
Periodontal Pathogens and Their Links to Neuroinflammation and Neurodegeneration



Abstract
:1. Introduction
Gingivitis and Its Progression to Periodontitis
2. Periodontal Pathogens, Virulence Factors and Host Response
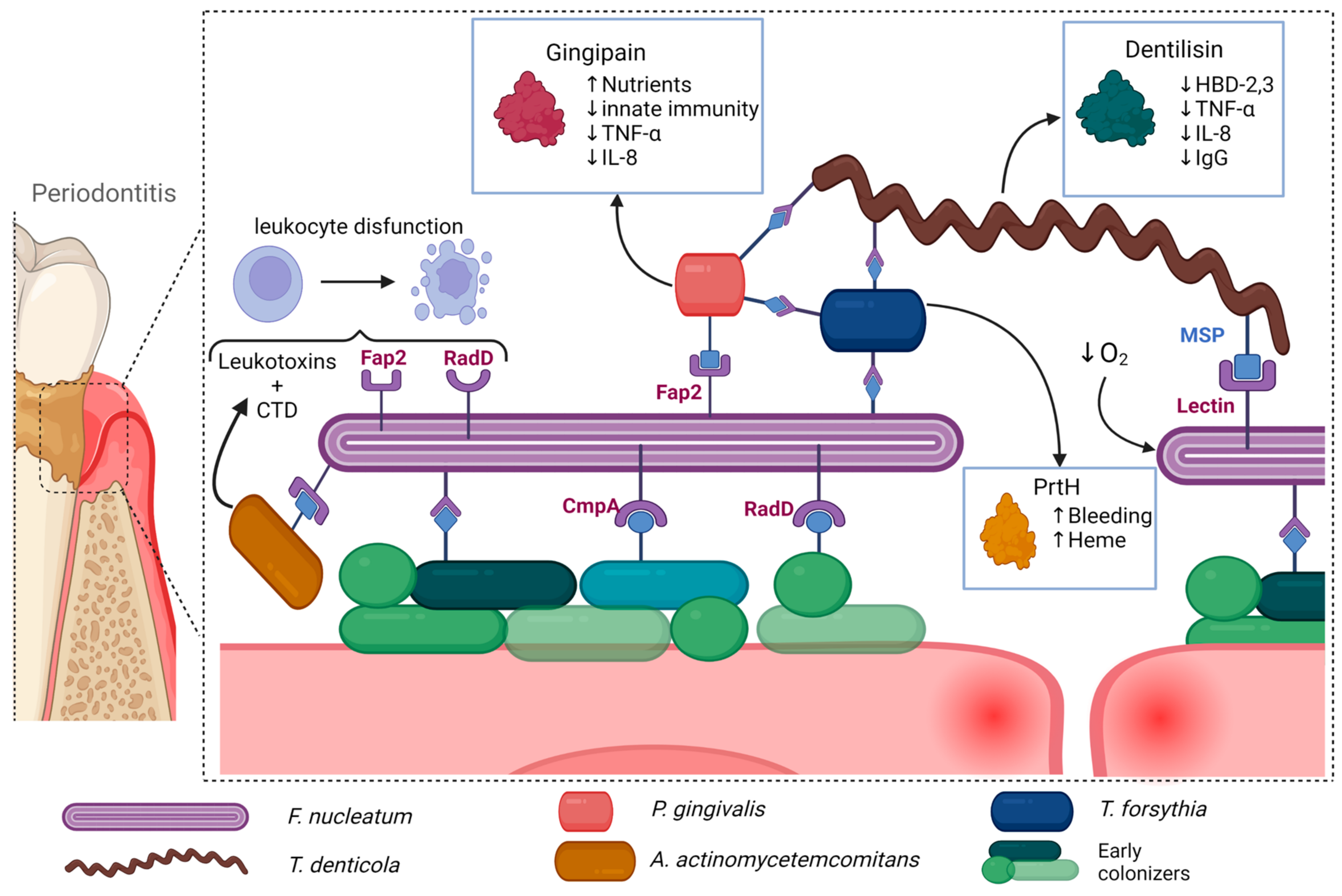
2.1. Porphyromonas gingivalis
2.2. Tannerella forsythia
2.3. Treponema denticola
2.4. Fusobacterium nucleatum
2.5. Aggregatibacter actinomycetemcomitans
2.6. Synergistic Behavior
2.7. Mechanisms of Systemic Inflammation and Its Correlation to Periodontitis
3. Neurodegenerative and Periodontal Disease
3.1. Periodontitis and Neurodegenerative Disorders
3.2. Bidirectional Interactions, Causation, and Critical Overview
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. Systemic Extraction of Data

Appendix A.2. Eligibility Criteria
References
- Raizada, M.K.; Joe, B.; Bryan, N.S.; Chang, E.B.; Dewhirst, F.E.; Borisy, G.G.; Galis, Z.S.; Henderson, W.; Jose, P.A.; Ketchum, C.J.; et al. Report of the National Heart, Lung, and Blood Institute Working Group on the Role of Microbiota in Blood Pressure Regulation: Current Status and Future Directions. Hypertension 2017, 70, 479–485. [Google Scholar] [CrossRef]
- Reynolds-Campbell, G.; Nicholson, A.; Thoms-Rodriguez, C.-A. Oral Bacterial Infections: Diagnosis and Management. Dent. Clin. N. Am. 2017, 61, 305–318. [Google Scholar] [CrossRef] [PubMed]
- Definition of Gingiva—NCI Dictionary of Cancer Terms—NCI. Available online: https://www.cancer.gov/publications/dictionaries/cancer-terms/def/gingiva (accessed on 24 November 2022).
- Rathee, M.; Jain, P. Gingivitis; StatPearls Publishing: Tampa, FL, USA, 2022. [Google Scholar]
- Nowicki, E.M.; Shroff, R.; Singleton, J.A.; Renaud, D.E.; Wallace, D.; Drury, J.; Zirnheld, J.; Colleti, B.; Ellington, A.D.; Lamont, R.J.; et al. Microbiota and Metatranscriptome Changes Accompanying the Onset of Gingivitis. mBio 2018, 9, e00575-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, R.C. Gingivitis. J. Clin. Periodontol. 1986, 13, 345–359. [Google Scholar] [CrossRef]
- Nazir, M.A. Prevalence of Periodontal Disease, Its Association with Systemic Diseases and Prevention. Int. J. Health Sci. 2017, 11, 72–80. [Google Scholar]
- Kassebaum, N.J.; Bernabé, E.; Dahiya, M.; Bhandari, B.; Murray, C.J.L.; Marcenes, W. Global Burden of Severe Periodontitis in 1990–2010. J. Dent. Res. 2014, 93, 1045–1053. [Google Scholar] [CrossRef]
- Huang, S.; Li, R.; Zeng, X.; He, T.; Zhao, H.; Chang, A.; Bo, C.; Chen, J.; Yang, F.; Knight, R.; et al. Predictive Modeling of Gingivitis Severity and Susceptibility via Oral Microbiota. ISME J. 2014, 8, 1768–1780. [Google Scholar] [CrossRef] [PubMed]
- Kinane, D.F.; Stathopoulou, P.G.; Papapanou, P.N. Periodontal Diseases. Nat. Rev. Dis. Prim. 2017, 3, 17038. [Google Scholar] [CrossRef] [PubMed]
- Abusleme, L.; Hoare, A.; Hong, B.-Y.; Diaz, P.I. Microbial Signatures of Health, Gingivitis, and Periodontitis. Periodontology 2000 2021, 86, 57–78. [Google Scholar] [CrossRef] [PubMed]
- Socransky, S.S.; Haffajee, A.D.; Cugini, M.A.; Smith, C.; Kent, R.L. Microbial Complexes in Subgingival Plaque. J. Clin. Periodontol. 1998, 25, 134–144. [Google Scholar] [CrossRef]
- Guthmiller, J.M.; Novak, K.F. Periodontal Diseases; ASM Press: Washington, DC, USA, 2002. [Google Scholar]
- Suzuki, N.; Yoneda, M.; Hirofuji, T. Mixed Red-Complex Bacterial Infection in Periodontitis. Int. J. Dent. 2013, 2013, 587279. [Google Scholar] [CrossRef]
- How, K.Y.; Song, K.P.; Chan, K.G. Porphyromonas Gingivalis: An Overview of Periodontopathic Pathogen below the Gum Line. Front. Microbiol. 2016, 7, 53. [Google Scholar] [CrossRef] [Green Version]
- Dominy, S.S.; Lynch, C.; Ermini, F.; Benedyk, M.; Marczyk, A.; Konradi, A.; Nguyen, M.; Haditsch, U.; Raha, D.; Griffin, C.; et al. Porphyromonas Gingivalis in Alzheimer’s Disease Brains: Evidence for Disease Causation and Treatment with Small-Molecule Inhibitors. Sci. Adv. 2019, 5, eaau3333. [Google Scholar] [CrossRef] [Green Version]
- Okamura, H.; Hirota, K.; Yoshida, K.; Weng, Y.; He, Y.; Shiotsu, N.; Ikegame, M.; Uchida-Fukuhara, Y.; Tanai, A.; Guo, J. Outer Membrane Vesicles of Porphyromonas Gingivalis: Novel Communication Tool and Strategy. Jpn. Dent. Sci. Rev. 2021, 57, 138–146. [Google Scholar] [CrossRef]
- Veith, P.D.; Chen, Y.-Y.; Gorasia, D.G.; Chen, D.; Glew, M.D.; O’Brien-Simpson, N.M.; Cecil, J.D.; Holden, J.A.; Reynolds, E.C. Porphyromonas Gingivalis Outer Membrane Vesicles Exclusively Contain Outer Membrane and Periplasmic Proteins and Carry a Cargo Enriched with Virulence Factors. J. Proteome Res. 2014, 13, 2420–2432. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Collyer, C.A. Gingipains from Porphyromonas Gingivalis—Complex Domain Structures Confer Diverse Functions. Eur. J. Microbiol. Immunol. (Bp) 2011, 1, 41–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, L.; Han, N.; Du, J.; Guo, L.; Luo, Z.; Liu, Y. Pathogenesis of Important Virulence Factors of Porphyromonas Gingivalis via Toll-Like Receptors. Front. Cell. Infect. Microbiol. 2019, 9, 262. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Chen, L.; Li, S.; Gu, Z.; Yan, J. Lipopolysaccharide (LPS) of Porphyromonas Gingivalis Induces IL-1beta, TNF-Alpha and IL-6 Production by THP-1 Cells in a Way Different from That of Escherichia Coli LPS. Innate Immun. 2008, 14, 99–107. [Google Scholar] [CrossRef]
- Zhang, J.; Yu, C.; Zhang, X.; Chen, H.; Dong, J.; Lu, W.; Song, Z.; Zhou, W. Porphyromonas Gingivalis Lipopolysaccharide Induces Cognitive Dysfunction, Mediated by Neuronal Inflammation via Activation of the TLR4 Signaling Pathway in C57BL/6 Mice. J. Neuroinflamm. 2018, 15, 37. [Google Scholar] [CrossRef]
- Groeger, S.; Jarzina, F.; Domann, E.; Meyle, J. Porphyromonas Gingivalis Activates NFκB and MAPK Pathways in Human Oral Epithelial Cells. BMC Immunol. 2017, 18, 1. [Google Scholar] [CrossRef] [Green Version]
- Hienz, S.A.; Paliwal, S.; Ivanovski, S. Mechanisms of Bone Resorption in Periodontitis. J. Immunol. Res. 2015, 2015, 615486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, Z.; Yoshizawa-Smith, S.; Glowacki, A.; Maltos, K.; Pacheco, C.; Shehabeldin, M.; Mulkeen, M.; Myers, N.; Chong, R.; Verdelis, K.; et al. Induction of M2 Macrophages Prevents Bone Loss in Murine Periodontitis Models. J. Dent. Res. 2019, 98, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Geatch, D.R.; Harris, J.I.; Heasman, P.A.; Taylor, J.J. In Vitro Studies of Lymphocyte Apoptosis Induced by the Periodontal Pathogen Porphyromonas Gingivalis. J. Periodontal Res. 1999, 34, 70–78. [Google Scholar] [CrossRef]
- Jotwani, R.; Cutler, C.W. Fimbriated Porphyromonas Gingivalis Is More Efficient than Fimbria-Deficient P. gingivalis in Entering Human Dendritic Cells in Vitro and Induces an Inflammatory Th1 Effector Response. Infect. Immun. 2004, 72, 1725–1732. [Google Scholar] [CrossRef] [Green Version]
- Werheim, E.R.; Senior, K.G.; Shaffer, C.A.; Cuadra, G.A. Oral Pathogen Porphyromonas Gingivalis Can Escape Phagocytosis of Mammalian Macrophages. Microorganisms 2020, 8, 1432. [Google Scholar] [CrossRef]
- Vincents, B.; Guentsch, A.; Kostolowska, D.; von Pawel-Rammingen, U.; Eick, S.; Potempa, J.; Abrahamson, M. Cleavage of IgG1 and IgG3 by Gingipain K from Porphyromonas Gingivalis May Compromise Host Defense in Progressive Periodontitis. FASEB J. 2011, 25, 3741–3750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorn, B.R.; Dunn, W.A.; Progulske-Fox, A. Porphyromonas Gingivalis Traffics to Autophagosomes in Human Coronary Artery Endothelial Cells. Infect. Immun. 2001, 69, 5698–5708. [Google Scholar] [CrossRef] [Green Version]
- Porphyromonas Gingivalis (ID 714)—Genome—NCBI. Available online: https://www.ncbi.nlm.nih.gov/genome/714 (accessed on 7 April 2023).
- Mendez, K.N.; Hoare, A.; Soto, C.; Bugueño, I.; Olivera, M.; Meneses, C.; Pérez-Donoso, J.M.; Castro-Nallar, E.; Bravo, D. Variability in Genomic and Virulent Properties of Porphyromonas Gingivalis Strains Isolated From Healthy and Severe Chronic Periodontitis Individuals. Front. Cell. Infect. Microbiol. 2019, 9, 246. [Google Scholar] [CrossRef]
- Rocha, F.G.; Berges, A.; Sedra, A.; Ghods, S.; Kapoor, N.; Pill, L.; Davey, M.E.; Fairman, J.; Gibson, F.C. A Porphyromonas Gingivalis Capsule-Conjugate Vaccine Protects From Experimental Oral Bone Loss. Front. Oral Health 2021, 2, 686402. [Google Scholar] [CrossRef]
- Dahlén, G.; Gmür, R.; Yoshino, T. Phenotypes, Serotypes and Antibiotic Susceptibility of Swedish Porphyromonas Gingivalis Isolates from Periodontitis and Periodontal Abscesses. Oral Microbiol. Immunol. 2007, 22, 80–86. [Google Scholar] [CrossRef]
- Naito, M.; Hirakawa, H.; Yamashita, A.; Ohara, N.; Shoji, M.; Yukitake, H.; Nakayama, K.; Toh, H.; Yoshimura, F.; Kuhara, S.; et al. Determination of the Genome Sequence of Porphyromonas Gingivalis Strain ATCC 33277 and Genomic Comparison with Strain W83 Revealed Extensive Genome Rearrangements in P. gingivalis. DNA Res. 2008, 15, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Biyikoğlu, B.; Ricker, A.; Diaz, P.I. Strain-Specific Colonization Patterns and Serum Modulation of Multi-Species Oral Biofilm Development. Anaerobe 2012, 18, 459–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davey, M.E.; Duncan, M.J. Enhanced Biofilm Formation and Loss of Capsule Synthesis: Deletion of a Putative Glycosyltransferase in Porphyromonas Gingivalis. J. Bacteriol. 2006, 188, 5510–5523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seers, C.A.; Mahmud, A.S.M.; Huq, N.L.; Cross, K.J.; Reynolds, E.C. Porphyromonas Gingivalis Laboratory Strains and Clinical Isolates Exhibit Different Distribution of Cell Surface and Secreted Gingipains. J. Oral Microbiol. 2020, 13, 1858001. [Google Scholar] [CrossRef]
- Chastain-Gross, R.P.; Xie, G.; Bélanger, M.; Kumar, D.; Whitlock, J.A.; Liu, L.; Raines, S.M.; Farmerie, W.G.; Daligault, H.E.; Han, C.S.; et al. Genome Sequence of Porphyromonas Gingivalis Strain 381. Genome Announc. 2017, 5, e01467-16. [Google Scholar] [CrossRef] [Green Version]
- Aduse-Opoku, J.; Joseph, S.; Devine, D.A.; Marsh, P.D.; Curtis, M.A. Molecular Basis for Avirulence of Spontaneous Variants of Porphyromonas Gingivalis: Genomic Analysis of Strains W50, BE1 and BR1. Mol. Oral Microbiol. 2022, 37, 122–132. [Google Scholar] [CrossRef]
- Sharma, A. Virulence Mechanisms of Tannerella Forsythia. Periodontol 2000 2010, 54, 106–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malinowski, B.; Węsierska, A.; Zalewska, K.; Sokołowska, M.M.; Bursiewicz, W.; Socha, M.; Ozorowski, M.; Pawlak-Osińska, K.; Wiciński, M. The Role of Tannerella Forsythia and Porphyromonas Gingivalis in Pathogenesis of Esophageal Cancer. Infect. Agents Cancer 2019, 14, 3. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, T.; Tomi, N.; Fukuyo, Y.; Ishikura, H.; Ohno, Y.; Arvind, R.; Arai, T.; Ishikawa, I.; Arakawa, S. Isolation and Identification of a Cytopathic Activity in Tannerella Forsythia. Biochem. Biophys. Res. Commun. 2006, 351, 133–139. [Google Scholar] [CrossRef]
- Tomi, N.; Fukuyo, Y.; Arakawa, S.; Nakajima, T. Pro-Inflammatory Cytokine Production from Normal Human Fibroblasts Is Induced by Tannerella Forsythia Detaching Factor. J. Periodontal Res. 2008, 43, 136–142. [Google Scholar] [CrossRef]
- Onishi, S.; Honma, K.; Liang, S.; Stathopoulou, P.; Kinane, D.; Hajishengallis, G.; Sharma, A. Toll-Like Receptor 2-Mediated Interleukin-8 Expression in Gingival Epithelial Cells by the Tannerella Forsythia Leucine-Rich Repeat Protein BspA. Infect. Immun. 2008, 76, 198–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.R.; Jun, H.K.; Choi, B.K. Tannerella Forsythia BspA Increases the Risk Factors for Atherosclerosis in ApoE(-/-) Mice. Oral Dis. 2014, 20, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Settem, R.P.; Honma, K.; Shankar, M.; Li, M.; LaMonte, M.; Xu, D.; Genco, R.J.; Browne, R.W.; Sharma, A. Tannerella Forsythia-Produced Methylglyoxal Causes Accumulation of Advanced Glycation Endproducts to Trigger Cytokine Secretion in Human Monocytes. Mol. Oral Microbiol. 2018, 33, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.-J.; Choi, Y.-J.; An, S.-J.; Lee, H.-R.; Jun, H.-K.; Choi, B.-K. Tannerella Forsythia GroEL Induces Inflammatory Bone Resorption and Synergizes with Interleukin-17. Mol. Oral Microbiol. 2017, 32, 301–313. [Google Scholar] [CrossRef]
- Bryzek, D.; Ksiazek, M.; Bielecka, E.; Karim, A.Y.; Potempa, B.; Staniec, D.; Koziel, J.; Potempa, J. A Pathogenic Trace of Tannerella Forsythia—Shedding of Soluble Fully Active Tumor Necrosis Factor α from the Macrophage Surface by Karilysin. Mol. Oral Microbiol. 2014, 29, 294–306. [Google Scholar] [CrossRef] [Green Version]
- Karim, A.Y.; Kulczycka, M.; Kantyka, T.; Dubin, G.; Jabaiah, A.; Daugherty, P.S.; Thogersen, I.B.; Enghild, J.J.; Nguyen, K.-A.; Potempa, J. A Novel Matrix Metalloprotease-like Enzyme (Karilysin) of the Periodontal Pathogen Tannerella Forsythia ATCC 43037. Biol. Chem. 2010, 391, 105–117. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.J.; Sekot, G.; Duman, M.; Chtcheglova, L.; Messner, P.; Peterlik, H.; Schäffer, C.; Hinterdorfer, P. Characterizing the S-Layer Structure and Anti-S-Layer Antibody Recognition on Intact Tannerella Forsythia Cells by Scanning Probe Microscopy and Small Angle X-Ray Scattering. J. Mol. Recognit. 2013, 26, 542–549. [Google Scholar] [CrossRef] [Green Version]
- Sekot, G.; Posch, G.; Messner, P.; Matejka, M.; Rausch-Fan, X.; Andrukhov, O.; Schäffer, C. Potential of the Tannerella Forsythia S-Layer to Delay the Immune Response. J. Dent. Res. 2011, 90, 109–114. [Google Scholar] [CrossRef] [Green Version]
- Shimotahira, N.; Oogai, Y.; Kawada-Matsuo, M.; Yamada, S.; Fukutsuji, K.; Nagano, K.; Yoshimura, F.; Noguchi, K.; Komatsuzawa, H. The Surface Layer of Tannerella Forsythia Contributes to Serum Resistance and Oral Bacterial Coaggregation. Infect. Immun. 2013, 81, 1198–1206. [Google Scholar] [CrossRef] [Green Version]
- Bloch, S.; Zwicker, S.; Bostanci, N.; Sjöling, Å.; Boström, E.A.; Belibasakis, G.N.; Schäffer, C. Immune Response Profiling of Primary Monocytes and Oral Keratinocytes to Different Tannerella Forsythia Strains and Their Cell Surface Mutants. Mol. Oral Microbiol. 2018, 33, 155–167. [Google Scholar] [CrossRef]
- Jusko, M.; Potempa, J.; Karim, A.Y.; Ksiazek, M.; Riesbeck, K.; Garred, P.; Eick, S.; Blom, A.M. A Metalloproteinase Karilysin Present in the Majority of Tannerella Forsythia Isolates Inhibits All Pathways of the Complement System. J. Immunol. 2012, 188, 2338–2349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloch, S.; Thurnheer, T.; Murakami, Y.; Belibasakis, G.N.; Schäffer, C. Behavior of Two Tannerella Forsythia Strains and Their Cell Surface Mutants in Multispecies Oral Biofilms. Mol. Oral Microbiol. 2017, 32, 404–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedrich, V.; Janesch, B.; Windwarder, M.; Maresch, D.; Braun, M.L.; Megson, Z.A.; Vinogradov, E.; Goneau, M.-F.; Sharma, A.; Altmann, F.; et al. Tannerella Forsythia Strains Display Different Cell-Surface Nonulosonic Acids: Biosynthetic Pathway Characterization and First Insight into Biological Implications. Glycobiology 2017, 27, 342–357. [Google Scholar] [CrossRef] [Green Version]
- Chinthamani, S.; Settem, R.P.; Honma, K.; Stafford, G.P.; Sharma, A. Tannerella Forsythia Strains Differentially Induce Interferon Gamma-Induced Protein 10 (IP-10) Expression in Macrophages Due to Lipopolysaccharide Heterogeneity. Pathog. Dis. 2022, 80, ftac008. [Google Scholar] [CrossRef] [PubMed]
- Ruby, J.; Martin, M.; Passineau, M.J.; Godovikova, V.; Fenno, J.C.; Wu, H. Activation of the Innate Immune System by Treponema Denticola Periplasmic Flagella through Toll-Like Receptor 2. Infect. Immun. 2017, 86, e00573-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaibani, P.; Caroli, F.; Nucci, C.; Sambri, V. Major Surface Protein Complex of Treponema Denticola Induces the Production of Tumor Necrosis Factor Alpha, Interleukin-1 Beta, Interleukin-6 and Matrix Metalloproteinase 9 by Primary Human Peripheral Blood Monocytes. J. Periodontal Res. 2010, 45, 361–366. [Google Scholar] [CrossRef]
- Fenno, J.C.; Hannam, P.M.; Leung, W.K.; Tamura, M.; Uitto, V.-J.; McBride, B.C. Cytopathic Effects of the Major Surface Protein and the Chymotrypsinlike Protease of Treponema Denticola. Infect. Immun. 1998, 66, 1869–1877. [Google Scholar] [CrossRef]
- Nussbaum, G.; Ben-Adi, S.; Genzler, T.; Sela, M.; Rosen, G. Involvement of Toll-like Receptors 2 and 4 in the Innate Immune Response to Treponema Denticola and Its Outer Sheath Components. Infect. Immun. 2009, 77, 3939–3947. [Google Scholar] [CrossRef] [Green Version]
- Grenier, D. Binding Properties of Treponema Denticola Lipooligosaccharide. J. Oral Microbiol. 2013, 5, 21517. [Google Scholar] [CrossRef]
- Ganther, S.; Radaic, A.; Malone, E.; Kamarajan, P.; Chang, N.-Y.N.; Tafolla, C.; Zhan, L.; Fenno, J.C.; Kapila, Y.L. Treponema Denticola Dentilisin Triggered TLR2/MyD88 Activation Upregulates a Tissue Destructive Program Involving MMPs via Sp1 in Human Oral Cells. PLoS Pathog. 2021, 17, e1009311. [Google Scholar] [CrossRef]
- Yamazaki, T.; Miyamoto, M.; Yamada, S.; Okuda, K.; Ishihara, K. Surface Protease of Treponema Denticola Hydrolyzes C3 and Influences Function of Polymorphonuclear Leukocytes. Microbes Infect. 2006, 8, 1758–1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokubu, E.; Kikuchi, Y.; Okamoto-Shibayama, K.; Nakamura, S.; Ishihara, K. Crawling Motility of Treponema Denticola Modulated by Outer Sheath Protein. Microbiol. Immunol. 2021, 65, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Nixon, C.S.; Steffen, M.J.; Ebersole, J.L. Cytokine Responses to Treponema Pectinovorum and Treponema Denticola in Human Gingival Fibroblasts. Infect. Immun. 2000, 68, 5284–5292. [Google Scholar] [CrossRef] [Green Version]
- Brissette, C.A.; Pham, T.-T.T.; Coats, S.R.; Darveau, R.P.; Lukehart, S.A. Treponema Denticola Does Not Induce Production of Common Innate Immune Mediators from Primary Gingival Epithelial Cells. Oral Microbiol. Immunol. 2008, 23, 474–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.E.; Choi, Y. Treponema Denticola Suppresses Expression of Human Beta-Defensin-2 in Gingival Epithelial Cells through Inhibition of TNFalpha Production and TLR2 Activation. Mol. Cells 2010, 29, 407–412. [Google Scholar] [CrossRef]
- Shin, J.E.; Kim, Y.S.; Oh, J.-E.; Min, B.-M.; Choi, Y. Treponema Denticola Suppresses Expression of Human {beta}-Defensin-3 in Gingival Epithelial Cells through Inhibition of the Toll-like Receptor 2 Axis. Infect. Immun. 2010, 78, 672–679. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, M.; Ishihara, K.; Okuda, K. The Treponema Denticola Surface Protease Dentilisin Degrades Interleukin-1β (IL-1β), IL-6, and Tumor Necrosis Factor Alpha. Infect. Immun. 2006, 74, 2462–2467. [Google Scholar] [CrossRef] [Green Version]
- Okuda, T.; Kimizuka, R.; Miyamoto, M.; Kato, T.; Yamada, S.; Okuda, K.; Ishihara, K. Treponema Denticola Induces Interleukin-8 and Macrophage Chemoattractant Protein 1 Production in Human Umbilical Vein Epithelial Cells. Microbes Infect. 2007, 9, 907–913. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, P.B.D.R.E.; De Lima, P.M.N.; Palma, A.L.D.R.; Abu Hasna, A.; Rossoni, R.D.; Junqueira, J.C.; De Oliveira, L.D. Review- The Periodontal Pathogen Treponema Denticola: An Atherosclerosis Risk Factor. Res. Soc. Dev. 2021, 10, e25810111637. [Google Scholar] [CrossRef]
- Han, Y.W. Fusobacterium Nucleatum: A Commensal-Turned Pathogen. Curr. Opin. Microbiol. 2015, 23, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Fan, L.; Lin, Y.; Shen, W.; Qi, Y.; Zhang, Y.; Chen, Z.; Wang, L.; Long, Y.; Hou, T.; et al. Fusobacterium Nucleatum Promotes Colorectal Cancer Metastasis through MiR-1322/CCL20 Axis and M2 Polarization. Gut Microbes 2021, 13, 1980347. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Guo, F.; Yu, Y.; Sun, T.; Ma, D.; Han, J.; Qian, Y.; Kryczek, I.; Sun, D.; Nagarsheth, N.; et al. Fusobacterium Nucleatum Promotes Chemoresistance to Colorectal Cancer by Modulating Autophagy. Cell 2017, 170, 548–563.e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, W.; Ji, X.; Zhang, X.; Tang, D.; Feng, Q. Persistent Exposure to Fusobacterium Nucleatum Triggers Chemokine/Cytokine Release and Inhibits the Proliferation and Osteogenic Differentiation Capabilities of Human Gingiva-Derived Mesenchymal Stem Cells. Front. Cell. Infect. Microbiol. 2019, 9, 429. [Google Scholar] [CrossRef]
- Fusobacterium Nucleatum Facilitates Apoptosis, ROS Generation, and Inflammatory Cytokine Production by Activating AKT/MAPK and NF- κ B Signaling Pathways in Human Gingival Fibroblasts—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/31737164/ (accessed on 2 March 2023).
- Yin, L.; Li, X.; Hou, J. Macrophages in Periodontitis: A Dynamic Shift between Tissue Destruction and Repair. Jpn. Dent. Sci. Rev. 2022, 58, 336–347. [Google Scholar] [CrossRef]
- Chen, Y.; Huang, Z.; Tang, Z.; Huang, Y.; Huang, M.; Liu, H.; Ziebolz, D.; Schmalz, G.; Jia, B.; Zhao, J. More Than Just a Periodontal Pathogen—The Research Progress on Fusobacterium Nucleatum. Front. Cell. Infect. Microbiol. 2022, 12, 64. [Google Scholar] [CrossRef] [PubMed]
- Bachrach, G.; Rosen, G.; Bellalou, M.; Naor, R.; Sela, M.N. Identification of a Fusobacterium Nucleatum 65 KDa Serine Protease. Oral Microbiol. Immunol. 2004, 19, 155–159. [Google Scholar] [CrossRef]
- Despins, C.A.; Brown, S.D.; Robinson, A.V.; Mungall, A.J.; Allen-Vercoe, E.; Holt, R.A. Modulation of the Host Cell Transcriptome and Epigenome by Fusobacterium Nucleatum. mBio 2021, 12, e0206221. [Google Scholar] [CrossRef]
- Razghonova, Y.; Zymovets, V.; Wadelius, P.; Rakhimova, O.; Manoharan, L.; Brundin, M.; Kelk, P.; Romani Vestman, N. Transcriptome Analysis Reveals Modulation of Human Stem Cells from the Apical Papilla by Species Associated with Dental Root Canal Infection. Int. J. Mol. Sci. 2022, 23, 14420. [Google Scholar] [CrossRef]
- Ang, M.Y.; Dutta, A.; Wee, W.Y.; Dymock, D.; Paterson, I.C.; Choo, S.W. Comparative Genome Analysis of Fusobacterium Nucleatum. Genome Biol. Evol. 2016, 8, 2928–2938. [Google Scholar] [CrossRef] [Green Version]
- Muchova, M.; Balacco, D.L.; Grant, M.M.; Chapple, I.L.C.; Kuehne, S.A.; Hirschfeld, J. Fusobacterium Nucleatum Subspecies Differ in Biofilm Forming Ability in Vitro. Front. Oral Health 2022, 3, 853618. [Google Scholar] [CrossRef]
- Belibasakis, G.N.; Maula, T.; Bao, K.; Lindholm, M.; Bostanci, N.; Oscarsson, J.; Ihalin, R.; Johansson, A. Virulence and Pathogenicity Properties of Aggregatibacter Actinomycetemcomitans. Pathogens 2019, 8, 222. [Google Scholar] [CrossRef] [Green Version]
- Naqvi, A.R.; Fordham, J.B.; Khan, A.; Nares, S. MicroRNAs Responsive to Aggregatibacter Actinomycetemcomitans and Porphyromonas Gingivalis LPS Modulate Expression of Genes Regulating Innate Immunity in Human Macrophages. Innate Immun. 2014, 20, 540–551. [Google Scholar] [CrossRef] [Green Version]
- Ohguchi, Y.; Ishihara, Y.; Ohguchi, M.; Koide, M.; Shirozu, N.; Naganawa, T.; Nishihara, T.; Noguchi, T. Capsular Polysaccharide from Actinobacillus Actinomycetemcomitans Inhibits IL-6 and IL-8 Production in Human Gingival Fibroblast. J. Periodontal Res. 2003, 38, 191–197. [Google Scholar] [CrossRef]
- Vernal, R.; Leon, R.; Herrera, D.; Garcia-Sanz, J.A.; Silva, A.; Sanz, M. Variability in the Response of Human Dendritic Cells Stimulated with Porphyromonas Gingivalis or Aggregatibacter Actinomycetemcomitans. J. Periodontal Res. 2008, 43, 689–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guentsch, A.; Puklo, M.; Preshaw, P.M.; Glockmann, E.; Pfister, W.; Potempa, J.; Eick, S. Neutrophils in Chronic and Aggressive Periodontitis in Interaction with Porphyromonas Gingivalis and Aggregatibacter Actinomycetemcomitans. J. Periodontal Res. 2009, 44, 368–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, A. Aggregatibacter Actinomycetemcomitans Leukotoxin: A Powerful Tool with Capacity to Cause Imbalance in the Host Inflammatory Response. Toxins 2011, 3, 242–259. [Google Scholar] [CrossRef] [Green Version]
- Pons, B.J.; Vignard, J.; Mirey, G. Cytolethal Distending Toxin Subunit B: A Review of Structure–Function Relationship. Toxins 2019, 11, 595. [Google Scholar] [CrossRef] [Green Version]
- Guerra, L.; Cortes-Bratti, X.; Guidi, R.; Frisan, T. The Biology of the Cytolethal Distending Toxins. Toxins 2011, 3, 172–190. [Google Scholar] [CrossRef] [Green Version]
- Akrivopoulou, C.; Green, I.M.; Donos, N.; Nair, S.P.; Ready, D. Aggregatibacter Actinomycetemcomitans Serotype Prevalence and Antibiotic Resistance in a UK Population with Periodontitis. J. Glob. Antimicrob. Resist. 2017, 10, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Åberg, C.H.; Kelk, P.; Johansson, A. Aggregatibacter Actinomycetemcomitans: Virulence of Its Leukotoxin and Association with Aggressive Periodontitis. Virulence 2014, 6, 188–195. [Google Scholar] [CrossRef] [Green Version]
- Brígido, J.A.; da Silveira, V.R.S.; Rego, R.O.; Nogueira, N.A.P. Serotypes of Aggregatibacter Actinomycetemcomitans in Relation to Periodontal Status and Geographic Origin of Individuals-a Review of the Literature. Med. Oral Patol. Oral Cir. Bucal 2014, 19, e184–e191. [Google Scholar] [CrossRef]
- Lin, F.-Y.; Hsiao, F.-P.; Huang, C.-Y.; Shih, C.-M.; Tsao, N.-W.; Tsai, C.-S.; Yang, S.-F.; Chang, N.-C.; Hung, S.-L.; Lin, Y.-W. Porphyromonas Gingivalis GroEL Induces Osteoclastogenesis of Periodontal Ligament Cells and Enhances Alveolar Bone Resorption in Rats. PLoS ONE 2014, 9, e102450. [Google Scholar] [CrossRef]
- Song, H.; Bélanger, M.; Whitlock, J.; Kozarov, E.; Progulske-Fox, A. Hemagglutinin B Is Involved in the Adherence of Porphyromonas Gingivalis to Human Coronary Artery Endothelial Cells. Infect. Immun. 2005, 73, 7267–7273. [Google Scholar] [CrossRef] [Green Version]
- Posch, G.; Andrukhov, O.; Vinogradov, E.; Lindner, B.; Messner, P.; Holst, O.; Schäffer, C. Structure and Immunogenicity of the Rough-Type Lipopolysaccharide from the Periodontal Pathogen Tannerella Forsythia. Clin. Vaccine Immunol. 2013, 20, 945–953. [Google Scholar] [CrossRef] [Green Version]
- Ikegami, A.; Honma, K.; Sharma, A.; Kuramitsu, H.K. Multiple Functions of the Leucine-Rich Repeat Protein LrrA of Treponema Denticola. Infect. Immun. 2004, 72, 4619–4627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nørskov-Lauritsen, N.; Claesson, R.; Jensen, A.B.; Åberg, C.H.; Haubek, D. Aggregatibacter Actinomycetemcomitans: Clinical Significance of a Pathobiont Subjected to Ample Changes in Classification and Nomenclature. Pathogens 2019, 8, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berezow, A.B.; Darveau, R.P. Microbial Shift and Periodontitis. Periodontol 2000 2011, 55, 36–47. [Google Scholar] [CrossRef]
- Holt, S.C.; Bramanti, T.E. Factors in Virulence Expression and Their Role in Periodontal Disease Pathogenesis. Crit. Rev. Oral Biol. Med. 1991, 2, 177–281. [Google Scholar] [CrossRef]
- Rosen, G.; Genzler, T.; Sela, M.N. Coaggregation of Treponema Denticola with Porphyromonas Gingivalis and Fusobacterium Nucleatum Is Mediated by the Major Outer Sheath Protein of Treponema Denticola. FEMS Microbiol. Lett. 2008, 289, 59–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inagaki, S.; Onishi, S.; Kuramitsu, H.K.; Sharma, A. Porphyromonas Gingivalis Vesicles Enhance Attachment, and the Leucine-Rich Repeat BspA Protein Is Required for Invasion of Epithelial Cells by “Tannerella Forsythia”. Infect. Immun. 2006, 74, 5023–5028. [Google Scholar] [CrossRef] [Green Version]
- Kin, L.X.; Butler, C.A.; Slakeski, N.; Hoffmann, B.; Dashper, S.G.; Reynolds, E.C. Metabolic Cooperativity between Porphyromonas Gingivalis and Treponema Denticola. J. Oral Microbiol. 2020, 12, 1808750. [Google Scholar] [CrossRef]
- Tan, K.H.; Seers, C.A.; Dashper, S.G.; Mitchell, H.L.; Pyke, J.S.; Meuric, V.; Slakeski, N.; Cleal, S.M.; Chambers, J.L.; McConville, M.J.; et al. Porphyromonas Gingivalis and Treponema Denticola Exhibit Metabolic Symbioses. PLoS Pathog. 2014, 10, e1003955. [Google Scholar] [CrossRef]
- Saito, Y.; Fujii, R.; Nakagawa, K.-I.; Kuramitsu, H.K.; Okuda, K.; Ishihara, K. Stimulation of Fusobacterium Nucleatum Biofilm Formation by Porphyromonas Gingivalis. Oral Microbiol. Immunol. 2008, 23, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, D.P.; Fitzsimonds, Z.R.; Lamont, R.J. Metabolic Signaling and Spatial Interactions in the Oral Polymicrobial Community. J. Dent. Res. 2019, 98, 1308–1314. [Google Scholar] [CrossRef] [PubMed]
- Saito, A.; Inagaki, S.; Kimizuka, R.; Okuda, K.; Hosaka, Y.; Nakagawa, T.; Ishihara, K. Fusobacterium Nucleatum Enhances Invasion of Human Gingival Epithelial and Aortic Endothelial Cells by Porphyromonas Gingivalis. FEMS Immunol. Med. Microbiol. 2008, 54, 349–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoneda, M.; Yoshikane, T.; Motooka, N.; Yamada, K.; Hisama, K.; Naito, T.; Okada, I.; Yoshinaga, M.; Hidaka, K.; Imaizumi, K.; et al. Stimulation of Growth of Porphyromonas Gingivalis by Cell Extracts from Tannerella Forsythia. J. Periodontal Res. 2005, 40, 105–109. [Google Scholar] [CrossRef]
- Chen, P.B.; Davern, L.B.; Katz, J.; Eldridge, J.H.; Michalek, S.M. Host Responses Induced by Co-Infection with Porphyromonas Gingivalis and Actinobacillus Actinomycetemcomitans in a Murine Model. Oral Microbiol. Immunol. 1996, 11, 274–281. [Google Scholar] [CrossRef]
- ter Steeg, P.F.; van der Hoeven, J.S. Growth Stimulation of Treponema Denticola by Periodontal Microorganisms. Antonie Van Leeuwenhoek 1990, 57, 63–70. [Google Scholar] [CrossRef]
- Sharma, A.; Inagaki, S.; Sigurdson, W.; Kuramitsu, H.K. Synergy between Tannerella Forsythia and Fusobacterium Nucleatum in Biofilm Formation. Oral Microbiol. Immunol. 2005, 20, 39–42. [Google Scholar] [CrossRef]
- Honma, K.; Ruscitto, A.; Sharma, A. β-Glucanase Activity of the Oral Bacterium Tannerella Forsythia Contributes to the Growth of a Partner Species, Fusobacterium Nucleatum, in Cobiofilms. Appl. Environ. Microbiol. 2017, 84, e01759-17. [Google Scholar] [CrossRef] [Green Version]
- Ruscitto, A.; Honma, K.; Veeramachineni, V.M.; Nishikawa, K.; Stafford, G.P.; Sharma, A. Regulation and Molecular Basis of Environmental Muropeptide Uptake and Utilization in Fastidious Oral Anaerobe Tannerella Forsythia. Front. Microbiol. 2017, 8, 648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Settem, R.P.; El-Hassan, A.T.; Honma, K.; Stafford, G.P.; Sharma, A. Fusobacterium Nucleatum and Tannerella Forsythia Induce Synergistic Alveolar Bone Loss in a Mouse Periodontitis Model. Infect. Immun. 2012, 80, 2436–2443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, D.; Meng, H.; Xu, L.; Zhang, L.; Chen, Z.; Feng, X.; Lu, R.; Sun, X.; Ren, X. Systemic Inflammation Markers in Patients With Aggressive Periodontitis: A Pilot Study. J. Periodontol. 2008, 79, 2340–2346. [Google Scholar] [CrossRef]
- D’Aiuto, F.; Parkar, M.; Andreou, G.; Suvan, J.; Brett, P.M.; Ready, D.; Tonetti, M.S. Periodontitis and Systemic Inflammation: Control of the Local Infection Is Associated with a Reduction in Serum Inflammatory Markers. J. Dent. Res. 2004, 83, 156–160. [Google Scholar] [CrossRef]
- El-Shinnawi, U.; Soory, M. Associations between Periodontitis and Systemic Inflammatory Diseases: Response to Treatment. Recent Pat. Endocr. Metab. Immune Drug Discov. 2013, 7, 169–188. [Google Scholar] [CrossRef]
- Dhotre, S.V.; Davane, M.S.; Nagoba, B.S. Periodontitis, Bacteremia and Infective Endocarditis: A Review Study. Arch. Pediatr. Infect. Dis. 2017, 5, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Martínez-García, M.; Hernández-Lemus, E. Periodontal Inflammation and Systemic Diseases: An Overview. Front. Physiol. 2021, 12, 709438. [Google Scholar] [CrossRef]
- Pisani, F.; Pisani, V.; Arcangeli, F.; Harding, A.; Singhrao, S.K. The Mechanistic Pathways of Periodontal Pathogens Entering the Brain: The Potential Role of Treponema Denticola in Tracing Alzheimer’s Disease Pathology. Int. J. Environ. Res. Public Health 2022, 19, 9386. [Google Scholar] [CrossRef]
- Local and Systemic Mechanisms Linking Periodontal Disease and Inflammatory Comorbidities|Nature Reviews Immunology. Available online: https://www.nature.com/articles/s41577-020-00488-6 (accessed on 6 March 2023).
- Bourgeois, D.; Inquimbert, C.; Ottolenghi, L.; Carrouel, F. Periodontal Pathogens as Risk Factors of Cardiovascular Diseases, Diabetes, Rheumatoid Arthritis, Cancer, and Chronic Obstructive Pulmonary Disease—Is There Cause for Consideration? Microorganisms 2019, 7, 424. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.-H.; Giuliani, F. The Role of Inflammation in Depression and Fatigue. Front. Immunol. 2019, 10, 1696. [Google Scholar] [CrossRef] [Green Version]
- Xia, Z.; Depierre, J.W.; Nässberger, L. Tricyclic Antidepressants Inhibit IL-6, IL-1β and TNF-α Release in Human Blood Monocytes and IL-2 and Interferon-γ in T Cells. Immunopharmacology 1996, 34, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Su, K.-P.; Lai, H.-C.; Peng, C.-Y.; Su, W.-P.; Chang, J.P.-C.; Pariante, C.M. Interferon-Alpha-Induced Depression: Comparisons between Early- and Late-Onset Subgroups and with Patients with Major Depressive Disorder. Brain. Behav. Immun. 2019, 80, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Capuron, L.; Ravaud, A.; Dantzer, R. Early Depressive Symptoms in Cancer Patients Receiving Interleukin 2 and/or Interferon Alfa-2b Therapy. J. Clin. Oncol. 2000, 18, 2143–2151. [Google Scholar] [CrossRef] [PubMed]
- Carrouel, F.; Viennot, S.; Santamaria, J.; Veber, P.; Bourgeois, D. Quantitative Molecular Detection of 19 Major Pathogens in the Interdental Biofilm of Periodontally Healthy Young Adults. Front. Microbiol. 2016, 7, 840. [Google Scholar] [CrossRef]
- Bourgeois, D.; Bravo, M.; Llodra, J.-C.; Inquimbert, C.; Viennot, S.; Dussart, C.; Carrouel, F. Calibrated Interdental Brushing for the Prevention of Periodontal Pathogens Infection in Young Adults—A Randomized Controlled Clinical Trial. Sci. Rep. 2019, 9, 15127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inquimbert, C.; Bourgeois, D.; Bravo, M.; Viennot, S.; Tramini, P.; Llodra, J.C.; Molinari, N.; Dussart, C.; Giraudeau, N.; Carrouel, F. The Oral Bacterial Microbiome of Interdental Surfaces in Adolescents According to Carious Risk. Microorganisms 2019, 7, 319. [Google Scholar] [CrossRef] [Green Version]
- Alkahtani, S.; AL-Johani, N.S.; Alarifi, S. Mechanistic Insights, Treatment Paradigms, and Clinical Progress in Neurological Disorders: Current and Future Prospects. Int. J. Mol. Sci. 2023, 24, 1340. [Google Scholar] [CrossRef]
- Wareham, L.K.; Liddelow, S.A.; Temple, S.; Benowitz, L.I.; Di Polo, A.; Wellington, C.; Goldberg, J.L.; He, Z.; Duan, X.; Bu, G.; et al. Solving Neurodegeneration: Common Mechanisms and Strategies for New Treatments. Mol. Neurodegener. 2022, 17, 23. [Google Scholar] [CrossRef]
- Bjelobaba, I.; Savic, D.; Lavrnja, I. Multiple Sclerosis and Neuroinflammation: The Overview of Current and Prospective Therapies. Curr. Pharm. Des. 2017, 23, 693–730. [Google Scholar] [CrossRef]
- Leng, F.; Edison, P. Neuroinflammation and Microglial Activation in Alzheimer Disease: Where Do We Go from Here? Nat. Rev. Neurol. 2021, 17, 157–172. [Google Scholar] [CrossRef]
- Reale, M.; Iarlori, C.; Thomas, A.; Gambi, D.; Perfetti, B.; Di Nicola, M.; Onofrj, M. Peripheral Cytokines Profile in Parkinson’s Disease. Brain. Behav. Immun. 2009, 23, 55–63. [Google Scholar] [CrossRef]
- Dadas, A.; Washington, J.; Marchi, N.; Janigro, D. Chapter 2—Blood–Brain Barrier in Disease States. In Nervous System Drug Delivery; Lonser, R.R., Sarntinoranont, M., Bankiewicz, K., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 21–37. ISBN 978-0-12-813997-4. [Google Scholar]
- Kirk, J.; Plumb, J.; Mirakhur, M.; McQuaid, S. Tight Junctional Abnormality in Multiple Sclerosis White Matter Affects All Calibres of Vessel and Is Associated with Blood-Brain Barrier Leakage and Active Demyelination. J. Pathol. 2003, 201, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Shinohara, M.; Shinohara, M.; Yamazaki, A.; Murray, M.E.; Liesinger, A.M.; Heckman, M.G.; Lesser, E.R.; Parisi, J.E.; Petersen, R.C.; et al. Selective Loss of Cortical Endothelial Tight Junction Proteins during Alzheimer’s Disease Progression. Brain 2019, 142, 1077–1092. [Google Scholar] [CrossRef]
- Berriat, F.; Lobsiger, C.S.; Boillée, S. The Contribution of the Peripheral Immune System to Neurodegeneration. Nat. Neurosci. 2023, 26, 942–954. [Google Scholar] [CrossRef]
- Kempuraj, D.; Thangavel, R.; Selvakumar, G.P.; Zaheer, S.; Ahmed, M.E.; Raikwar, S.P.; Zahoor, H.; Saeed, D.; Natteru, P.A.; Iyer, S.; et al. Brain and Peripheral Atypical Inflammatory Mediators Potentiate Neuroinflammation and Neurodegeneration. Front. Cell. Neurosci. 2017, 11, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Ran, M.; Li, H.; Lin, Y.; Ma, K.; Yang, Y.; Fu, X.; Yang, S. New Insight into Neurological Degeneration: Inflammatory Cytokines and Blood–Brain Barrier. Front. Mol. Neurosci. 2022, 15, 1013933. [Google Scholar] [CrossRef] [PubMed]
- Galea, I. The Blood–Brain Barrier in Systemic Infection and Inflammation. Cell. Mol. Immunol. 2021, 18, 2489–2501. [Google Scholar] [CrossRef]
- Kaye, E.K.; Valencia, A.; Baba, N.; Spiro, A.; Dietrich, T.; Garcia, R.I. Tooth Loss and Periodontal Disease Predict Poor Cognitive Function in Older Men. J. Am. Geriatr. Soc. 2010, 58, 713–718. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, N.; Morikawa, M.; Tomioka, K.; Yanagi, M.; Amano, N.; Kurumatani, N. Association between Tooth Loss and the Development of Mild Memory Impairment in the Elderly: The Fujiwara-Kyo Study. J. Alzheimer’s Dis. 2015, 44, 777–786. [Google Scholar] [CrossRef]
- Nilsson, H.; Sanmartin Berglund, J.; Renvert, S. Longitudinal Evaluation of Periodontitis and Development of Cognitive Decline among Older Adults. J. Clin. Periodontol. 2018, 45, 1142–1149. [Google Scholar] [CrossRef]
- Saito, S.; Ohi, T.; Murakami, T.; Komiyama, T.; Miyoshi, Y.; Endo, K.; Satoh, M.; Asayama, K.; Inoue, R.; Kikuya, M.; et al. Association between Tooth Loss and Cognitive Impairment in Community-Dwelling Older Japanese Adults: A 4-Year Prospective Cohort Study from the Ohasama Study. BMC Oral Health 2018, 18, 142. [Google Scholar] [CrossRef]
- Iwasaki, M.; Kimura, Y.; Ogawa, H.; Yamaga, T.; Ansai, T.; Wada, T.; Sakamoto, R.; Ishimoto, Y.; Fujisawa, M.; Okumiya, K.; et al. Periodontitis, Periodontal Inflammation, and Mild Cognitive Impairment: A 5-Year Cohort Study. J. Periodontal Res. 2019, 54, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Huang, X.; Gong, Y.; Sun, J. Association between Tooth Loss Rate and Risk of Mild Cognitive Impairment in Older Adults: A Population-Based Longitudinal Study. Aging 2021, 13, 21599–21609. [Google Scholar] [CrossRef]
- Thomson, W.M.; Barak, Y. Tooth Loss and Dementia: A Critical Examination. J. Dent. Res. 2021, 100, 226–231. [Google Scholar] [CrossRef]
- Arrivé, E.; Letenneur, L.; Matharan, F.; Laporte, C.; Helmer, C.; Barberger-Gateau, P.; Miquel, J.L.; Dartigues, J.F. Oral Health Condition of French Elderly and Risk of Dementia: A Longitudinal Cohort Study. Community Dent. Oral Epidemiol. 2012, 40, 230–238. [Google Scholar] [CrossRef]
- Tzeng, N.-S.; Chung, C.-H.; Yeh, C.-B.; Huang, R.-Y.; Yuh, D.-Y.; Huang, S.-Y.; Lu, R.-B.; Chang, H.-A.; Kao, Y.-C.; Chiang, W.-S.; et al. Are Chronic Periodontitis and Gingivitis Associated with Dementia? A Nationwide, Retrospective, Matched-Cohort Study in Taiwan. Neuroepidemiology 2016, 47, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-T.; Lee, H.-C.; Hu, C.-J.; Huang, L.-K.; Chao, S.-P.; Lin, C.-P.; Su, E.C.-Y.; Lee, Y.-C.; Chen, C.-C. Periodontitis as a Modifiable Risk Factor for Dementia: A Nationwide Population-Based Cohort Study. J. Am. Geriatr. Soc. 2017, 65, 301–305. [Google Scholar] [CrossRef]
- Lee, Y.-L.; Hu, H.-Y.; Huang, L.-Y.; Chou, P.; Chu, D. Periodontal Disease Associated with Higher Risk of Dementia: Population-Based Cohort Study in Taiwan. J. Am. Geriatr. Soc. 2017, 65, 1975–1980. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, K.; Ohara, T.; Furuta, M.; Takeshita, T.; Shibata, Y.; Hata, J.; Yoshida, D.; Yamashita, Y.; Ninomiya, T. Tooth Loss and Risk of Dementia in the Community: The Hisayama Study. J. Am. Geriatr. Soc. 2017, 65, e95–e100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.; Kim, K.; Chang, J.; Kim, S.M.; Kim, S.J.; Cho, H.-J.; Park, S.M. Association of Chronic Periodontitis on Alzheimer’s Disease or Vascular Dementia. J. Am. Geriatr. Soc. 2019, 67, 1234–1239. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.-H.; Jeong, S.-N.; Lee, J.-H. Severe Periodontitis with Tooth Loss as a Modifiable Risk Factor for the Development of Alzheimer, Vascular, and Mixed Dementia: National Health Insurance Service-National Health Screening Retrospective Cohort 2002–2015. J. Periodontal Implant. Sci. 2020, 50, 303–312. [Google Scholar] [CrossRef]
- Lee, C.-Y.; Chang, C.-C.; Lin, C.-S.; Yeh, C.-C.; Hu, C.-J.; Wu, C.-Z.; Chen, T.-L.; Liao, C.-C. Risk of Dementia in Patients with Periodontitis and Related Protective Factors: A Nationwide Retrospective Cohort Study. J. Clin. Periodontol. 2020, 47, 1428–1436. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-R.; Son, M.; Kim, Y.-R.; Kang, H.-K. Risk of Dementia According to the Severity of Chronic Periodontitis in Korea: A Nationwide Retrospective Cohort Study. Epidemiol. Health 2022, 44, e2022077. [Google Scholar] [CrossRef] [PubMed]
- Holmer, J.; Eriksdotter, M.; Häbel, H.; Hed Myrberg, I.; Jonsson, A.; Pussinen, P.J.; Garcia-Ptacek, S.; Jansson, L.; Sandborgh-Englund, G.; Buhlin, K. Periodontal Conditions and Incident Dementia: A Nationwide Swedish Cohort Study. J. Periodontol. 2022, 93, 1378–1386. [Google Scholar] [CrossRef] [PubMed]
- Kiuchi, S.; Cooray, U.; Kusama, T.; Yamamoto, T.; Abbas, H.; Nakazawa, N.; Kondo, K.; Osaka, K.; Aida, J. Oral Status and Dementia Onset: Mediation of Nutritional and Social Factors. J. Dent. Res. 2022, 101, 420–427. [Google Scholar] [CrossRef] [PubMed]
- Ide, M.; Harris, M.; Stevens, A.; Sussams, R.; Hopkins, V.; Culliford, D.; Fuller, J.; Ibbett, P.; Raybould, R.; Thomas, R.; et al. Periodontitis and Cognitive Decline in Alzheimer’s Disease. PLoS ONE 2016, 11, e0151081. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-K.; Wu, Y.-T.; Chang, Y.-C. Association between Chronic Periodontitis and the Risk of Alzheimer’s Disease: A Retrospective, Population-Based, Matched-Cohort Study. Alzheimer’s Res. Ther. 2017, 9, 56. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.C.; Sheu, J.J.; Lin, H.C.; Jensen, D.A. Increased Risk of Parkinsonism Following Chronic Periodontitis: A Retrospective Cohort Study. Mov. Disord. 2013, 28, 1307–1308. [Google Scholar] [CrossRef]
- Chen, C.-K.; Wu, Y.-T.; Chang, Y.-C. Periodontal Inflammatory Disease Is Associated with the Risk of Parkinson’s Disease: A Population-Based Retrospective Matched-Cohort Study. PeerJ 2017, 5, e3647. [Google Scholar] [CrossRef] [Green Version]
- Woo, H.G.; Chang, Y.; Lee, J.S.; Song, T.-J. Association of Tooth Loss with New-Onset Parkinson’s Disease: A Nationwide Population-Based Cohort Study. Park. Dis. 2020, 2020, 4760512. [Google Scholar] [CrossRef]
- Jeong, E.; Park, J.-B.; Park, Y.-G. Evaluation of the Association between Periodontitis and Risk of Parkinson’s Disease: A Nationwide Retrospective Cohort Study. Sci. Rep. 2021, 11, 16594. [Google Scholar] [CrossRef]
- Kamer, A.R.; Craig, R.G.; Pirraglia, E.; Dasanayake, A.P.; Norman, R.G.; Boylan, R.J.; Nehorayoff, A.; Glodzik, L.; Brys, M.; de Leon, M.J. TNF-α and Antibodies to Periodontal Bacteria Discriminate between Alzheimer’s Disease Patients and Normal Subjects. J. Neuroimmunol. 2009, 216, 92–97. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.-P.; Lee, Y.-P.; Hwang, M.-J.; Chiang, C.-P. Fusobacterium Nucleatum-Caused Brain Abscess—Case Report. J. Dent. Sci. 2021, 16, 776–777. [Google Scholar] [CrossRef] [PubMed]
- Foschi, F.; Izard, J.; Sasaki, H.; Sambri, V.; Prati, C.; Müller, R.; Stashenko, P. Treponema Denticola in Disseminating Endodontic Infections. J. Dent. Res. 2006, 85, 761–765. [Google Scholar] [CrossRef] [Green Version]
- Schwahn, C.; Frenzel, S.; Holtfreter, B.; Van der Auwera, S.; Pink, C.; Bülow, R.; Friedrich, N.; Völzke, H.; Biffar, R.; Kocher, T.; et al. Effect of Periodontal Treatment on Preclinical Alzheimer’s Disease—Results of a Trial Emulation Approach. Alzheimer’s Dement. 2022, 18, 127–141. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Zhang, S.; Duan, L.; Yang, F.; Zhang, K.; Yan, F.; Ge, S. Periodontitis Deteriorates Cognitive Function and Impairs Neurons and Glia in a Mouse Model of Alzheimer’s Disease. J. Alzheimer’s Dis. 2021, 79, 1785–1800. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.; Qian, X.; Wang, Q.; Huang, L.; Ge, S. Experimental Periodontitis Deteriorates Cognitive Function and Impairs Insulin Signaling in a Streptozotocin-Induced Alzheimer’s Disease Rat Model. J. Alzheimer’s Dis. 2022, 88, 57–74. [Google Scholar] [CrossRef]
- Wang, X.; Tong, Y.; Zhang, J.; Khan, N.; Zhang, K.; Bai, H.; Zhang, Q.; Chen, Y. Neuroinflammation Changes with Periodontal Inflammation Status during Periodontitis in Wild-Type Mice. Oral Dis. 2021, 27, 1001–1011. [Google Scholar] [CrossRef]
- Feng, Y.-K.; Wu, Q.-L.; Peng, Y.-W.; Liang, F.-Y.; You, H.-J.; Feng, Y.-W.; Li, G.; Li, X.-J.; Liu, S.-H.; Li, Y.-C.; et al. Oral P. gingivalis Impairs Gut Permeability and Mediates Immune Responses Associated with Neurodegeneration in LRRK2 R1441G Mice. J. Neuroinflamm. 2020, 17, 347. [Google Scholar] [CrossRef]
- Ilievski, V.; Zuchowska, P.K.; Green, S.J.; Toth, P.T.; Ragozzino, M.E.; Le, K.; Aljewari, H.W.; O’Brien-Simpson, N.M.; Reynolds, E.C.; Watanabe, K. Chronic Oral Application of a Periodontal Pathogen Results in Brain Inflammation, Neurodegeneration and Amyloid Beta Production in Wild Type Mice. PLoS ONE 2018, 13, e0204941. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-X.; Kang, X.-N.; Cao, Y.; Zheng, D.-X.; Lu, Y.-M.; Pang, C.-F.; Wang, Z.; Cheng, B.; Peng, Y. Porphyromonas Gingivalis Induces Depression via Downregulating P75NTR-Mediated BDNF Maturation in Astrocytes. Brain Behav. Immun. 2019, 81, 523–534. [Google Scholar] [CrossRef]
- Bachtiar, E.W.; Septiwidyati, T.R. Possible Role of Porphyromonas Gingivalis in the Regulation of E2F1, CDK11, and INOS Gene Expression in Neuronal Cell Cycle: A Preliminary Study. J. Int. Soc. Prev. Community Dent. 2021, 11, 582–587. [Google Scholar] [CrossRef]
- Su, X.; Tang, Z.; Lu, Z.; Liu, Y.; He, W.; Jiang, J.; Zhang, Y.; Wu, H. Oral Treponema Denticola Infection Induces Aβ1–40 and Aβ1–42 Accumulation in the Hippocampus of C57BL/6 Mice. J. Mol. Neurosci. 2021, 71, 1506–1514. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Su, X.; Tang, Z.; Jian, L.; Zhu, H.; Cheng, X.; Wu, H. Treponema Denticola Induces Neuronal Apoptosis by Promoting Amyloid-β Accumulation in Mice. Pathogens 2022, 11, 1150. [Google Scholar] [CrossRef]
- Gustavsen, M.W.; Celius, E.G.; Moen, S.M.; Bjølgerud, A.; Berg-Hansen, P.; Nygaard, G.O.; Sandvik, L.; Lie, B.A.; Harbo, H.F. No Association between Multiple Sclerosis and Periodontitis after Adjusting for Smoking Habits. Eur. J. Neurol. 2015, 22, 588–590. [Google Scholar] [CrossRef]
- Al-Ansari, A. Is There an Association between Multiple Sclerosis and Oral Health? Evid. Based Dent. 2021, 22, 44–45. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Strain | Genome Size (bp) | Capsule | Fimbriae/Type | Mono-Species Biofilm Formation | Gingipain Activity | Virulence Potential | Citation |
|---|---|---|---|---|---|---|---|
| ATCC 33277 | 2,354,886 | none | Abundant/Type I | Moderate | High | Moderate | [15,32,33,34,35,36,37,38] |
| FDC 381 | 2,378,872 | none | Abundant/Type I | Moderate | Medium | Moderate | [15,33,34,36,37,38,39] |
| W83 | 2,343,476 | K1 encapsulated | Poor/Type IV | None | High | High | [15,32,34,35,36,37,38] |
| W50 | 2,345,841 | K1 encapsulated | Poor/Type IV | None | High | High | [15,34,36,38,40] |
| Bacteria | Component | Function | Citation |
|---|---|---|---|
| Porphyromonas gingivalis | LPS | Induces inflammation and activates the host immune system | [20,21] |
| Gingipains | Accounts for 85% of the total proteolytic activity; In addition to increasing nutrient availability, it has an immunoregulatory purpose and plays a role in bacterial coaggregation | [19,20] | |
| Fimbriae | Promotes adhesion, invasion of host cells, biofilm formation, and bacterial motility | [32] | |
| GroEL | Induces inflammation and activates the host immune system | [97] | |
| Hemagglutinins | Promotes adherence to host cells and is used for heme acquisition | [98] | |
| Capsule | Polysaccharide layer that protects the bacterium from phagocytosis by host immune cells | [33] | |
| Tannerella forsythia | LPS | Induces inflammation and activates the host immune system | [99] |
| PrtH | Protease that plays a role in cell detachment | [41,43] | |
| BspA | Necessary for attachment and invasion of epithelial cells | [41,45] | |
| Karilysin | Immunoregulatory, causes bacterial serum resistance | [50] | |
| Methylglyoxal | Causes inflammation and tissue damage | [47] | |
| GroEL | Induces inflammation and activates the host immune system | [48] | |
| S-layer | Involved in coaggregation, immunoevasion, adhesion, and invasion of cells | [52,53] | |
| Treponema denticola | LOS | Impacts adherence to host cells and bacterial coaggregation | [63] |
| MSP | induces inflammatory responses and forms cytotoxic pores | [60,61,62] | |
| Dentilsin | Stimulates tissue destruction and activates C3 complement pathway; it also has an immunoregulatory purpose and plays a role in bacterial coaggregation | [64,65,66,71,72] | |
| Leucine-rich-repeat A | Impacts adherence and invasion of host cells, while also being important for bacterial coaggregation | [100] | |
| Fusobacterium nucleatum | LPS | Induces inflammation and activates the host immune system | [79,80] |
| Main Adhesin (FadA) | Required for the invasion and adhesion to host cells | [80] | |
| Other Adhesins (Fap2, RadD, and aid1) | Impacts binding to a variety of different bacteria and host proteins | [74,80] | |
| Fusolisin | Impacts nutrient availability while also having an immunoregulatory role | [81] | |
| Aggregatibacter actinomycetemcomitans | LPS | Induces inflammation and activates the host immune system | [86,87] |
| Leukotoxins | Disrupts immune function | [86,91] | |
| CTD | Induces DNA damage and cell cycle arrest which leads and apoptosis, with a pronounced impact on immune cells | [92,93] | |
| Fimbriae | Impacts adherence and bacterial coaggregation | [101] |
| Disease Investigated | Study Design/Duration | Question | Adjusting for: | Number of Participants/Age [Years] | Results | Author, Date, Journal, and Citation |
|---|---|---|---|---|---|---|
| Mild cognitive impairment (MCI) | Prospective cohort study (8 years) | Does periodontitis correlate with cognitive decline? | Age, gender, race, education, income, smoking, alcohol consumption, and diabetes | N = 558 (total) Age group = 52–75 Age mean = 64.7 ± 4.3 | No significant correlation was found between tooth loss at baseline and cognitive capabilities. | Supawadee Naorungroj et al. (2014) Community Dentistry and Oral Epidemiology’s [145] |
| Mild memory impairment (MMI) | Prospective cohort study (5 years) | Does periodontitis correlate with a decline in memory? | Age, gender, MMSE-total, education length, smoking, drinking, blood pressure, cancer, myocardial infarction, cerebrovascular disease, diabetes mellitus, hypertension, and dyslipidemia | N = 2335 (total) Age group ≥ 65 Age median = 71 | Each tooth lost at baseline slightly increased the odds of developing memory impairment: (OR = 1.02; 95% Cl 1.00–1.03; p = 0.039) | Nozomi Okamoto et al. (2015) Journal of Alzheimer’s Disease [146] |
| Mild cognitive impairment (MCI) | Prospective cohort study (6 years) | Does periodontitis correlate with cognitive decline? | Age, gender, living alone, education, alcohol consumption, BMI ischemic heart disease, traumatic brain injury, and depression | N = 715 (total) N = 214 (with periodontitis) Age group = 60–96 Age mean = NA | Bone loss caused by periodontitis may be associated with faster cognitive decline: (OR = 2.2; 95% CI 1.2–3.8; NA) | Helena Nilsson et al. (2018) Journal of Clinical Periodontology [147] |
| Mild cognitive impairment (MCI) | Prospective cohort study (4 years) | Does periodontitis correlate with cognitive decline? | Age, gender, hypertension, diabetes, cerebrovascular/cardiovascular disease, hypercholesterolemia, depressive symptoms, body mass index, smoking status, drinking status, duration of education, and baseline MMSE score | N = 140 (total) Age group ≥ 65 Age mean = 70.9 ± 4.3 | Tooth loss may be associated with cognitive impairment: (OR = 3.31; 95% Cl 1.07–10.2; p = 0.037) | Sho Saito et al. (2018) BMC Oral Health [148] |
| Mild cognitive impairment (MCI) | Longitudinal cohort study (5 years) | Is there an association between mild cognitive impairment and periodontitis? | Age, sex, smoking status, educational level, physical activity level, obesity, depression, and diabetes | N = 179 (total) Age group ≥ 75 Age mean = 80.1 ± 4.4 | Severe periodontitis was significantly associated with MCI: (OR = 3.58; 95% Cl 1.45–8.87; p < 0.01) | Masanori Iwasaki et al. (2019) Journal of Periodontal Research [149] |
| Mild cognitive impairment (MCI) | Longitudinal study (median following of 5.93 years) | Is there an association between mild cognitive impairment and severity of tooth loss? | Age, gender, ethnicity, residence, marriage status, occupation, education, smoking and drinking, activities of daily life (ADL) score, physical performance score, food diversity score, social activity score, and chronic disease score | N = 11862 (total) N = 3966 (developed MCI) Age group ≥ 65 Age mean = 81.41 | Higher tooth loss rate was associated with an increased risk of MCI in elderly subjects. 1–2 tooth loss per year [mild] (OR = 1.16; 95% Cl 1.04–1.29; NA) >2 tooth loss per year [severe] (OR = 1.28; 95% Cl 1.17–1.40; NA) | Shuyu Xu et al. (2021) Aging [150] |
| Dementia | Prospective study (longitudinal checkup every 3 years for up to 32 years) | Does severity of tooth loss correlate with dementia progression? | Age, years of education, smoking, BMI, medication use, intake of substances, coronary heart disease, stroke, hypertension, cardiovascular disease, cancer, and diabetes | N = 597 (total) Age group = 28–70 Age mean = 48 ± 8 | Each tooth lost per decade since the baseline examination increased the risks of lowering the Mini-Mental State Examination score: (HR = 1.12, 95% CI 1.05–1.18; p < 0.05) | Kaye et al. (2010) Journal of the American Geriatrics Society [151] |
| Dementia | Longitudinal cohort study (5 years) | Are periodontal conditions associated with dementia onset? | Age, gender, educational level, living condition, tobacco intake, alcohol consumption, BMI, stroke, angina pectoris, diabetes, hypertension, depression, and myocardial infarction | N = 348 (total) N = 246(with some form of periodontal condition) Age group = 66–80 Age median = 70 | Periodontal condition was not associated with an increased risk of dementia: (HR = 1.13; 95% CI 0.60–2.12; NA) | E. Arrivé et al. (2011) Community Dentistry and Oral Epidemiology’s [152] |
| Dementia | Retrospective matched-cohort study (10 years) | Is there an Association between Chronic Periodontitis and Gingivitis with Dementia? | Age, sex, monthly income, urbanization level, geographic region, hypertension, diabetes, hyperlipidemia, obesity, depression, chronic kidney disease, and stroke | N = 8828 (total) N = 2207 (with periodontitis or gingivitis) Age group ≥ 20 Age mean = NA | Patients with chronic periodontitis and gingivitis have a higher risk of developing dementia: (HR = 2.54; 95% CI 1.297–3.352; p = 0.002). | Nian-Sheng Tzeng et al. (2016) Journal of Neuroepidemiology [153] |
| Dementia | Prospective matched-cohort study (13 years) | Is there an association of chronic periodontitis with dementia onset? | Age, gender, geographic region, urbanization level, hypertension, diabetes mellitus, cardiovascular disease, congestive heart failure, atrial fibrillation, stroke, and chronic renal disease | N = 6056 (total) N = 3028 (with periodontitis) Age group ≥ 65 Age mean = 72.4 ± NA | Patients with periodontitis were at higher risk of developing dementia: (HR = 1.16; 95% Cl 1.01–1.32; p = 0.03) | Yao-Tung Lee et. al (2016) Journal of the American Geriatrics Society [154] |
| Dementia | Retrospective cohort study (10 years) | Does treatment of periodontitis affect dementia onset and progression? | Age, gender, socioeconomic status, residential urbanicity, hypertension, diabetes, and hyperlipidemia | N = 182,747 (total—all with periodontitis) N = 19,674 (did not receive treatment for periodontitis) N = 6133 (developed dementia) Age group ≥ 45 Age mean = NA | Subjects who had not received Periodontitis treatment were at greater risk of developing dementia: (HR = 1.14; 95% CI 1.04–1.24; p ≤ 0.001) | Ya-Ling Lee et al. (2017) Journal of the American Geriatrics Society [155] |
| Dementia | Prospective cohort study (5 years) | Does severity of tooth loss correlate with dementia onset? | Age, gender, occupation, educational level, tobacco intake, alcohol consumption, BMI, stroke, angina pectoris, diabetes, hypertension, and tooth brushing frequency | N = 1566 (total) N = 180 (developed dementia) Age group ≥ 60 Age mean = NA | Tooth loss may be associated with a higher risk of dementia: 10–19 teeth: (HR = 1.62, 95% CI = 1.06–2.46), 1–9 teeth (HR = 1.81, 95% CI = 1.11–2.94), 0 teeth (HR = 1.63, 95% CI = 0.95–2.80) | Kenji Takeuchi et al. (2017) Journal of the American Geriatrics Society [156] |
| Dementia | Retrospective matched-cohort study (10 years) | Is there an association of chronic periodontitis with dementia onset? | Age, sex, household income, smoking status, alcohol consumption, physical activity, BMI, blood pressure, fasting serum glucose, total cholesterol, and Charlson Comorbidity Index | N = 262,349 (total) N = 46,344 (with periodontitis) Age group ≥ 50 Age mean = 60.2 ± 7.3 | Chronic periodontitis may be associated with a higher risk of developing dementia: Overall dementia (HR = 1.06; 95% CI 1.01–1.11; p = 0.015) Alzheimer’s (HR = 1.05; 95% CI 1.00–1.11; p = 0.042). The risk-increasing effect of chronic periodontitis on dementia tended to be stronger among participants with healthy lifestyle behaviors. | Seulggie Choi et al. (2019) Journal of the American Geriatrics Society [157] |
| Dementia | Retrospective matched-cohort study (14 years) | Is severe periodontitis with tooth loss a risk factor for the onset of dementia? | Age, sex, household income, insurance status, BMI, total cholesterol level, smoking status, drinking status, physical activity, hypertension, and diabetes mellitus | N = 20,230 (total) N = 10,115 (with periodontitis) Age group = 40–80 Age mean = NA | Patients with severe periodontitis with 1–9 remaining teeth were at higher risk for developing dementia: Alzheimer’s (HR = 1.08; 95% CI 1.01–1.14; p = 0.022) Vascular dementia (HR = 1.24; 95% CI 1.16–1.32; p < 0.001) Mixed dementia (HR = 1.16; 95% CI 1.09–1.24; p < 0.001) | Do-Hyung Kim et al. (2020) Journal of periodontal and implant science [158] |
| Dementia | Retrospective matched-cohort study (14 years) | Is there an association of chronic periodontitis with dementia onset? | Age, gender, influenza vaccination, income, hypertension, mental disorders, diabetes, ischemic heart disease, stroke, hyperlipidemia, chronic obstructive pulmonary disease, heart failure, liver cirrhosis, and traumatic brain injury | N = 102,036 (total) N = 56,018 (with periodontitis) Age group ≥ 50 Age mean = NA | Patients with periodontitis were at higher risk of developing dementia: (HR = 1.73; 95% CI 1.61–1.86; p < 0.0001) Periodontitis was associated with a higher risk of developing dementia in people with no underlying medical conditions: =0 (HR 6.16; 95% CI 5.13–7.40; p < 0.05) =1 (HR 1.27; 95% CI 1.10–1.46; p < 0.05) ≥2 (HR 1.20; 95% CI 1.09–1.31; p < 0.05) | Chia-Yen Lee et al. (2020) Journal of Clinical Periodontology [159] |
| Dementia | Retrospective longitudinal matched-cohort study (13 years with median following of 6.6 years) | Is there a difference in dementia onset between groups of people severe or mild chronic periodontitis? | Age, gender, income level, smoking status, alcohol consumption, regular exercise, BMI, systolic blood pressure, diastolic blood pressure, fasting blood glucose levels, total cholesterol levels, hypertension, diabetes, dyslipidemia, heart disease, cerebrovascular disease, depression, and Charlson Comorbidity Index | N = 8624 (with mild chronic periodontitis) N = 8624 (with severe chronic periodontitis) Age group ≥ 60 Age mean = 70.9 ± 4.8 | Dementia onset was significantly associated with the severity of periodontitis. (HR = 1.15; 95% CI; 1.04–1.27; p = 0.01) | Seon-Rye Kim et al. (2022) Epidemiology and health [160] |
| Dementia | Cohort study (Follow up 7.6 ± 1.1 years) | Is there an association of deep probing pocket depths and tooth count with dementia onset? | Age, gender, civil status, disposable income, education, geographical area, and Charlson Comorbidity Index | N = 37,174 (total) N = 7992 (with ≥4 teeth and/or dental implants with probing pocket depth ≥6 mm) Age group = 40–80 Age mean = 61 | Tooth loss and deep probing pocket depth does not increase the risk of developing dementia: (HR = 1.13; 95% CI 0.39–3.24; p > 0.05) | Jacob Holmer et al. (2022) Journal of Clinical Periodontology [161] |
| Dementia | Prospective cohort study (6 years) | Does tooth loss increase the risk of dementia onset? | Age, gender, marital status, denture use, education level, income level, social network, smoking status, alcohol consumption, diabetes treatment, and hypertension | N = 35,744 (total) N = 22,164 (<20 teeth) Age group ≥ 65 Age mean = 73.2 ± 5.5 | Number of teeth may increase the risk of dementia onset: (HR = 1.14; 95% CI 1.01–1.28 p = 0.041) When the nutritional and social mediators (weight loss, vegetable and fruit intake, and regular exercise) were included, the effect was reduced to: HR = 1.10 95% CI; 0.98–1.25; p = 0.113) | S Kiuchi et al. (2022) Journal of Dental Research [162] |
| Alzheimer’s disease (AD) | Case-control study with a follow-up over a six-month period | Does periodontitis affect the speed of cognitive decline? | Age, gender, and baseline cognitive status | N = 52 (total) N = 20 (with periodontitis) Age group = NA Age mean = 77.7 ± 8.6 | The presence of periodontitis at baseline was not related to baseline cognitive state: (MD = 1.1; 95% Cl -5.6–7.7; p = 0.8) Rate of cognitive decline: (MD = 4.9; 95% Cl 1.2–8.6; p = 0.01) | Mark Ide et al. (2016) PlOS One [163] |
| Alzheimer’s disease (AD) | Retrospective matched-cohort study (8 years) | Is there an association between chronic periodontitis and the risk of Alzheimer’s disease? | Age, gender, index year, urbanization level, hypertension, hyperlipidemia, chronic kidney disease, depression, stroke, diabetes mellitus, and traumatic brain injury | N = 27,963 (total) N = 9291 (with periodontitis) Age group ≥ 50 Age mean = 54.1 ± 10.5 | The association between chronic periodontitis and AD was significant in patients that have had over 10 years of exposure to chronic periodontitis: (HR = 1.707; 95% CI 1.152–2.528; p = 0.0077). | Chang-Kai Chen et al. (2017) Alzheimer’s Research and Therapy [164] |
| Parkinson’s disease (PD) | Retrospective matched-cohort study (5 years) | Does chronic periodontitis increase the risk of PD incidence? | Age, gender, urbanization level of residence, index year, stroke, and depression | N = 320,106 (total N = 53,351 (with periodontitis) Age group ≥ 40 Age mean = NA | Chronic periodontitis may increase the risk of Parkinson’s disease: (HR = 1.43; 95% 1.32–1.55; NA) | Tsai-Ching Liu et al. (2013) Movement Disorders [165] |
| Parkinson’s disease (PD) | Retrospective matched-cohort study (8 years) | Does periodontitis and its severity correlate with PD incidence? | Age, gender, CCI score, urbanization level, stroke, depression, hyperlipidemia, cancer, chronic kidney disease, and traumatic brain injury | N = 16,188 (total) N = 5396 (with periodontitis) Age group ≥ 40 Age mean = 54.1 ± 10.5 | Periodontitis correlates with the incidence rate of PD: (HR = 1.431; 95% CI 1.141–1.794; p = 0.002) | Chang-Kai Chen et al. (2017) Peer.J [166] |
| Parkinson’s disease (PD) | Longitudinal cohort study (10.4 years) | Does chronic periodontitis increase the risk of PD incidence? | Age, gender, income level, body mass index, lifestyle habits, such as alcohol intake, smoking status, physical activity, blood pressure, renal disease, hypertension, diabetes mellitus, and dyslipidemia | N = 153,165 N ≈ 30,480 (with periodontitis) Age group ≥ 40 Age mean = 52.8 | Chronic periodontitis did not significantly increase the risk of PD incidence: (HR = 0.91; 95% Cl 0.78–1.06; p = 0.232) Tooth loss (≥15) correlated to higher PD incidence: (HR = 1.38; 1.03–1.85; p = 0.029) | Ho Geol Woo et al. (2020) Parkinson’s Disease [167] |
| Parkinson’s disease (PD) | Retrospective cohort study in South Korea (8 years) | Do periodontitis and its severity correlate with PD incidence? | Age, sex, smoking status, drinking habits, exercise habits, income level, BMI, diabetes mellitus, hypertension, dyslipidemia, stroke, and depression | N = 6,856,180(total) N = 903,063 (with periodontitis) Age group ≥ 40 Age mean = 55.47 ± 9.97 | The severity of periodontitis correlates with the incidence rate of PD: (HR = 1.114; 95% CI 1.062–1.146; p < 0.001) | Eunkyung Jeong et al. (2021) Scientific report—Nature [168] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Visentin, D.; Gobin, I.; Maglica, Ž. Periodontal Pathogens and Their Links to Neuroinflammation and Neurodegeneration. Microorganisms 2023, 11, 1832. https://doi.org/10.3390/microorganisms11071832
Visentin D, Gobin I, Maglica Ž. Periodontal Pathogens and Their Links to Neuroinflammation and Neurodegeneration. Microorganisms. 2023; 11(7):1832. https://doi.org/10.3390/microorganisms11071832
Chicago/Turabian StyleVisentin, David, Ivana Gobin, and Željka Maglica. 2023. "Periodontal Pathogens and Their Links to Neuroinflammation and Neurodegeneration" Microorganisms 11, no. 7: 1832. https://doi.org/10.3390/microorganisms11071832
APA StyleVisentin, D., Gobin, I., & Maglica, Ž. (2023). Periodontal Pathogens and Their Links to Neuroinflammation and Neurodegeneration. Microorganisms, 11(7), 1832. https://doi.org/10.3390/microorganisms11071832