
The Potential of Digested Sludge-Assimilating Microflora for Biogas Production from Food Processing Wastes

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
2.2. Preparation of Powders of FPWs
2.3. Seed Culture
2.4. Vial-Scale Biogas Fermentation
2.5. Biogas Analysis
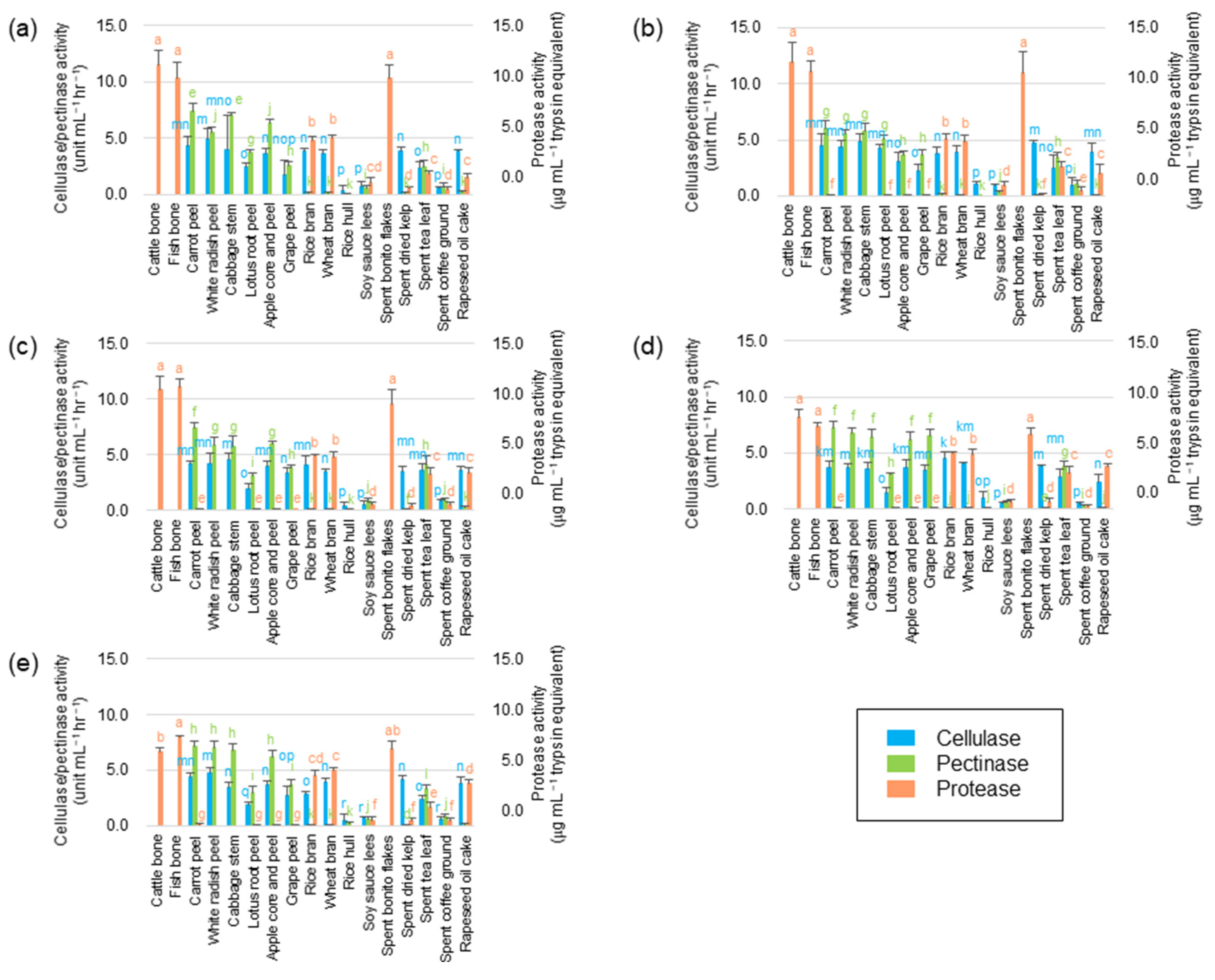
2.6. Enzymatic Assay
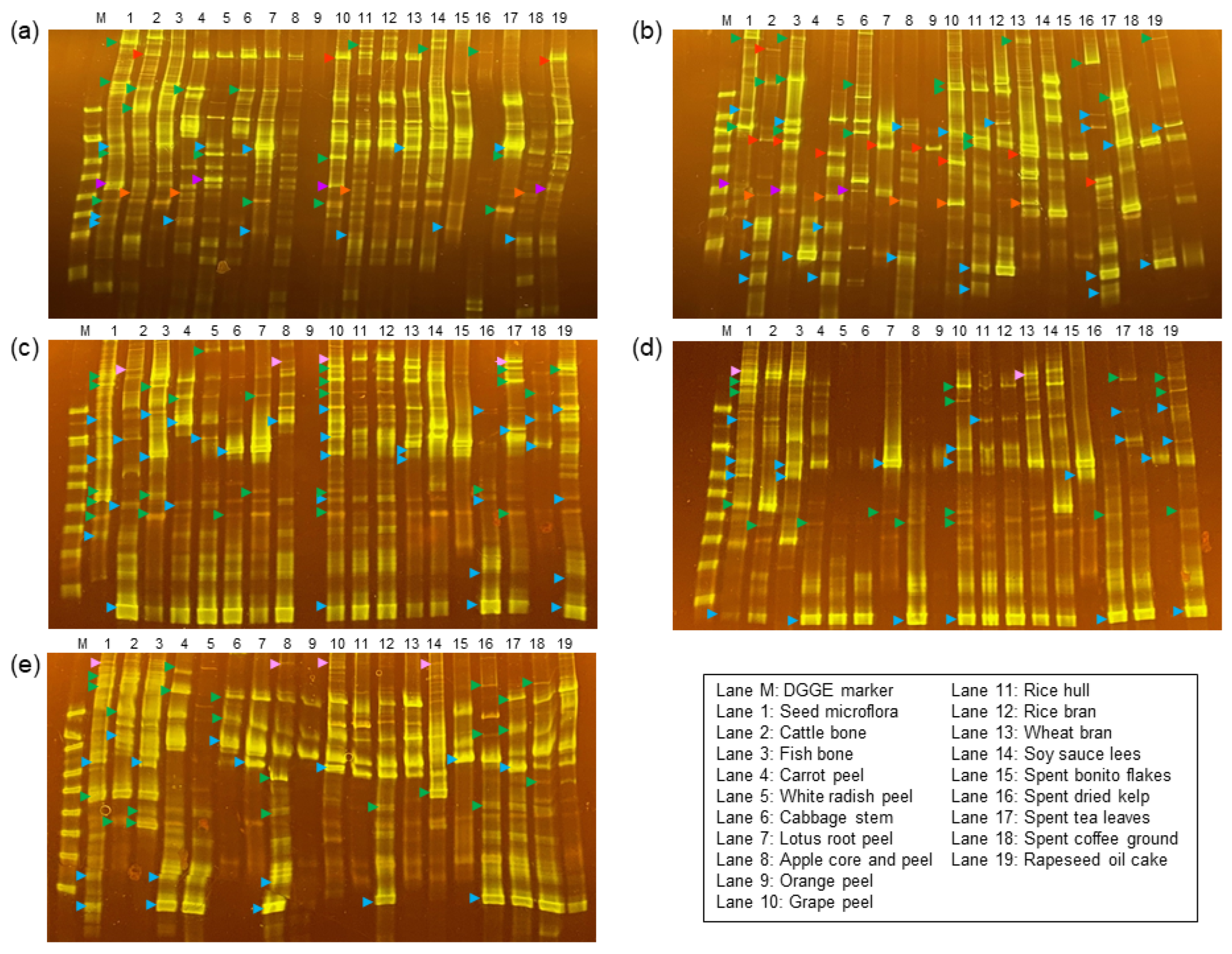
2.7. Community Fingerprinting of Microflorae
2.8. DNA Sequencing of the PCR-DGGE Amplicons
2.9. Statistical Analysis
3. Results and Discussion
3.1. Biogas Production from FPWs by the Subcultured Microflorae
3.2. Eubacterial Compositions of the Subcultured Microflorae
3.3. Archaea Compositions of the Subcultured Microflorae
3.4. Enzymatic Activity in Subcultured Microflorae
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- UNEP. Food waste amounts: Measured estimates and extrapolations. In Food Waste Index Report 2021; UNEP: Nairobi, Kenya, 2021; pp. 54–72. Available online: https://www.unep.org/resources/report/unep-food-waste-index-report-2021 (accessed on 25 July 2023).
- FAO. Food Loss and Waste—Framing the Issues. In The State of Food and Agriculture; FAO: Rome, Italy, 2019; pp. 1–19. Available online: https://www.fao.org/3/ca6030en/ca6030en.pdf (accessed on 25 July 2023).
- Zhang, C.; Kang, X.; Wang, F.; Tian, Y.; Liu, T.; Su, Y.; Qian, T.; Zhang, Y. Valorization of food waste for cost-effective reducing sugar recovery in a two-stage enzymatic hydrolysis platform. Energy 2020, 208, 118379. [Google Scholar] [CrossRef]
- Ali, A.; Bux Mahar, R.; Panhwar, S.; Ahmed Keerio, H.; Hussain Khokhar, N.; Suja, F.; Rundong, L. Generation of green renewable energy through anaerobic digestion technology (ADT): Technical insights review. Waste Biomass Valoriz. 2023, 14, 663–686. [Google Scholar] [CrossRef]
- Ferdeș, M.; Zăbavă, B.Ș.; Paraschiv, G.; Ionescu, M.; Dincă, M.N.; Moiceanu, G. Food waste management for biogas production in the context of sustainable development. Energies 2022, 15, 6268. [Google Scholar] [CrossRef]
- Food Waste Reduction Alliance. Analysis of U.S. Food Waste among Food Manufacturers, Retailers, and Restaurants. 2016. Available online: https://foodwastealliance.org/wp-content/uploads/2020/05/FWRA-Food-Waste-Survey-2016-Report_Final.pdf (accessed on 25 July 2023).
- Ministry of Agriculture, Forestry and Fisheries of Japan. Annual Amount of Food Waste Generated and Recycling Rate of Cyclical Food Resources. 2019. Available online: http://www.maff.go.jp/j/shokusan/recycle/syokuhin/kouhyou.html (accessed on 25 July 2023). (In Japanese).
- Mehrabi, Z. Likely decline in the number of farms globally by the middle of the century. Nat. Sustain. 2023, 6, 949–954. [Google Scholar] [CrossRef]
- Ma, S.; Jiang, F.; Huang, Y.; Zhang, Y.; Wang, S.; Fan, H.; Liu, B.; Li, Q.; Yin, L.; Wang, H.; et al. A microbial gene catalog of anaerobic digestion from full-scale biogas plants. GigaScience 2021, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Roy, P.; Mohanty, A.K.; Dick, P.; Misra, M. A review on the challenges and choices for food waste valorization: Environmental and economic impacts. ACS Environ. 2023, 3, 58–75. [Google Scholar] [CrossRef]
- Xu, F.; Li, Y.; Ge, X.; Yang, L.; Li, Y. Anaerobic digestion of food waste–Challenges and opportunities. Bioresour. Technol. 2018, 247, 1047–1058. [Google Scholar] [CrossRef]
- Polizzi, C.; Alatriste-Mondragón, F.; Munz, G. The role of organic load and ammonia inhibition in anaerobic digestion of tannery fleshing. Water Resour. Ind. 2018, 19, 25–34. [Google Scholar] [CrossRef]
- Lizundia, E.; Luzi, F.; Puglia, D. Organic waste valorization towards circular and sustainable biocomposites. Green Chem. 2022, 24, 5429–5459. [Google Scholar] [CrossRef]
- Kon, A.; Omata, S.; Hayakawa, Y.; Aburai, N.; Fujii, K. Microflora communities which can convert digested sludge to biogas. Environ. Technol. 2022, 43, 2391–2403. [Google Scholar] [CrossRef]
- Hayakawa, Y.; Aburai, N.; Fujii, K. Development of digested sludge-assimilating and biohydrogen-yielding microflorae. Fermentation 2023, 9, 175. [Google Scholar] [CrossRef]
- Santiago, S.G.; Trably, E.; Latrille, E.; Buitron, G.; Moreno-Andrade, I. The hydraulic retention time influences the abundance of Enterobacter, Clostridium and Lactobacillus during the hydrogen production from food waste. Lett. Appl. Microbiol. 2019, 69, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.H.; Sim, Y.B.; Baik, J.H.; Park, J.H.; Kim, S.M.; Yang, J.; Kim, S.H. Effect of genus Clostridium abundance on mixed-culture fermentation converting food waste into biohydrogen. Bioresour. Technol. 2021, 342, 125942. [Google Scholar] [CrossRef] [PubMed]
- Nzeteu, C.; Joyce, A.; Thorn, C.; McDonnell, K.; Shirran, S.; O’Flaherty, V.; Abram, F. Resource recovery from the anaerobic digestion of food waste is underpinned by cross-kingdom microbial activities. Bioresour. Technol. Rep. 2021, 16, 100847. [Google Scholar] [CrossRef]
- Xiao, L.; Deng, Z.; Fung, K.Y.; Ng, K.M. Biohydrogen generation from anaerobic digestion of food waste. Int. J. Hydrogen Energy 2013, 38, 13907–13913. [Google Scholar] [CrossRef]
- Torres-León, C.; Chávez-González, M.L.; Hernández-Almanza, A.; Martínez-Medina, G.A.; Ramírez-Guzmán, N.; Londoño-Hernández, L.; Aguilar, C.N. Recent advances on the microbiological and enzymatic processing for conversion of food wastes to valuable bioproducts. Curr. Opin. Food Sci. 2021, 38, 40–45. [Google Scholar] [CrossRef]
- Al Talebi, Z.A.; Al-Kawaz, H.S.; Mahdi, R.K.; Al-Hassnawi, A.T.; Alta’ee, A.H.; Hadwan, A.M.; Abbas khudhair, D.; Hadwan, M.H. An optimized protocol for estimating cellulase activity in biological samples. Anal. Biochem. 2022, 655, 114860. [Google Scholar] [CrossRef]
- Heuer, H.; Krsek, M.; Baker, P.; Smalla, K.; Wellington, E.M.H. Analysis of actinomycete communities by specific amplification of genes encoding 16S rRNA and gel electrophoretic separation in denaturing gradients. Appl. Environ. Microbiol. 1997, 63, 3233–3241. [Google Scholar] [CrossRef]
- Zhou, M.; Hernandez-Sanabria, E.; Guan, L.L. Characterization of variation in rumen methanogenic communities under different dietary and host feed efficiency conditions, as determined by PCR-denaturing gradient gel electrophoresis analysis. Appl. Environ. Microbiol. 2010, 76, 3776–3786. [Google Scholar] [CrossRef]
- May, L.A.; Smiley, B.; Schmidt, M.G. Comparative denaturing gradient gel electrophoresis analysis of fungal communities associated with whole plant corn silage. Can. J. Microbiol. 2001, 47, 829–841. [Google Scholar] [CrossRef]
- Salangsang, M.C.D.; Sekine, M.; Akizuki, S.; Sakai, H.D.; Kurosawa, N.; Toda, T. Effect of carbon to nitrogen ratio of food waste and short resting period on microbial accumulation during anaerobic digestion. Biomass Bioenergy 2022, 162, 106481. [Google Scholar] [CrossRef]
- Azis, F.A.; Choo, M.; Suhaimi, H.; Abas, P.E. The Effect of initial carbon to nitrogen ratio on kitchen waste composting maturity. Sustainability 2023, 15, 6191. [Google Scholar] [CrossRef]
- Li, Y.; Liu, S.; Zhao, C.; Zhang, Z.; Nie, D.; Tang, W.; Li, Y. The chemical composition and antibacterial and antioxidant activities of five citrus essential oils. Molecules 2022, 27, 7044. [Google Scholar] [CrossRef] [PubMed]
- Fagbohungbe, M.O.; Herbert, B.M.J.; Hurst, L.; Li, H.; Usmani, S.Q.; Semple, K.T. Impact of biochar on the anaerobic digestion of citrus peel waste. Bioresour. Technol. 2016, 216, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Fang, W.; Liang, J.; Nabi, M.; Cai, Y.; Wang, Q.; Zhang, P.; Zhang, G. Biochar application in anaerobic digestion: Performances, mechanisms, environmental assessment and circular economy. Resour. Conserv. Recycl. 2023, 188, 106720. [Google Scholar] [CrossRef]
- Wu, G.; Healy, M.G.; Zhan, X. Effect of the solid content on anaerobic digestion of meat and bone meal. Bioresour. Technol. 2009, 100, 4326–4331. [Google Scholar] [CrossRef]
- Nges, I.A.; Mbatia, B.; Bjornsson, L. Improved utilization of fish waste by anaerobic digestion following omega-3 fatty acids extraction. J. Environ. Manag. 2012, 110, 159–165. [Google Scholar] [CrossRef]
- Paone, E.; Fazzino, F.; Pizzone, D.M.; Scurria, A.; Pagliaro, M.; Ciriminna, R.; Calabrò, P.S. Towards the anchovy biorefinery: Biogas production from anchovy processing Waste after fish oil extraction with biobased limonene. Sustainability 2021, 13, 2428. [Google Scholar] [CrossRef]
- Agrawal, A.; Chaudhari, P.K.; Ghosh, P. Anaerobic digestion of fruit and vegetable waste: A critical review of associated challenges. Environ. Sci. Pollut. Res. 2023, 30, 24987–25012. [Google Scholar] [CrossRef]
- Sivaramakrishnan, R.; Ramprakash, B.; Ramadoss, G.; Suresh, S.; Pugazhendhi, A.; Incharoensakdi, A. High potential of Rhizopus treated rice bran waste for the nutrient-free anaerobic fermentative biohydrogen production. Bioresour. Technol. 2021, 319, 124193. [Google Scholar] [CrossRef]
- Pan, C.; Fan, Y.; Hou, H. Fermentative production of hydrogen from wheat bran by mixed anaerobic cultures. Ind. Eng. Chem. Res. 2008, 47, 5812–5818. [Google Scholar] [CrossRef]
- Silvestri, S.; Roman, L.M.G.; Alcaraz-González, V.; Carissimi, E. Rice husk to produce both bio-H2 via dark fermentation and photocatalytic hydrochar. Waste Biomass Valor. 2023; published online. [Google Scholar] [CrossRef]
- Tosuner, Z.V.; Taylan, G.G.; Özmıhçı, S. Effects of rice husk particle size on biohydrogen production under solid state fermentation. Int. J. Hydrogen Energy 2019, 44, 18785–18791. [Google Scholar] [CrossRef]
- Arumugam, A.; Sandhya, M.; Ponnusami, V. Biohydrogen and polyhydroxyalkanoate co-production by Enterobacter aerogenes and Rhodobacter sphaeroides from Calophyllum inophyllum oil cake. Bioresour. Technol. 2014, 164, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Vanyan, L.; Cenian, A.; Trchounian, K. Biogas and biohydrogen production using spent coffee grounds and alcohol production waste. Energies 2022, 15, 5935. [Google Scholar] [CrossRef]
- Prajapati, B.R.; Sharma, R.K.; Amatya, I.M. Efect of reduced gas pressure on yield of biogas and other physicochemical parameters. Int. J. Energy Environ. Eng. 2021, 12, 31–37. [Google Scholar] [CrossRef]
- Ghimire, A.; Frunzo, L.; Pirozzi, F.; Trably, E.; Escudie, R.; Lens, P.N.L.; Esposito, G. A review on dark fermentative biohydrogen production from organic biomass: Process parameters and use of by-products. Appl. Energy 2015, 144, 73–95. [Google Scholar] [CrossRef]
- Yin, D.M.; Westerholm, M.; Qiao, W.; Bi, S.J.; Wandera, S.M.; Fan, R.; Jiang, M.M.; Dong, R.J. An explanation of the methanogenic pathway for methane production in anaerobic digestion of nitrogen-rich materials under mesophilic and thermophilic conditions. Bioresour. Technol. 2018, 264, 42–50. [Google Scholar] [CrossRef]
- Buettner, C.; von Bergen, M.; Jehmlich, N.; Noll, M. Pseudomonas spp. are key players in agricultural biogas substrate degradation. Sci. Rep. 2019, 9, 12871. [Google Scholar] [CrossRef]
- Zoelzer, F.; Burger, A.L.; Dierkes, P.W. Unraveling differences in fecal microbiota stability in mammals: From high variable carnivores and consistently stable herbivores. Anim. Microbiome 2021, 3, 77. [Google Scholar] [CrossRef]
- Zeikus, J.G.; Winfrey, M.R. Temperature limitation of methanogenesis in aquatic sediments. Appl. Environ. Microbiol. 1976, 31, 99–107. [Google Scholar] [CrossRef]
- Großkopf, R.; Janssen, P.H.; Liesack, W. Diversity and structure of the methanogenic community in anoxic rice paddy soil microcosms as examined by cultivation and direct 16S rRNA gene sequence retrieval. Appl. Environ. Microbiol. 1998, 64, 960–969. [Google Scholar] [CrossRef] [PubMed]
- Ike, M.; Inoue, D.; Miyano, T.; Liu, T.T.; Sei, K.; Soda, S.; Kadoshin, S. Microbial population dynamics during startup of a full-scale anaerobic digester treating industrial food waste in Kyoto eco-energy project. Bioresour. Technol. 2010, 101, 3952–3957. [Google Scholar] [CrossRef]
- Vítězová, M.; Kohoutová, A.; Vítěz, T.; Hanišáková, N.; Kushkevych, I. Methanogenic microorganisms in industrial wastewater anaerobic treatment. Processes 2020, 8, 1546. [Google Scholar] [CrossRef]
- Park, S.G.; Rhee, C.; Shin, S.G.; Shin, J.; Mohamed, H.O.; Choi, Y.G.; Chae, K.J. Methanogenesis stimulation and inhibition for the production of different target electrobiofuels in microbial electrolysis cells through an on-demand control strategy using the coenzyme M and 2-bromoethanesulfonate. Environ. Int. 2019, 131, 105006. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.B.; de Jonge, N.; Dolriis, M.D.; Kragelund, C.; Fischer, C.H.; Eskesen, M.R.; Noer, K.; Møller, H.B.; Ottosen, L.D.M.; Nielsen, J.L.; et al. Cellulolytic and xylanolytic microbial communities associated with lignocellulose-rich wheat straw degradation in anaerobic digestion. Front. Microbiol. 2021, 12, 645174. [Google Scholar] [CrossRef]
- Zhu, R.; Wang, D.H.; Zheng, Y.; Zou, H.; Fu, S.F. Understanding the mechanisms behind micro-aeration to enhance anaerobic digestion of corn straw. Fuel 2022, 318, 123604. [Google Scholar] [CrossRef]
- Mshandete, A.M.; Björnsson, L.; Kivaisi, A.K.; Rubindamayugi, M.S.T.; Mattiasson, B. Two-stage anaerobic digestion of aerobic pre-treated sisal leaf decortications residues: Hydrolases activities and biogas production profile. Afr. J. Biochem. Res. 2008, 2, 211–218. [Google Scholar]
- Parawira, W.; Murto, M.; Read, J.S.; Mattiasson, B. Profile of hydrolases and biogas production during two-stage mesophilic anaerobic digestion of solid potato waste. Process Biochem. 2005, 40, 2945–2952. [Google Scholar] [CrossRef]
- Gao, S.; Huang, Y.; yang, L.; Wang, H.; Zhao, M.; Xu, Z.; Huang, Z.; Ruan, W. Evaluation the anaerobic digestion performance of solid residual kitchen waste by NaHCO3 buffering. Energy Convers. Manag. 2015, 93, 166–174. [Google Scholar] [CrossRef]
- Kuhn, F.; Berghahn, E.; Sperotto, R.A. Use of biotechnological approaches to add value to rice hulls. Biotechnol. Prog. 2019, 35, e2861. [Google Scholar] [CrossRef]
- Koh, K.H.; Tham, F.Y. Screening of traditional Chinese medicinal plants for quorum-sensing inhibitors activity. J. Microbiol. Immunol. Infect. 2011, 44, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Thomas, L.; Joseph, A.; Gottumukkala, L.D. Xylanase and cellulase systems of Clostridium sp.: An insight on molecular approaches for strain improvement. Bioresour. Technol. 2014, 158, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Reddy, M.P.; Saritha, K.V. Effects of the culture media optimization on pectinase production by Enterobacter sp. PSTB-1. 3 Biotech 2016, 6, 207. [Google Scholar] [CrossRef] [PubMed]
- Mahato, R.K.; Kumar, D.; Rajagopalan, G. Biohydrogen production from fruit waste by Clostridium strain BOH3. Renew. Energy 2020, 153, 1368–1377. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, T.; Zhang, Z.; Tahir, N.; Zhang, Q. Biohydrogen production from Humulus scandens by dark fermentation: Potential evaluation and process optimization. Int. J. Hydrogen Energy 2020, 45, 3760–3768. [Google Scholar] [CrossRef]
- Sarma, M.K.; Ramkumar, N.; Subudhi, S. Biohydrogen production from aquatic plant and algae biomass by Enterobacter cloacae strain DT-1. Chem. Eng. Technol. 2023, 46, 234–241. [Google Scholar] [CrossRef]
- Sukumaran, S.; Raguvaran, K.; Nagarajan, K.; Chinnaperumal, K.; Maheswaran, R.; Chinnasamy, R. Isolation, molecular identification, and characterization of cellulolytic microbes from the gut of grasshopper Eyprepocnemis alacris (Serv. 1838). Biomass Convers. Biorefinery 2023, 1–13. [Google Scholar] [CrossRef]
- Sheng, P.; Li, Y.; Marshall, S.D.G.; Zhang, H. High genetic diversity of microbial cellulase and hemicellulase genes in the hindgut of Holotrichia parallela larvae. Int. J. Mol. Sci. 2015, 16, 16545–16559. [Google Scholar] [CrossRef]
- Contesini, F.J.; Melo, R.R.D.; Sato, H.H. An overview of Bacillus proteases: From production to application. Crit. Rev. Biotechnol. 2018, 38, 321–334. [Google Scholar] [CrossRef]
- Ren, Z.; You, W.; Wu, S.; Poetsch, A.; Xu, C. Secretomic analyses of Ruminiclostridium papyrosolvens reveal its enzymatic basis for lignocellulose degradation. Biotechnol. Biofuels 2019, 12, 183. [Google Scholar] [CrossRef]
- Wang, N.; Yan, Z.; Liu, N.; Zhang, X.; Xu, C. Synergy of cellulase systems between Acetivibrio thermocellus and Thermoclostridium stercorarium in consolidated-bioprocessing for cellulosic ethanol. Microorganisms 2022, 10, 502. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Food Waste | TOC | TN | TOC/TN Ratio | pH |
|---|---|---|---|---|
| Cattle bone | 87.8 | 27.5 | 3.2 | 8.3 |
| Fish bone | 66.3 | 20.9 | 3.2 | 5.8 |
| Carrot peel | 230.7 | 3.8 | 60.7 | 5.3 |
| White radish peel | 230.0 | 16.3 | 14.1 | 5.0 |
| Cabbage stem | 233.5 | 15.6 | 14.9 | 5.4 |
| Lotus stem peel | 236.9 | 10.1 | 23.5 | 6.8 |
| Apple core and peel | 318.6 | 1.9 | 170.8 | 4.8 |
| Orange peel | 311.5 | 7.1 | 44.2 | 4.4 |
| Grape peel | 206.1 | 4.0 | 51.9 | 4.5 |
| Rice hull | 18.4 | 0.5 | 37.3 | 6.6 |
| Rice bran | 147.1 | 13.1 | 11.2 | 6.6 |
| Wheat bran | 104.1 | 6.4 | 16.2 | 6.4 |
| Soy sauce lees | 62.9 | 11.1 | 5.7 | 5.9 |
| Spent bonito flakes | 20.1 | 6.7 | 3.0 | 6.1 |
| Spent dried kelp | 156.9 | 4.2 | 37.5 | 6.5 |
| Spent tea leaves | 103.4 | 6.4 | 16.2 | 5.5 |
| Spent coffee ground | 62.7 | 5.0 | 12.6 | 5.3 |
| Rapeseed oil cake | 96.0 | 5.6 | 17.0 | 5.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasaka, S.; Sakamoto, S.; Fujii, K. The Potential of Digested Sludge-Assimilating Microflora for Biogas Production from Food Processing Wastes. Microorganisms 2023, 11, 2321. https://doi.org/10.3390/microorganisms11092321
Hasaka S, Sakamoto S, Fujii K. The Potential of Digested Sludge-Assimilating Microflora for Biogas Production from Food Processing Wastes. Microorganisms. 2023; 11(9):2321. https://doi.org/10.3390/microorganisms11092321
Chicago/Turabian StyleHasaka, Sato, Saki Sakamoto, and Katsuhiko Fujii. 2023. "The Potential of Digested Sludge-Assimilating Microflora for Biogas Production from Food Processing Wastes" Microorganisms 11, no. 9: 2321. https://doi.org/10.3390/microorganisms11092321
APA StyleHasaka, S., Sakamoto, S., & Fujii, K. (2023). The Potential of Digested Sludge-Assimilating Microflora for Biogas Production from Food Processing Wastes. Microorganisms, 11(9), 2321. https://doi.org/10.3390/microorganisms11092321