
The Gut-Wrenching Effects of Cryptosporidiosis and Giardiasis in Children

Abstract
:1. Introduction
2. The Causative Parasites
2.1. Cryptosporidium Species
2.2. Giardia duodenalis
3. Transmission and Prevalence
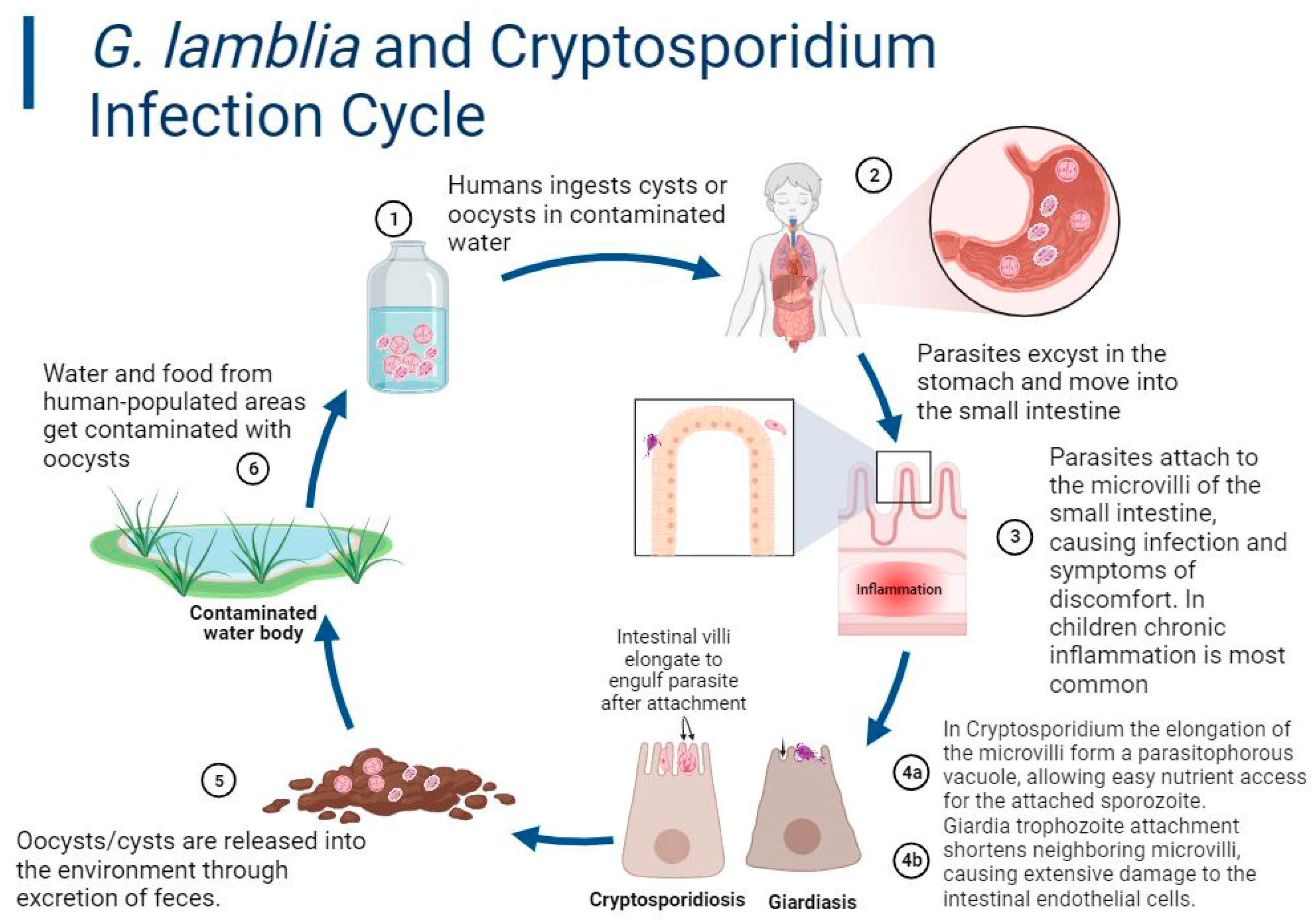
4. Pathogenesis and Clinical Presentation
4.1. Cryptosporidiosis
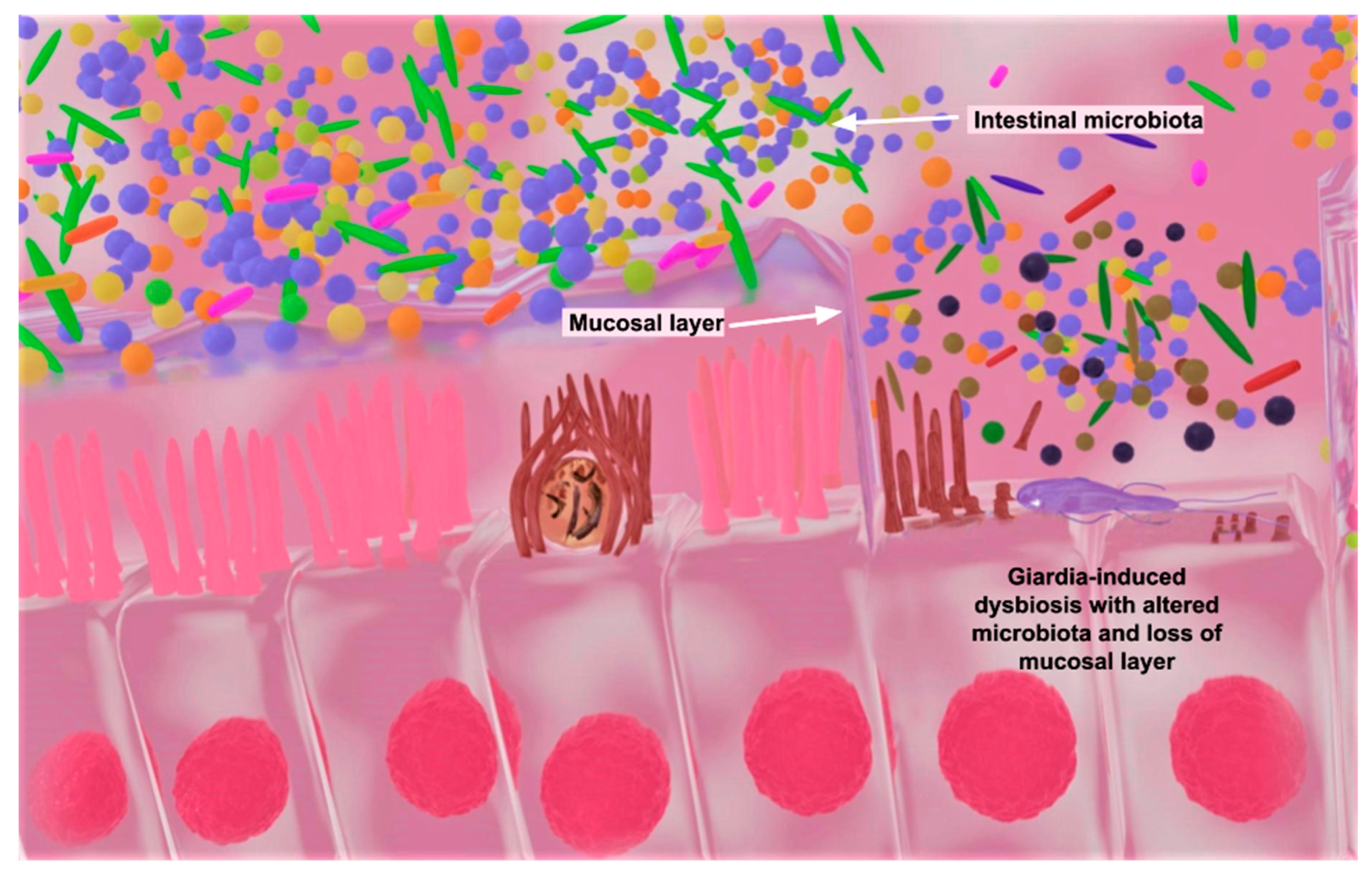
4.2. Giardiasis
5. Symptoms and Clinical Manifestation
6. Diagnosis
7. Current Therapy for Cryptosporidiosis and Giardiasis

{kind=link}
{kind=link}
| Drug | Recommended Dose for Children | Duration of Treatment | Side Effects | References |
|---|---|---|---|---|
| Nitazoxanide * | 100–200 mg per 12 h | 3 days | Hives; difficulty breathing; swelling of throat, tongue, eyes, and face; headache; stomachache; nausea; discolored urine. | [147] |
| Metronidazole # | 15 mg/kg per day | 3 times a day for 5 days | Dizziness; headache; vomiting/nausea; diarrhea; stomach cramps; loss of appetite; dry mouth and metallic taste in mouth. | [148] |
| Tinidazole # | 50 mg/kg per day | 1 dose | Nausea/vomiting; diarrhea; bitter or metallic taste in mouth; abdominal pain. | [149,150] |
| Secnidazole # | 30 mg/kg | 1 or 2 doses | Gastrointestinal tract disorders similar to Tinidazole. | [151] |
| Albendazole # | 10 mg/kg per day | 5 days | Abdominal pain; nausea/vomiting; headache. | [152] |
| Mebendazole # | 100 mg | 2 times a day for 3 days | Loss of appetite; abdominal pain; diarrhea, flatulence; nausea/vomiting; headache; tinnitus. | [153] |
8. Conclusions and Future Outlook
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Abbafati, C.; Abbas, K.M.; Abbasi, M.; Abbasifard, M.; Abbasi-Kangevari, M.; Abbastabar, H.; Abd-Allah, F.; Abdelalim, A.; Abdollahi, M.; Abdollahpour, I.; et al. Global burden of 369 diseases and injuries in 204 countries and territories, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [Google Scholar]
- Lanata, C.F.; Fischer-Walker, C.L.; Olascoaga, A.C.; Torres, C.X.; Aryee, M.J.; Black, R.E.; Child Health Epidemiology Reference Group of the World Health Organization and UNICEF. Global causes of diarrheal disease mortality in children <5 years of age: A systematic review. PLoS ONE 2013, 8, e72788. [Google Scholar] [CrossRef]
- Okeke, I.N.; Nataro, J.P. Enteroaggregative Escherichia coli. Lancet Infect. Dis. 2001, 1, 304–313. [Google Scholar] [CrossRef]
- Bruzzese, E.; Giannattasio, A.; Guarino, A. Antibiotic treatment of acute gastroenteritis in children. F1000Research 2018, 7, 193. [Google Scholar] [CrossRef]
- Kotloff, K.L. Bacterial diarrhoea. Curr. Opin. Pediatr. 2022, 34, 147–155. [Google Scholar] [CrossRef]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef]
- Parashar, U.D.; Nelson, E.A.; Kang, G. Diagnosis, management, and prevention of rotavirus gastroenteritis in children. BMJ 2013, 347, f7204. [Google Scholar] [CrossRef]
- Burnett, E.; Jonesteller, C.L.; Tate, J.E.; Yen, C.; Parashar, U.D. Global Impact of Rotavirus Vaccination on Childhood Hospitalizations and Mortality From Diarrhea. J. Infect. Dis. 2017, 215, 1666–1672. [Google Scholar] [CrossRef]
- Donowitz, J.R.; Alam, M.; Kabir, M.; Ma, J.Z.; Nazib, F.; Platts-Mills, J.A.; Bartelt, L.A.; Haque, R.; Petri, W.A., Jr. A Prospective Longitudinal Cohort to Investigate the Effects of Early Life Giardiasis on Growth and All Cause Diarrhea. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2016, 63, 792–797. [Google Scholar] [CrossRef]
- Platts-Mills, J.A.; Babji, S.; Bodhidatta, L.; Gratz, J.; Haque, R.; Havt, A.; McCormick, B.J.; McGrath, M.; Olortegui, M.P.; Samie, A.; et al. Pathogen-specific burdens of community diarrhoea in developing countries: A multisite birth cohort study (MAL-ED). Lancet Glob. Health 2015, 3, e564–e575. [Google Scholar] [CrossRef]
- Hajissa, K.; Islam, M.A.; Sanyang, A.M.; Mohamed, Z. Prevalence of intestinal protozoan parasites among school children in africa: A systematic review and meta-analysis. PLoS Neglected Trop. Dis. 2022, 16, e0009971. [Google Scholar] [CrossRef]
- Kotloff, K.L.; Nataro, J.P.; Blackwelder, W.C.; Nasrin, D.; Farag, T.H.; Panchalingam, S.; Wu, Y.; Sow, S.O.; Sur, D.; Breiman, R.F.; et al. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): A prospective, case-control study. Lancet 2013, 382, 209–222. [Google Scholar] [CrossRef]
- Marcenac, P.; Traore, A.; Kim, S.; Prentice-Mott, G.; Berendes, D.M.; Powell, H.; Kasumba, I.N.; Nasrin, D.; Jones, J.C.M.; Zaman, S.M.A.; et al. Giardia Detection and Codetection With Other Enteric Pathogens in Young Children in the Vaccine Impact on Diarrhea in Africa (VIDA) Case-Control Study: 2015–2018. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2023, 76, S106–S113. [Google Scholar] [CrossRef]
- Operario, D.J.; Platts-Mills, J.A.; Nadan, S.; Page, N.; Seheri, M.; Mphahlele, J.; Praharaj, I.; Kang, G.; Araujo, I.T.; Leite, J.P.G.; et al. Etiology of Severe Acute Watery Diarrhea in Children in the Global Rotavirus Surveillance Network Using Quantitative Polymerase Chain Reaction. J. Infect. Dis. 2017, 216, 220–227. [Google Scholar] [CrossRef]
- Abdel-Messih, I.A.; Wierzba, T.F.; Abu-Elyazeed, R.; Ibrahim, A.F.; Ahmed, S.F.; Kamal, K.; Sanders, J.; Frenck, R. Diarrhea associated with Cryptosporidium parvum among young children of the Nile River Delta in Egypt. J. Trop. Pediatr. 2005, 51, 154–159. [Google Scholar] [CrossRef]
- Gatei, W.; Wamae, C.N.; Mbae, C.; Waruru, A.; Mulinge, E.; Waithera, T.; Gatika, S.M.; Kamwati, S.K.; Revathi, G.; Hart, C.A. Cryptosporidiosis: Prevalence, genotype analysis, and symptoms associated with infections in children in Kenya. Am. J. Trop. Med. Hyg. 2006, 75, 78–82. [Google Scholar] [CrossRef]
- Garzon, M.; Pereira-da-Silva, L.; Seixas, J.; Papoila, A.L.; Alves, M. Subclinical Enteric Parasitic Infections and Growth Faltering in Infants in Sao Tome, Africa: A Birth Cohort Study. Int. J. Environ. Res. Public Health 2018, 15, 688. [Google Scholar] [CrossRef]
- White, A.C. Cryptosporidium Species. In Mandell, Douglas, and Bennett’s Principles and Practice of Infectious Diseases, 8th ed.; Bennett, J.E., Dolin, R., Blaser, M.J., Eds.; Elsevier/Saunders: Philadelphia, PA, USA, 2015. [Google Scholar]
- Ryan, U.M.; Feng, Y.; Fayer, R.; Xiao, L. Taxonomy and molecular epidemiology of Cryptosporidium and Giardia—A 50 year perspective (1971–2021). Int. J. Parasitol. 2021, 51, 1099–1119. [Google Scholar] [CrossRef]
- Cunha, F.S.; Peralta, R.H.S.; Peralta, J.M. New insights into the detection and molecular characterization of Cryptosporidium with emphasis in Brazilian studies: A review. Rev. Inst. Med. Trop. Sao Paulo 2019, 61, e28. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Feng, Y. Molecular epidemiologic tools for waterborne pathogens Cryptosporidium spp. and Giardia duodenalis. Food Waterborne Parasitol. 2017, 8–9, 14–32. [Google Scholar] [CrossRef]
- Chappell, C.L.; Okhuysen, P.C.; Langer-Curry, R.C.; Akiyoshi, D.E.; Widmer, G.; Tzipori, S. Cryptosporidium meleagridis: Infectivity in healthy adult volunteers. Am. J. Trop. Med. Hyg. 2011, 85, 238–242. [Google Scholar] [CrossRef]
- Beser, J.; Toresson, L.; Eitrem, R.; Troell, K.; Winiecka-Krusnell, J.; Lebbad, M. Possible zoonotic transmission of Cryptosporidium felis in a household. Infect. Ecol. Epidemiol. 2015, 5, 28463. [Google Scholar] [CrossRef]
- Thomson, S.; Hamilton, C.A.; Hope, J.C.; Katzer, F.; Mabbott, N.A.; Morrison, L.J.; Innes, E.A. Bovine cryptosporidiosis: Impact, host-parasite interaction and control strategies. Vet. Res. 2017, 48, 42. [Google Scholar] [CrossRef]
- Tyzzer, E. Cryptosporidium parvum (sp. nov.), a coccidium found in the small intestine of the common mouse. Arch. Für Protistenkd. 1912, 26, 394–418. [Google Scholar]
- Cama, V.A.; Bern, C.; Roberts, J.; Cabrera, L.; Sterling, C.R.; Ortega, Y.; Gilman, R.H.; Xiao, L. Cryptosporidium species and subtypes and clinical manifestations in children, Peru. Emerg. Infect. Dis. 2008, 14, 1567–1574. [Google Scholar] [CrossRef]
- Bouzid, M.; Hunter, P.R.; Chalmers, R.M.; Tyler, K.M. Cryptosporidium pathogenicity and virulence. Clin. Microbiol. Rev. 2013, 26, 115–134. [Google Scholar] [CrossRef]
- Current, W.L.; Garcia, L.S. Cryptosporidiosis. Clin. Microbiol. Rev. 1991, 4, 325–358. [Google Scholar] [CrossRef]
- Dhal, A.K.; Panda, C.; Yun, S.I.; Mahapatra, R.K. An update on Cryptosporidium biology and therapeutic avenues. J. Parasit. Dis. 2022, 46, 923–939. [Google Scholar] [CrossRef]
- Boulter-Bitzer, J.I.; Lee, H.; Trevors, J.T. Molecular targets for detection and immunotherapy in Cryptosporidium parvum. Biotechnol. Adv. 2007, 25, 13–44. [Google Scholar] [CrossRef]
- Ryan, U.; Zahedi, A. Molecular epidemiology of giardiasis from a veterinary perspective. Adv. Parasitol. 2019, 106, 209–254. [Google Scholar] [CrossRef]
- Feng, Y.; Xiao, L. Zoonotic potential and molecular epidemiology of Giardia species and giardiasis. Clin. Microbiol. Rev. 2011, 24, 110–140. [Google Scholar] [CrossRef]
- Simner, P.J. Medical Parasitology Taxonomy Update: January 2012 to December 2015. J. Clin. Microbiol. 2017, 55, 43–47. [Google Scholar] [CrossRef]
- Monis, P.T.; Thompson, R.C. Cryptosporidium and Giardia-zoonoses: Fact or fiction? Infect. Genet. Evol. 2003, 3, 233–244. [Google Scholar] [CrossRef]
- Michaels, S.A.; Hulverson, M.A.; Whitman, G.R.; Tran, L.T.; Choi, R.; Fan, E.; McNamara, C.W.; Love, M.S.; Ojo, K.K. Repurposing the Kinase Inhibitor Mavelertinib for Giardiasis Therapy. Antimicrob. Agents Chemother. 2022, 66, e0001722. [Google Scholar] [CrossRef]
- Seabolt, M.H.; Roellig, D.M.; Konstantinidis, K.T. Genomic comparisons confirm Giardia duodenalis sub-assemblage AII as a unique species. Front. Cell. Infect. Microbiol. 2022, 12, 1010244. [Google Scholar] [CrossRef]
- Adam, R.D. Giardia duodenalis: Biology and Pathogenesis. Clin. Microbiol. Rev. 2021, 34, e0002419. [Google Scholar] [CrossRef]
- Broglia, A.; Weitzel, T.; Harms, G.; Caccio, S.M.; Nockler, K. Molecular typing of Giardia duodenalis isolates from German travellers. Parasitol. Res. 2013, 112, 3449–3456. [Google Scholar] [CrossRef]
- Soliman, R.H.; Fuentes, I.; Rubio, J.M. Identification of a novel Assemblage B subgenotype and a zoonotic Assemblage C in human isolates of Giardia intestinalis in Egypt. Parasitol. Int. 2011, 60, 507–511. [Google Scholar] [CrossRef]
- Heyworth, M.F. Giardia duodenalis genetic assemblages and hosts. Parasite 2016, 23, 13. [Google Scholar] [CrossRef]
- Calder, P.C.; Jackson, A.A. Undernutrition, infection and immune function. Nutr. Res. Rev. 2000, 13, 3–29. [Google Scholar] [CrossRef]
- Astiazaran-Garcia, H.; Inigo-Figueroa, G.; Quihui-Cota, L.; Anduro-Corona, I. Crosstalk between Zinc Status and Giardia Infection: A New Approach. Nutrients 2015, 7, 4438–4452. [Google Scholar] [CrossRef] [PubMed]
- Khalil, I.A.; Troeger, C.; Rao, P.C.; Blacker, B.F.; Brown, A.; Brewer, T.G.; Colombara, D.V.; De Hostos, E.L.; Engmann, C.; Guerrant, R.L.; et al. Morbidity, mortality, and long-term consequences associated with diarrhoea from Cryptosporidium infection in children younger than 5 years: A meta-analyses study. Lancet Glob. Health 2018, 6, e758–e768. [Google Scholar] [CrossRef] [PubMed]
- Squire, S.A.; Ryan, U. Cryptosporidium and Giardia in Africa: Current and future challenges. Parasites Vectors 2017, 10, 195. [Google Scholar] [CrossRef] [PubMed]
- Pires, S.M.; Fischer-Walker, C.L.; Lanata, C.F.; Devleesschauwer, B.; Hall, A.J.; Kirk, M.D.; Duarte, A.S.; Black, R.E.; Angulo, F.J. Aetiology-Specific Estimates of the Global and Regional Incidence and Mortality of Diarrhoeal Diseases Commonly Transmitted through Food. PLoS ONE 2015, 10, e0142927. [Google Scholar] [CrossRef] [PubMed]
- Sow, S.O.; Muhsen, K.; Nasrin, D.; Blackwelder, W.C.; Wu, Y.; Farag, T.H.; Panchalingam, S.; Sur, D.; Zaidi, A.K.; Faruque, A.S.; et al. The Burden of Cryptosporidium Diarrheal Disease among Children <24 Months of Age in Moderate/High Mortality Regions of Sub-Saharan Africa and South Asia, Utilizing Data from the Global Enteric Multicenter Study (GEMS). PLoS Neglected Trop. Dis. 2016, 10, e0004729. [Google Scholar] [CrossRef]
- Gilbert, I.H.; Vinayak, S.; Striepen, B.; Manjunatha, U.H.; Khalil, I.A.; Van Voorhis, W.C. Safe and effective treatments are needed for cryptosporidiosis, a truly neglected tropical disease. BMJ Glob. Health 2023, 8, e012540. [Google Scholar] [CrossRef]
- Hajare, S.T.; Chekol, Y.; Chauhan, N.M. Assessment of prevalence of Giardia lamblia infection and its associated factors among government elementary school children from Sidama zone, SNNPR, Ethiopia. PLoS ONE 2022, 17, e0264812. [Google Scholar] [CrossRef]
- Todd, E. Foodborne Diseases: Overview of Biological Hazards and Foodborne Diseases; Motarjemi, Y., Ed.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 1, pp. 221–242. [Google Scholar]
- Busatti, H.G.; Santos, J.F.; Gomes, M.A. The old and new therapeutic approaches to the treatment of giardiasis: Where are we? Biologics 2009, 3, 273–287. [Google Scholar]
- DuPont, H.L.; Chappell, C.L.; Sterling, C.R.; Okhuysen, P.C.; Rose, J.B.; Jakubowski, W. The infectivity of Cryptosporidium parvum in healthy volunteers. N. Engl. J. Med. 1995, 332, 855–859. [Google Scholar] [CrossRef]
- Okhuysen, P.C.; Chappell, C.L.; Crabb, J.H.; Sterling, C.R.; DuPont, H.L. Virulence of three distinct Cryptosporidium parvum isolates for healthy adults. J. Infect. Dis. 1999, 180, 1275–1281. [Google Scholar] [CrossRef] [PubMed]
- Chappell, C.L.; Okhuysen, P.C.; Langer-Curry, R.; Widmer, G.; Akiyoshi, D.E.; Tanriverdi, S.; Tzipori, S. Cryptosporidium hominis: Experimental challenge of healthy adults. Am. J. Trop. Med. Hyg. 2006, 75, 851–857. [Google Scholar] [CrossRef]
- Korpe, P.; Ni, Z.M.; Kabir, M.; Alam, M.; Ferdous, T.; Ara, R.; Munday, R.M.; Haque, R.; Duggal, P. Prospective Cohort Study of Cryptosporidium Infection and Shedding in Infants and Their Households. Clin. Infect. Dis. 2023, 76, 2178–2186. [Google Scholar] [CrossRef]
- Adam, R.D. Biology of Giardia lamblia. Clin. Microbiol. Rev. 2001, 14, 447–475. [Google Scholar] [CrossRef] [PubMed]
- Dunn, N.; Juergens, A.L. Giardiasis. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Sarkar, R.; Ajjampur, S.S.R.; Prabakaran, A.D.; Geetha, J.C.; Sowmyanarayanan, T.V.; Kane, A.; Duara, J.; Muliyil, J.; Balraj, V.; Naumova, E.N.; et al. Cryptosporidiosis Among Children in an Endemic Semiurban Community in Southern India: Does a Protected Drinking Water Source Decrease Infection? Clin. Infect. Dis. 2013, 57, 398–406. [Google Scholar] [CrossRef] [PubMed]
- D’Antonio, R.G.; Winn, R.E.; Taylor, J.P.; Gustafson, T.L.; Current, W.L.; Rhodes, M.M.; Gary, G.W., Jr.; Zajac, R.A. A waterborne outbreak of cryptosporidiosis in normal hosts. Ann. Intern. Med. 1985, 103, 886–888. [Google Scholar] [CrossRef] [PubMed]
- Graczyk, T.K.; Fayer, R.; Cranfield, M.R. Zoonotic transmission of Cryptosporidium parvum: Implications for water-borne cryptosporidiosis. Parasitol. Today 1997, 13, 348–351. [Google Scholar] [CrossRef] [PubMed]
- Mac Kenzie, W.R.; Hoxie, N.J.; Proctor, M.E.; Gradus, M.S.; Blair, K.A.; Peterson, D.E.; Kazmierczak, J.J.; Addiss, D.G.; Fox, K.R.; Rose, J.B. A massive outbreak in Milwaukee of Cryptosporidium infection transmitted through the public water supply. N. Engl. J. Med. 1994, 331, 161–167. [Google Scholar] [CrossRef]
- Escobedo, A.A.; Almirall, P.; Robertson, L.J.; Franco, R.M.; Hanevik, K.; Morch, K.; Cimerman, S. Giardiasis: The ever-present threat of a neglected disease. Infect. Disord. Drug Targets 2010, 10, 329–348. [Google Scholar] [CrossRef]
- Tzipori, S. Cryptosporidiosis in perspective. Adv. Parasitol. 1988, 27, 63–129. [Google Scholar]
- Bamaiyi, P.H.; Redhuan, N.E.M. Prevalence and risk factors for cryptosporidiosis: A global, emerging, neglected zoonosis. Asian Biomed. 2016, 10, 309–325. [Google Scholar] [CrossRef]
- Suolaniemi, J.; Autio, T.; Heikkinen, J.; Rasanen, K. Knowledge, Attitudes, and Practices of Finnish Dairy Farmers on Cryptosporidiosis. J. Agromed. 2023, 28, 288–299. [Google Scholar] [CrossRef]
- Rumsey, P.; Waseem, M. Giardia lamblia Enteritis. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Bowman, D.D.; Lucio-Forster, A. Cryptosporidiosis and giardiasis in dogs and cats: Veterinary and public health importance. Exp. Parasitol. 2010, 124, 121–127. [Google Scholar] [CrossRef]
- Cai, W.; Ryan, U.; Xiao, L.; Feng, Y. Zoonotic giardiasis: An update. Parasitol. Res. 2021, 120, 4199–4218. [Google Scholar] [CrossRef]
- Winter, G.; Gooley, A.A.; Williams, K.L.; Slade, M.B. Characterization of a major sporozoite surface glycoprotein of Cryptosporidum parvum. Funct. Integr. Genom. 2000, 1, 207–217. [Google Scholar] [CrossRef]
- Cui, Z.H.; Wang, L.Y.; Wang, Y.X.; Li, J.; Wang, R.J.; Sun, M.F.; Zhang, L.X. Cryptosporidium parvum gp40/15 Is Associated with the Parasitophorous Vacuole Membrane and Is a Potential Vaccine Target. Microorganisms 2020, 8, 363. [Google Scholar] [CrossRef]
- Tzipori, S.; Ward, H. Cryptosporidiosis: Biology, pathogenesis and disease. Microbes Infect. 2002, 4, 1047–1058. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.M.; Keithly, J.S.; Paya, C.V.; LaRusso, N.F. Cryptosporidiosis. N. Engl. J. Med. 2002, 346, 1723–1731. [Google Scholar] [CrossRef]
- Borad, A.; Ward, H. Human immune responses in cryptosporidiosis. Future Microbiol. 2010, 5, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Leitch, G.J.; He, Q. Cryptosporidiosis-an overview. J. Biomed. Res. 2012, 25, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Lumadue, J.A.; Manabe, Y.C.; Moore, R.D.; Belitsos, P.C.; Sears, C.L.; Clark, D.P. A clinicopathologic analysis of AIDS-related cryptosporidiosis. AIDS 1998, 12, 2459–2466. [Google Scholar] [CrossRef]
- Ojcius, D.M.; Perfettini, J.L.; Bonnin, A.; Laurent, F. Caspase-dependent apoptosis during infection with Cryptosporidium parvum. Microbes Infect. 1999, 1, 1163–1168. [Google Scholar] [CrossRef]
- Kelly, P.; Jack, D.L.; Naeem, A.; Mandanda, B.; Pollok, R.C.; Klein, N.J.; Turner, M.W.; Farthing, M.J. Mannose-binding lectin is a component of innate mucosal defense against Cryptosporidium parvum in AIDS. Gastroenterology 2000, 119, 1236–1242. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, B.; Huston, C.; Wagner, D.; Noel, F.; Rouzier, P.; Pape, J.; Bois, G.; Larsson, C.; Alston, W.; Tenney, K. Serum mannose-binding lectin deficiency is associated with cryptosporidiosis in young Haitian children. Clin. Infect. Dis. 2006, 43, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Carmolli, M.; Duggal, P.; Haque, R.; Lindow, J.; Mondal, D.; Petri, W.A., Jr.; Mourningstar, P.; Larsson, C.J.; Sreenivasan, M.; Khan, S.; et al. Deficient serum mannose-binding lectin levels and MBL2 polymorphisms increase the risk of single and recurrent Cryptosporidium infections in young children. J. Infect. Dis. 2009, 200, 1540–1547. [Google Scholar] [CrossRef]
- Mead, J.R. Early immune and host cell responses to Cryptosporidium infection. Front. Parasitol. 2023, 2, 1113950. [Google Scholar] [CrossRef]
- Kirkpatrick, B.D.; Noel, F.; Rouzier, P.D.; Powell, J.L.; Pape, J.W.; Bois, G.; Alston, W.K.; Larsson, C.J.; Tenney, K.; Ventrone, C.; et al. Childhood cryptosporidiosis is associated with a persistent systemic inflammatory response. Clin. Infect. Dis. 2006, 43, 604–608. [Google Scholar] [CrossRef] [PubMed]
- Ballinger, A. Fundamental mechanisms of growth failure in inflammatory bowel disease. Horm. Res. Paediatr. 2002, 58 (Suppl. 1), 7–10. [Google Scholar] [CrossRef]
- Amat, C.B.; Motta, J.P.; Fekete, E.; Moreau, F.; Chadee, K.; Buret, A.G. Cysteine Protease-Dependent Mucous Disruptions and Differential Mucin Gene Expression in Giardia duodenalis Infection. Am. J. Pathol. 2017, 187, 2486–2498. [Google Scholar] [CrossRef]
- Connaris, S.; Greenwell, P. Glycosidases in mucin-dwelling protozoans. Glycoconj. J. 1997, 14, 879–882. [Google Scholar] [CrossRef]
- Otani, T.; Furuse, M. Tight Junction Structure and Function Revisited. Trends Cell Biol. 2020, 30, 805–817. [Google Scholar] [CrossRef]
- Solaymani-Mohammadi, S. Mucosal Defense Against Giardia at the Intestinal Epithelial Cell Interface. Front. Immunol. 2022, 13, 817468. [Google Scholar] [CrossRef]
- Halliez, M.C.; Motta, J.P.; Feener, T.D.; Guerin, G.; LeGoff, L.; Francois, A.; Colasse, E.; Favennec, L.; Gargala, G.; Lapointe, T.K.; et al. Giardia duodenalis induces paracellular bacterial translocation and causes postinfectious visceral hypersensitivity. Am. J. Physiol. Liver Physiol. 2016, 310, G574–G585. [Google Scholar] [CrossRef]
- Barash, N.R.; Maloney, J.G.; Singer, S.M.; Dawson, S.C. Giardia Alters Commensal Microbial Diversity throughout the Murine Gut. Infect. Immun. 2017, 85, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Laurent, F.; Eckmann, L.; Savidge, T.C.; Morgan, G.; Theodos, C.; Naciri, M.; Kagnoff, M.F. Cryptosporidium parvum infection of human intestinal epithelial cells induces the polarized secretion of C-X-C chemokines. Infect. Immun. 1997, 65, 5067–5073. [Google Scholar] [CrossRef]
- Circu, M.L.; Aw, T.Y. Intestinal redox biology and oxidative stress. Semin. Cell Dev. Biol. 2012, 23, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Zenobia, C.; Hajishengallis, G. Basic biology and role of interleukin-17 in immunity and inflammation. Periodontol. 2000 2015, 69, 142–159. [Google Scholar] [CrossRef] [PubMed]
- Fekete, E.; Allain, T.; Siddiq, A.; Sosnowski, O.; Buret, A.G. Giardia spp. and the Gut Microbiota: Dangerous Liaisons. Front. Microbiol. 2020, 11, 618106. [Google Scholar] [CrossRef]
- Halliez, M.C.; Buret, A.G. Extra-intestinal and long term consequences of Giardia duodenalis infections. World J. Gastroenterol. WJG 2013, 19, 8974–8985. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, S.M.; Stark, D.; Harkness, J.; Ellis, J. Enteric protozoa in the developed world: A public health perspective. Clin. Microbiol. Rev. 2012, 25, 420–449. [Google Scholar] [CrossRef]
- Checkley, W.; White, A.C., Jr.; Jaganath, D.; Arrowood, M.J.; Chalmers, R.M.; Chen, X.M.; Fayer, R.; Griffiths, J.K.; Guerrant, R.L.; Hedstrom, L.; et al. A review of the global burden, novel diagnostics, therapeutics, and vaccine targets for Cryptosporidium. Lancet Infect. Dis. 2015, 15, 85–94. [Google Scholar] [CrossRef]
- Weber-Stiehl, S.; Jarke, L.; Castrillon-Betancur, J.C.; Gilbert, F.; Sommer, F. Mitochondrial Function and Microbial Metabolites as Central Regulators of Intestinal Immune Responses and Cancer. Front. Microbiol. 2022, 13, 919424. [Google Scholar] [CrossRef] [PubMed]
- Hakim, H.; Dallas, R.; Wolf, J.; Tang, L.; Schultz-Cherry, S.; Darling, V.; Johnson, C.; Karlsson, E.A.; Chang, T.C.; Jeha, S.; et al. Gut Microbiome Composition Predicts Infection Risk During Chemotherapy in Children With Acute Lymphoblastic Leukemia. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2018, 67, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.K.; Kim, M.; Bakshi, U.; Cunningham, K.Y.; Davis, J.M., 3rd; Lazaridis, K.N.; Nelson, H.; Chia, N.; Sung, J. A predictive index for health status using species-level gut microbiome profiling. Nat. Commun. 2020, 11, 4635. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.C.; Olson, M.E.; Zhu, G.; Enomoto, S.; Abrahamsen, M.S.; Hijjawi, N.S. Cryptosporidium and cryptosporidiosis. Adv. Parasitol. 2005, 59, 77–158. [Google Scholar] [CrossRef]
- Feasey, N.A.; Healey, P.; Gordon, M.A. Review article: The aetiology, investigation and management of diarrhoea in the HIV-positive patient. Aliment. Pharmacol. Ther. 2011, 34, 587–603. [Google Scholar] [CrossRef]
- O’Connor, R.M.; Shaffie, R.; Kang, G.; Ward, H.D. Cryptosporidiosis in patients with HIV/AIDS. AIDS 2011, 25, 549–560. [Google Scholar] [CrossRef]
- Shirley, D.A.; Moonah, S.N.; Kotloff, K.L. Burden of disease from cryptosporidiosis. Curr. Opin. Infect. Dis. 2012, 25, 555–563. [Google Scholar] [CrossRef]
- Toro-Londono, M.A.; Bedoya-Urrego, K.; Garcia-Montoya, G.M.; Galvan-Diaz, A.L.; Alzate, J.F. Intestinal parasitic infection alters bacterial gut microbiota in children. PeerJ 2019, 7, e6200. [Google Scholar] [CrossRef]
- Wegayehu, T.; Adamu, H.; Petros, B. Prevalence of Giardia duodenalis and Cryptosporidium species infections among children and cattle in North Shewa Zone, Ethiopia. MC Infect. Dis. 2013, 13, 419. [Google Scholar] [CrossRef]
- Han, M.Y.; Xiao, S.M.; An, W.; Sang, C.H.; Li, H.Y.; Ma, J.F.; Yang, M. Co-infection risk assessment of Giardia and Cryptosporidium with HIV considering synergistic effects and age sensitivity using disability-adjusted life years. Water Res. 2020, 175, 115698. [Google Scholar] [CrossRef]
- Gebre, B.; Alemayehu, T.; Girma, M.; Ayalew, F.; Tadesse, B.T.; Shemelis, T. Cryptosporidiosis And Other Intestinal Parasitic Infections And Concomitant Threats Among HIV-Infected Children In Southern Ethiopia Receiving First-Line Antiretroviral Therapy. HIV/AIDS Res. Palliat. Care 2019, 11, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Muhsen, K.; Levine, M.M. A systematic review and meta-analysis of the association between Giardia lamblia and endemic pediatric diarrhea in developing countries. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2012, 55 (Suppl. 4), S271–S293. [Google Scholar] [CrossRef] [PubMed]
- Michaels, S.A.; Hennessey, K.M.; Paragas, N.; Paredez, A.R.; Ojo, K.K. A Curious Case for Development of Kinase Inhibitors as Antigiardiasis Treatments Using Advanced Drug Techniques. ACS Infect. Dis. 2021, 7, 943–947. [Google Scholar] [CrossRef] [PubMed]
- Dann, S.M.; Le, C.H.Y.; Hanson, E.M.; Ross, M.C.; Eckmann, L. Giardia Infection of the Small Intestine Induces Chronic Colitis in Genetically Susceptible Hosts. J. Immunol. 2018, 201, 548–559. [Google Scholar] [CrossRef]
- Litleskare, S.; Rortveit, G.; Eide, G.E.; Hanevik, K.; Langeland, N.; Wensaas, K.A. Prevalence of Irritable Bowel Syndrome and Chronic Fatigue 10 Years After Giardia Infection. Clin. Gastroenterol. Hepatol. 2018, 16, 1064–1072.e4. [Google Scholar] [CrossRef]
- Di Prisco, M.C.; Hagel, I.; Lynch, N.R.; Jimenez, J.C.; Rojas, R.; Gil, M.; Mata, E. Association between giardiasis and allergy. Ann. Allergy Asthma Immunol. 1998, 81, 261–265. [Google Scholar] [CrossRef]
- Rogawski, E.T.; Bartelt, L.A.; Platts-Mills, J.A.; Seidman, J.C.; Samie, A.; Havt, A.; Babji, S.; Trigoso, D.R.; Qureshi, S.; Shakoor, S.; et al. Determinants and Impact of Giardia Infection in the First 2 Years of Life in the MAL-ED Birth Cohort. J. Pediatr. Infect. Dis. Soc. 2017, 6, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Nash, T.E.; Herrington, D.A.; Losonsky, G.A.; Levine, M.M. Experimental human infections with Giardia lamblia. J. Infect. Dis. 1987, 156, 974–984. [Google Scholar] [CrossRef]
- Homan, W.L.; Mank, T.G. Human giardiasis: Genotype linked differences in clinical symptomatology. Int. J. Parasitol. 2001, 31, 822–826. [Google Scholar] [CrossRef]
- Wang, Y.; Gonzalez-Moreno, O.; Roellig, D.M.; Oliver, L.; Huguet, J.; Guo, Y.; Feng, Y.; Xiao, L. Epidemiological distribution of genotypes of Giardia duodenalis in humans in Spain. Parasites Vectors 2019, 12, 432. [Google Scholar] [CrossRef]
- Chin, A.C.; Teoh, D.A.; Scott, K.G.; Meddings, J.B.; Macnaughton, W.K.; Buret, A.G. Strain-dependent induction of enterocyte apoptosis by Giardia lamblia disrupts epithelial barrier function in a caspase-3-dependent manner. Infect. Immun. 2002, 70, 3673–3680. [Google Scholar] [CrossRef] [PubMed]
- Davids, B.J.; Liu, C.M.; Hanson, E.M.; Le, C.H.Y.; Ang, J.; Hanevik, K.; Fischer, M.; Radunovic, M.; Langeland, N.; Ferella, M.; et al. Identification of Conserved Candidate Vaccine Antigens in the Surface Proteome of Giardia lamblia. Infect. Immun. 2019, 87, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Faubert, G. Immune response to Giardia duodenalis. Clin. Microbiol. Rev. 2000, 13, 35–54. [Google Scholar] [CrossRef]
- Chan, R.; Chen, J.; York, M.K.; Setijono, N.; Kaplan, R.L.; Graham, F.; Tanowitz, H.B. Evaluation of a combination rapid immunoassay for detection of Giardia and Cryptosporidium antigens. J. Clin. Microbiol. 2000, 38, 393–394. [Google Scholar] [CrossRef]
- Garcia, L.S.; Shimizu, R.Y.; Novak, S.; Carroll, M.; Chan, F. Commercial assay for detection of Giardia lamblia and Cryptosporidium parvum antigens in human fecal specimens by rapid solid-phase qualitative immunochromatography. J. Clin. Microbiol. 2003, 41, 209–212. [Google Scholar] [CrossRef]
- Hooshyar, H.; Rostamkhani, P.; Arbabi, M.; Delavari, M. Giardia lamblia infection: Review of current diagnostic strategies. Gastroenterol. Hepatol. Bed Bench 2019, 12, 3–12. [Google Scholar]
- Khurana, S.; Chaudhary, P. Laboratory diagnosis of cryptosporidiosis. Trop. Parasitol. 2018, 8, 2–7. [Google Scholar] [CrossRef]
- Kabir, M.; Ahmed, E.; Hossain, B.; Alam, M.; Ahmed, S.; Taniuchi, M.; Gilchrist, C.A.; Houpt, E.R.; Faruque, A.; Petri, W.A., Jr. Giardia/Cryptosporidium QUIK CHEK assay is more specific than quantitative polymerase chain reaction for rapid point-of-care diagnosis of cryptosporidiosis in infants in Bangladesh. Clin. Infect. Dis. 2018, 67, 1897–1903. [Google Scholar] [CrossRef] [PubMed]
- Kabir, M.; Alam, M.; Nayak, U.; Arju, T.; Hossain, B.; Tarannum, R.; Khatun, A.; White, J.A.; Ma, J.N.Z.; Haque, R.; et al. Nonsterile immunity to cryptosporidiosis in infants is associated with mucosal IgA against the sporozoite and protection from malnutrition. PLoS Pathog. 2021, 17, e1009445. [Google Scholar] [CrossRef]
- Chang, L.J.; Hsiao, C.J.; Chen, B.; Liu, T.Y.; Ding, J.; Hsu, W.T.; Su-Ortiz, V.; Chen, S.T.; Su, K.Y.; Wu, H.P.; et al. Accuracy and comparison of two rapid multiplex PCR tests for gastroenteritis pathogens: A systematic review and meta-analysis. BMJ Open Gastroenterol. 2021, 8, e000553. [Google Scholar] [CrossRef]
- Farsi, T.A.; Weerakoon, S.; Mohsin, J.; Al Mashayakhi, H.; Ahmed, K.; Al Maani, A.; Aboqusida, K.; Al Sukaiti, N. Disseminated Cryptosporidiosis in an Infant with Non-HIV Pediatric Immunodeficiency: First Case Report from Oman. Oman Med. J. 2021, 36, e326. [Google Scholar] [CrossRef] [PubMed]
- Fox, L.M.; Saravolatz, L.D. Nitazoxanide: A new thiazolide antiparasitic agent. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2005, 40, 1173–1180. [Google Scholar] [CrossRef]
- Sparks, H.; Nair, G.; Castellanos-Gonzalez, A.; White, A.C., Jr. Treatment of Cryptosporidium: What We Know, Gaps, and the Way Forward. Curr. Trop. Med. Rep. 2015, 2, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Rossignol, J.F.; Ayoub, A.; Ayers, M.S. Treatment of diarrhea caused by Cryptosporidium parvum: A prospective randomized, double-blind, placebo-controlled study of Nitazoxanide. J. Infect. Dis. 2001, 184, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Amadi, B.; Mwiya, M.; Musuku, J.; Watuka, A.; Sianongo, S.; Ayoub, A.; Kelly, P. Effect of nitazoxanide on morbidity and mortality in Zambian children with cryptosporidiosis: A randomised controlled trial. Lancet 2002, 360, 1375–1380. [Google Scholar] [CrossRef]
- Amadi, B.; Mwiya, M.; Sianongo, S.; Payne, L.; Watuka, A.; Katubulushi, M.; Kelly, P. High dose prolonged treatment with nitazoxanide is not effective for cryptosporidiosis in HIV positive Zambian children: A randomised controlled trial. BMC Infect. Dis. 2009, 9, 195. [Google Scholar] [CrossRef]
- Di Santo, N.; Ehrisman, J. A functional perspective of nitazoxanide as a potential anticancer drug. Mutat. Res. Mol. Mech. Mutagen. 2014, 768, 16–21. [Google Scholar] [CrossRef]
- Jasenosky, L.D.; Cadena, C.; Mire, C.E.; Borisevich, V.; Haridas, V.; Ranjbar, S.; Nambu, A.; Bavari, S.; Soloveva, V.; Sadukhan, S.; et al. The FDA-Approved Oral Drug Nitazoxanide Amplifies Host Antiviral Responses and Inhibits Ebola Virus. iScience 2019, 19, 1279–1290. [Google Scholar] [CrossRef]
- Shane, A.L.; Mody, R.K.; Crump, J.A.; Tarr, P.I.; Steiner, T.S.; Kotloff, K.; Langley, J.M.; Wanke, C.; Warren, C.A.; Cheng, A.C.; et al. 2017 Infectious Diseases Society of America Clinical Practice Guidelines for the Diagnosis and Management of Infectious Diarrhea. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2017, 65, e45–e80. [Google Scholar] [CrossRef]
- Bajait, C.; Thawani, V. Role of zinc in pediatric diarrhea. Indian J. Pharmacol. 2011, 43, 232–235. [Google Scholar] [CrossRef]
- Li, S.T.; Grossman, D.C.; Cummings, P. Loperamide therapy for acute diarrhea in children: Systematic review and meta-analysis. PLoS Med. 2007, 4, e98. [Google Scholar] [CrossRef] [PubMed]
- Nabarro, L.E.; Lever, R.A.; Armstrong, M.; Chiodini, P.L. Increased incidence of nitroimidazole-refractory giardiasis at the Hospital for Tropical Diseases, London: 2008–2013. Clin. Microbiol. Infect. 2015, 21, 791–796. [Google Scholar] [CrossRef] [PubMed]
- Farthing, M.J. Giardiasis. Gastroenterol. Clin. N. Am. 1996, 25, 493–515. [Google Scholar] [CrossRef] [PubMed]
- Tejman-Yarden, N.; Eckmann, L. New approaches to the treatment of giardiasis. Curr. Opin. Infect. Dis. 2011, 24, 451–456. [Google Scholar] [CrossRef]
- Lalle, M.; Hanevik, K. Treatment-refractory giardiasis: Challenges and solutions. Infect. Drug Resist. 2018, 11, 1921–1933. [Google Scholar] [CrossRef]
- Leitsch, D. Drug Resistance in the Microaerophilic Parasite Giardia lamblia. Curr. Trop. Med. Rep. 2015, 2, 128–135. [Google Scholar] [CrossRef]
- Gardner, T.B.; Hill, D.R. Treatment of giardiasis. Clin. Microbiol. Rev. 2001, 14, 114–128. [Google Scholar] [CrossRef]
- Bawa, S.; Kumar, S.; Drabu, S.; Kumar, R. Structural modifications of quinoline-based antimalarial agents: Recent developments. J. Pharm. Bioallied Sci. 2010, 2, 64–71. [Google Scholar] [CrossRef]
- Lofmark, S.; Edlund, C.; Nord, C.E. Metronidazole is still the drug of choice for treatment of anaerobic infections. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2010, 50 (Suppl. 1), S16–S23. [Google Scholar] [CrossRef]
- Riches, A.; Hart, C.J.S.; Trenholme, K.R.; Skinner-Adams, T.S. Anti-Giardia Drug Discovery: Current Status and Gut Feelings. J. Med. Chem. 2020, 63, 13330–13354. [Google Scholar] [CrossRef]
- Ibrahim, A.A.; El-Housseiny, G.S.; Aboshanab, K.M.; Yassien, M.A.; Hassouna, N.A. Paromomycin production from Streptomyces rimosus NRRL 2455: Statistical optimization and new synergistic antibiotic combinations against multidrug resistant pathogens. BMC Microbiol. 2019, 19, 18. [Google Scholar] [CrossRef]
- Singh, N.; Narayan, S. Nitazoxanide A Broad Spectrum Antimicrobial. Med. J. Armed Forces India 2011, 67, 67–68. [Google Scholar] [CrossRef] [PubMed]
- Rossignol, J.F.; Lopez-Chegne, N.; Julcamoro, L.M.; Carrion, M.E.; Bardin, M.C. Nitazoxanide for the empiric treatment of pediatric infectious diarrhea. Trans. R. Soc. Trop. Med. Hyg. 2012, 106, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Standing, J.F.; Ongas, M.O.; Ogwang, C.; Kagwanja, N.; Murunga, S.; Mwaringa, S.; Ali, R.; Mturi, N.; Timbwa, M.; Manyasi, C.; et al. Dosing of Ceftriaxone and Metronidazole for Children With Severe Acute Malnutrition. Clin. Pharmacol. Ther. 2018, 104, 1165–1174. [Google Scholar] [CrossRef] [PubMed]
- Speelman, P. Single-Dose Tinidazole for the Treatment of Giardiasis. Antimicrob. Agents Chemother. 1985, 27, 227–229. [Google Scholar] [CrossRef] [PubMed]
- Escobedo, A.A.; Alvarez, G.; Gonzalez, M.E.; Almirall, P.; Canete, R.; Cimerman, S.; Ruiz, A.; Perez, R. The treatment of giardiasis in children: Single-dose tinidazole compared with 3 days of nitazoxanide. Ann. Trop. Med. Parasitol. 2008, 102, 199–207. [Google Scholar] [CrossRef]
- Gillis, J.C.; Wiseman, L.R. Secnidazole. A review of its antimicrobial activity, pharmacokinetic properties and therapeutic use in the management of protozoal infections and bacterial vaginosis. Drugs 1996, 51, 621–638. [Google Scholar] [CrossRef]
- Morch, K.; Hanevik, K. Giardiasis treatment: An update with a focus on refractory disease. Curr. Opin. Infect. Dis. 2020, 33, 355–364. [Google Scholar] [CrossRef]
- Perez, F.; Vallet, T.; Bravo, Z.; Callahan, K.; Ruiz, F. Acceptability of Mebendazole Chewable Tablet in Children Aged 2 to 4 Years in Peru. Pharmaceutics 2021, 14, 27. [Google Scholar] [CrossRef]


| Some Differences | Similarities | References | |
|---|---|---|---|
| Giardia | Cryptosporidium | ||
| Microbial biofilm disruption, altered species diversity, and abundance in intestinal microbiota. | No significant changes in the bacterial microbiota in diversity or structure. | Diarrhea is a common associated symptom. | [91,102] |
| Increased cases more often in children aged 3–6 months old. | More common in children aged 6–12 months old. | Infection lasts longer in children than adults with treatment. | [62,91,103] |
| Children are 17.9-fold more sensitive to infection than adults. | Children with Cryptosporidium are 10.6-fold more sensitive to the infection than adults. | Could cause malnutrition due to lack of nutrient absorption. | [102,104] |
| Giardia infection in children may cause inflammation of the joints. | Children that are HIV-positive are more susceptible to Cryptosporidium. | No significant difference in prevalence between males and females among children. | [91,104,105] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prabakaran, M.; Weible, L.J.; Champlain, J.D.; Jiang, R.Y.; Biondi, K.; Weil, A.A.; Van Voorhis, W.C.; Ojo, K.K. The Gut-Wrenching Effects of Cryptosporidiosis and Giardiasis in Children. Microorganisms 2023, 11, 2323. https://doi.org/10.3390/microorganisms11092323
Prabakaran M, Weible LJ, Champlain JD, Jiang RY, Biondi K, Weil AA, Van Voorhis WC, Ojo KK. The Gut-Wrenching Effects of Cryptosporidiosis and Giardiasis in Children. Microorganisms. 2023; 11(9):2323. https://doi.org/10.3390/microorganisms11092323
Chicago/Turabian StylePrabakaran, Mayuri, Lyssa J. Weible, Joshua D. Champlain, Ryan Ye Jiang, Katalina Biondi, Ana A. Weil, Wesley C. Van Voorhis, and Kayode K. Ojo. 2023. "The Gut-Wrenching Effects of Cryptosporidiosis and Giardiasis in Children" Microorganisms 11, no. 9: 2323. https://doi.org/10.3390/microorganisms11092323
APA StylePrabakaran, M., Weible, L. J., Champlain, J. D., Jiang, R. Y., Biondi, K., Weil, A. A., Van Voorhis, W. C., & Ojo, K. K. (2023). The Gut-Wrenching Effects of Cryptosporidiosis and Giardiasis in Children. Microorganisms, 11(9), 2323. https://doi.org/10.3390/microorganisms11092323