V.; Green, R.; Cheung, M.B.; Bedi, R.; Mohapatra, S.; Mohapatra, S.S. Osteopontin plays a pivotal role in increasing severity of respiratory syncytial virus infection. PLoS ONE 2018, 13, e0192709. [Google Scholar] [CrossRef]
- Wang, J.; Li, X.; Wang, Y.; Li, Y.; Shi, F.; Diao, H. Osteopontin aggravates acute lung injury in influenza virus infection by promoting macrophages necroptosis. Cell Death Discov. 2022, 8, 97. [Google Scholar] [CrossRef]
- Varım, C.; Demirci, T.; Cengiz, H.; Hacıbekiroğlu, İ.; Tuncer, F.B.; Çokluk, E.; Toptan, H.; Karabay, O.; Yıldırım, İ. Relationship between serum osteopontin levels and the severity of COVID-19 infection. Wien. Klin. Wochenschr. 2021, 133, 298–302. [Google Scholar] [CrossRef]
- Moreno-Altamirano, M.M.B.; Romano, M.; Legorreta-Herrera, M.; Sánchez-García, F.J.; Colston, M.J. Gene expression in human macrophages infected with dengue virus serotype-2. Scand. J. Immunol. 2004, 60, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Gimba, E.R.P.; Brum, M.C.M.; De Moraes, G.N. Full-length osteopontin and its splice variants as modulators of chemoresistance and radioresistance (Review). Int. J. Oncol. 2019, 54, 420–430. [Google Scholar] [CrossRef]
- Kleinman, J.G.; Wesson, J.A.; Hughes, J. Osteopontin and Calcium Stone Formation. Nephron Physiol. 2004, 98, p43–p47. [Google Scholar] [CrossRef]
- Christensen, B.; Petersen, T.E.; Sørensen, E.S. Post-translational modification and proteolytic processing of urinary osteopontin. Biochem. J. 2008, 411, 53–61. [Google Scholar] [CrossRef]
- Bastos, A.C.S.F.; Blunck, C.B.; Emerenciano, M.; Gimba, E.R.P. Osteopontin and their roles in hematological malignancies: Splice variants on the new avenues. Cancer Lett. 2017, 408, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Sousa, T.D.C.; Martins, J.S.C.C.; Miranda, M.D.; Garcia, C.C.; Resende, P.C.; Santos, C.A.; Debur, M.D.C.; Rodrigues, R.R.; Cavalcanti, A.C.; Gregianini, T.S.; et al. Detection and localization of surgically resectable cancers with a multi-analyte blood test. Physiol. Behav. 2017, 359, 926–930. [Google Scholar] [CrossRef]
- Silva, G.R.; Mattos, D.S.; Bastos, A.C.F.; Viana, B.P.P.B.; Brum, M.C.M.; Ferreira, L.B.; Gimba, E.R.P. Osteopontin-4 and Osteopontin-5 splice variants are expressed in several tumor cell lines. Mol. Biol. Rep. 2020, 47, 8339–8345. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, J.; Sarkar-Dutta, M.; McRae, S.; Ramachandran, A.; Kumar, B.; Waris, G. Osteopontin Regulates Hepatitis C Virus (HCV) Replication and Assembly by Interacting with HCV Proteins and Lipid Droplets and by Binding to Receptors αVβ3 and CD44. J. Virol. 2018, 92, e02116-17. [Google Scholar] [CrossRef]
- Liu, H.B.; Chen, Q.Y.; Wang, X.Y.; Zhang, L.J.; Hu, L.P.; Harrison, T.J.; Wang, C.; Fang, Z.L. Infection with Hepatitis B Virus May Increase the Serum Concentrations of Osteopontin. Intervirology 2021, 64, 126–134. [Google Scholar] [CrossRef]
- Zhu, Y.; Wei, Y.; Chen, J.; Cui, G. Osteopontin Exacerbates Pulmonary Damage in Influenza-Induced Lung Injury. Jpn. J. Infect. Dis. 2015, 68, 467–473. [Google Scholar] [CrossRef]
- Ding, X.M.; Wang, Y.F.; Lyu, Y.; Zou, Y.; Wang, X.; Ruan, S.M.; Wu, W.H.; Liu, H.; Sun, Y.; Zhang, R.L.; et al. The effect of influenza A (H1N1) pdm09 virus infection on cytokine production and gene expression in BV2 microglial cells. Virus Res. 2022, 312, 198716. [Google Scholar] [CrossRef]
- Li, K.; McCaw, J.M.; Cao, P. Modelling within-host macrophage dynamics in influenza virus infection. J. Theor. Biol. 2021, 508, 110492. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Xu, W.; Ai, X.; Zhu, Y.; Geng, P.; Niu, Y.; Zhu, H.; Zhou, W.; Huang, H.; Shi, X. Autophagy and Exosome Coordinately Enhance Macrophage M1 Polarization and Recruitment in Influenza A Virus Infection. Front. Immunol. 2022, 13, 722053. [Google Scholar] [CrossRef] [PubMed]
- Koutsakos, M.; Kedzierska, K.; Subbarao, K. Immune Responses to Avian Influenza Viruses. J. Immunol. 2019, 202, 382–391. [Google Scholar] [CrossRef]
- Ashkar, S.; Weber, G.F.; Panoutsakopoulou, V.; Sanchirico, M.E.; Jansson, M.; Zawaideh, S.; Rittling, S.R.; Denhardt, D.T.; Glimcher, M.J.; Cantor, H. Eta-1 (osteopontin): An early component of type-1 (cell-mediated) immunity. Science 2000, 287, 860–864. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.C.; Sacramento, C.Q.; Pereira-dutra, F.S.; Silva, P.P.; Mattos, M.; de Freitas, C.S.; de Melo, G.R.; Macedo, M.C.; Azevedo-quintanilha, I.G.; Carlos, A.S.; et al. Macrophages Undergo Necroptosis during Severe Influenza A Infection and Contribute to Virus-Associated Cytokine Storm Short Title: Macrophages Necroptosis during Influenza A Infection. bioRxiv 2022. [Google Scholar] [CrossRef]
- Xi-zhi, J.; Guoa, B.; Thomas, P.G. New fronts emerge in the influenza cytokine storm Xi-zhi. Physiol. Behav. 2017, 39, 541–550. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Characteristics | Negative | ILI | SARI | Fatal | Total |
|---|---|---|---|---|---|
| Number of samples | 65 | 78 | 76 | 22 | 241 |
| Age (years), median ± SD | 17 ± 30.9 | 28 ± 22.6 | 46 ± 23.7 | 49 ± 14.9 | 35 ± 25.7 |
| Male, n (%) | 34 (53.1) | 47 (60.3) | 35 (46.0) | 10 (45.4) | 126 (52.3) |
| Female, n (%) | 31 (46.9) | 31 (39.7) | 41 (54.0) | 12 (54.6) | 115 (47.7) |
| Symptoms, n/N (%) | |||||
| Dyspnea | 27/50 (54.0) | NA | 71/76 (93.4) | 16/16 (100) | 114/142 (80.3) |
| Respiratory distress | 23/65 (35.4) | NA | 36/76 (47.4) | 4/22 (18.2) | 59/163 (36.2) |
| Oxygen saturation < 95% | 15/65 (23.1) | NA | 28/76 (36.8) | 4/22 (18.2) | 47/163 (28.8) |
| Comorbidities, n/N (%) * | 11/28 (39.3) | 8/23 (34.8) | 24/48 (50.0) | 3/4 (75.0) | 46/103 (44.7) |
| mRNA Detection | Groups | n/N (%) | p-Value |
|---|---|---|---|
| tOPN | Negative | 12/65 (18.5) | <0.05 |
| A(H1N1)pdm09 | 60/176 (34.1) | ||
| ILI | 22/78 (28.2) | 0.14 | |
| Severe (SARI + fatal) | 38/98 (38.8) | ||
| Non-fatal (ILI + SARI) | 47/154 (30.5) | <0.01 | |
| Fatal | 13/22 (59.1) | ||
| OPNa | Negative | 2/65 (3.1) | 0.61 |
| A(H1N1)pdm09 | 8/176 (4.5) | ||
| ILI | 2/78 (2.6) | 0.26 | |
| Severe (SARI + fatal) | 6/98 (6.1) | ||
| Non-fatal (ILI + SARI) | 5/154 (3.2) | 0.03 | |
| Fatal | 5/22 (13.6) | ||
| OPNb | Negative | 0/65 (0) | NA |
| A(H1N1)pdm09 | 0/176 (0) | ||
| ILI | 0/78 (0) | NA | |
| Severe (SARI + fatal) | 0/98 (0) | ||
| Non-fatal (ILI + SARI) | 0/154 (0) | NA | |
| Fatal | 0/22 (0) | ||
| OPNc | Negative | 8/65 (12.3) | 0.84 |
| A(H1N1)pdm09 | 20/176 (11.4) | ||
| ILI | 10/78 (12.8) | 0.59 | |
| Severe (SARI + fatal) | 10/98 (10.2) | ||
| Non-fatal (ILI + SARI) | 16/154 (10.4) | 0.28 | |
| Fatal | 4/22 (18.2) | ||
| OPN4 | Negative | 43/65 (66.1) | 0.05 |
| A(H1N1)pdm09 | 138/176 (78.4) | ||
| ILI | 54/78 (69.2) | <0.01 | |
| Severe (SARI + fatal) | 84/98 (85.7) | ||
| Non-fatal (ILI + SARI) | 119/154 (77.3) | 0.33 | |
| Fatal | 19/22 (86.4) | ||
| OPN5 | Negative | 0/65 (0) | NA |
| A(H1N1)pdm09 | 0/176 (0) | ||
| ILI | 0/78 (0) | NA | |
| Severe (SARI + fatal) | 0/98 (0) | ||
| Non-fatal (ILI + SARI) | 0/154 (0) | NA | |
| Fatal | 0/22 (86.4) |
| mRNA Detection | Dyspnea | Respiratory Failure | Oxygen Saturation < 95% | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Positive | Negative | Positive | Negative | Positive | Negative | ||||
| n/N (%) | n/N (%) | p-Value | n/N (%) | n/N (%) | p-Value | n/N (%) | n/N (%) | p-Value | |
| tOPN | 32/87 (36.8) | 6/32 (18.7) | 0.04 | 4/40 (35.0) | 46/136 (33.8) | 0.79 | 9/32 (28.1) | 51/144 (35.4) | 0.41 |
| OPN4 | 75/87 (86.2) | 21/32 (65.6) | 0.01 | 36/40 (90.0) | 102/136 (75.0) | 0.02 | 30/32 (93.7) | 108/144 (75.0) | 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martins, J.S.C.d.C.; Sousa, T.d.C.; Oliveira, M.d.L.d.A.; Gimba, E.R.P.; Siqueira, M.M.; Matos, A.d.R. Total Osteopontin and Its Isoform OPN4 Are Differently Expressed in Respiratory Samples during Influenza A(H1N1)pdm09 Infection and Progression. Microorganisms 2023, 11, 1349. https://doi.org/10.3390/microorganisms11051349
Martins JSCdC, Sousa TdC, Oliveira MdLdA, Gimba ERP, Siqueira MM, Matos AdR. Total Osteopontin and Its Isoform OPN4 Are Differently Expressed in Respiratory Samples during Influenza A(H1N1)pdm09 Infection and Progression. Microorganisms. 2023; 11(5):1349. https://doi.org/10.3390/microorganisms11051349
Chicago/Turabian StyleMartins, Jéssica Santa Cruz de Carvalho, Thiago das Chagas Sousa, Maria de Lourdes de Aguiar Oliveira, Etel Rodrigues Pereira Gimba, Marilda Mendonça Siqueira, and Aline da Rocha Matos. 2023. "Total Osteopontin and Its Isoform OPN4 Are Differently Expressed in Respiratory Samples during Influenza A(H1N1)pdm09 Infection and Progression" Microorganisms 11, no. 5: 1349. https://doi.org/10.3390/microorganisms11051349
APA StyleMartins, J. S. C. d. C., Sousa, T. d. C., Oliveira, M. d. L. d. A., Gimba, E. R. P., Siqueira, M. M., & Matos, A. d. R. (2023). Total Osteopontin and Its Isoform OPN4 Are Differently Expressed in Respiratory Samples during Influenza A(H1N1)pdm09 Infection and Progression. Microorganisms, 11(5), 1349. https://doi.org/10.3390/microorganisms11051349