

Halanaerobium polyolivorans sp. nov.—A Novel Halophilic Alkalitolerant Bacterium Capable of Polyol Degradation: Physiological Properties and Genomic Insights
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Enrichment and Isolation
2.2. Morphological Characterization
2.3. Physiological Characterization
2.4. Analytical Assays
2.5. Genome Sequencing and Bioinformatic Analyses
3. Results
3.1. Isolation
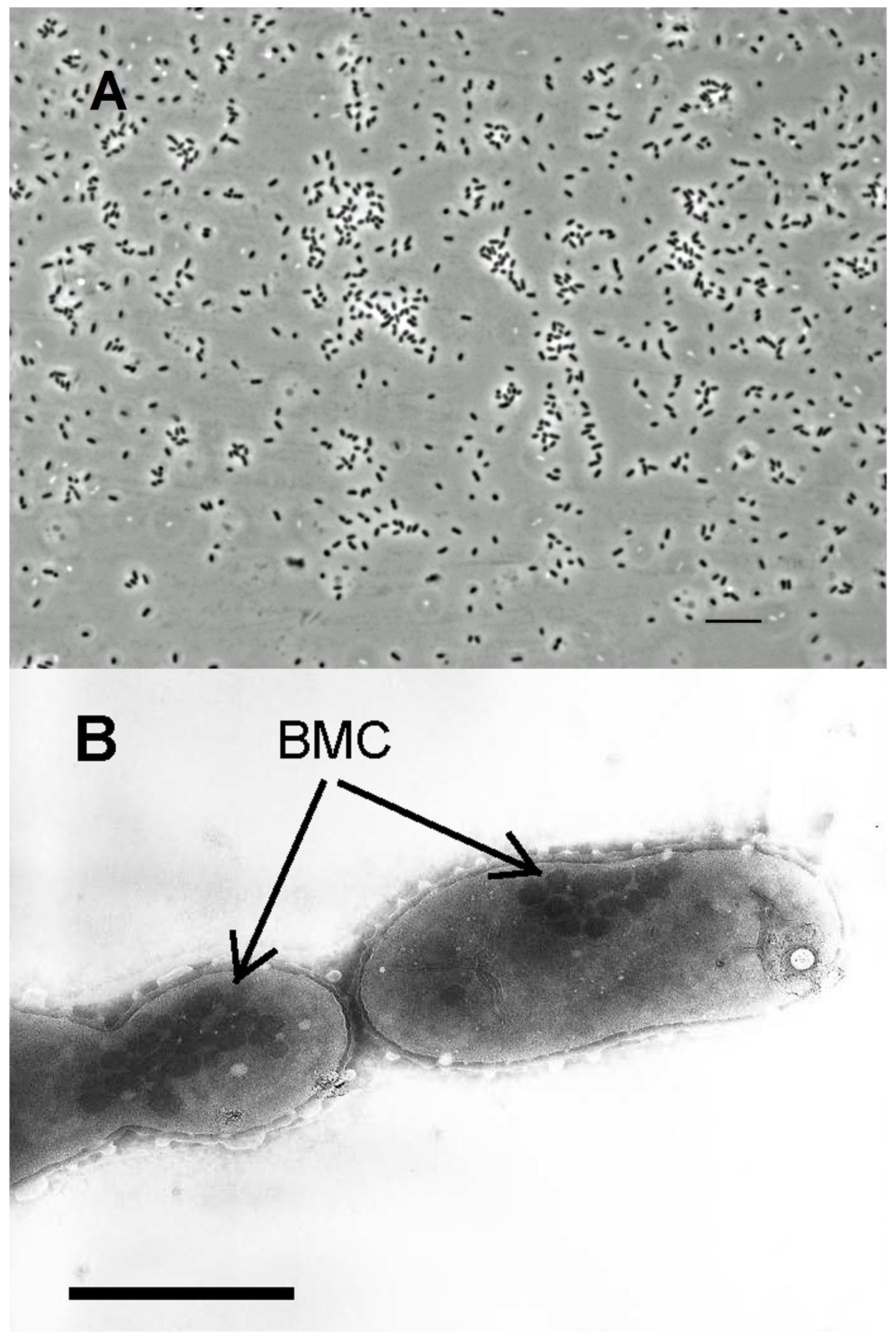
3.2. Colonies and Cell Morphology
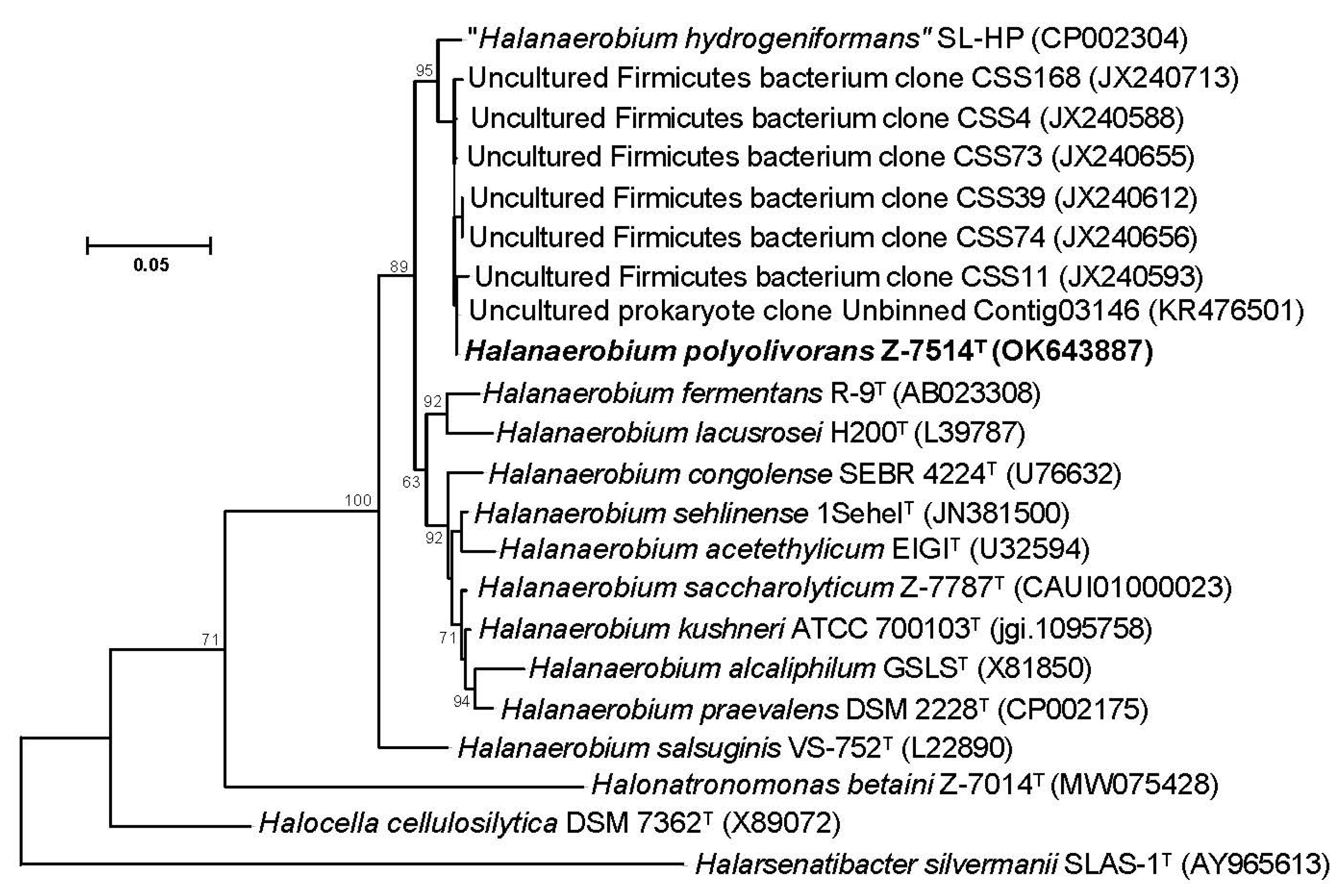
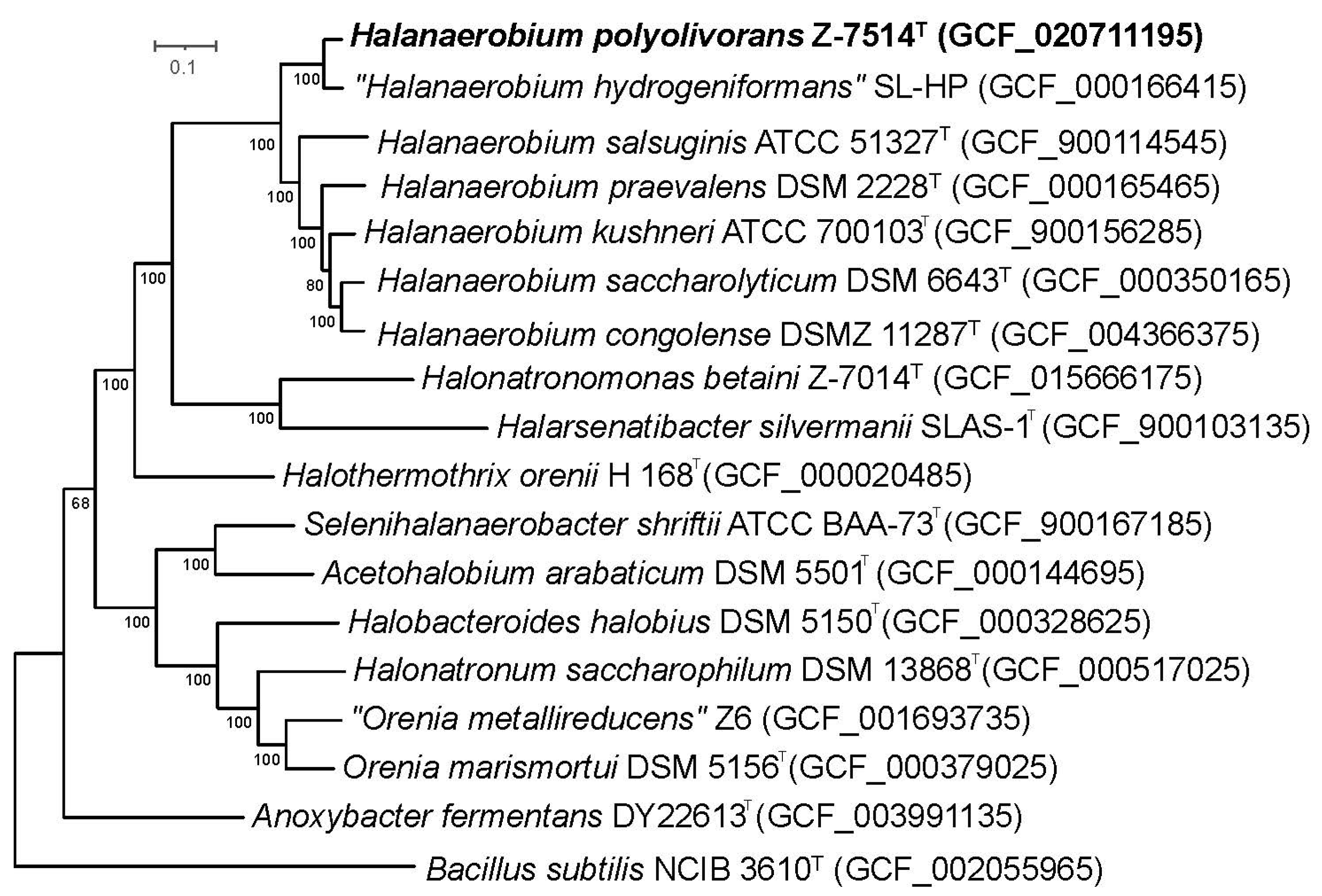
3.3. Phylogenetic and Genomic Characterization
3.4. Physiological Properties
3.4.1. Physicochemical Characteristics of Growth
3.4.2. Substrates, Electron Acceptors and Phenotypical Comparisons
3.4.3. Utilization of Polyols
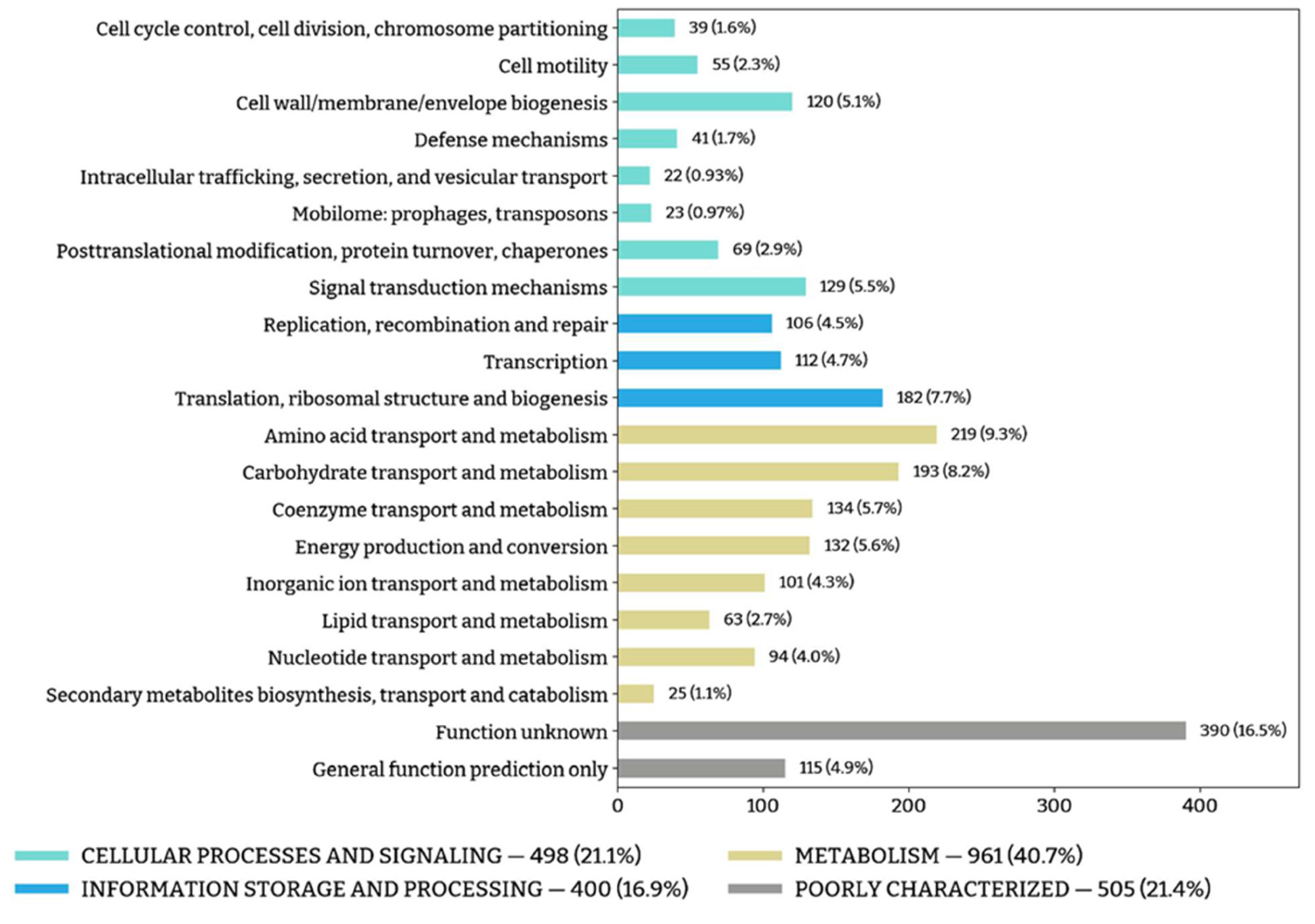
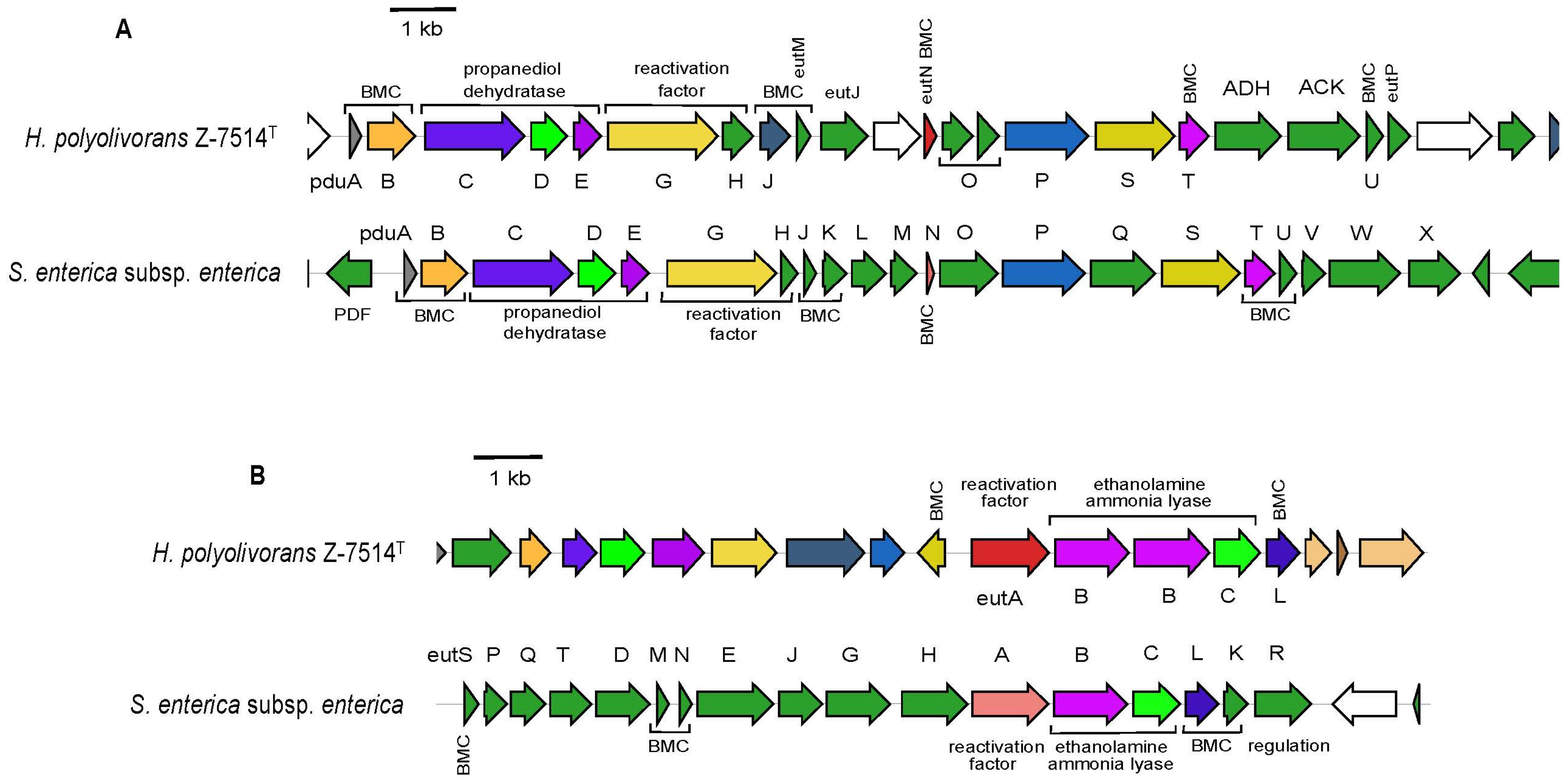
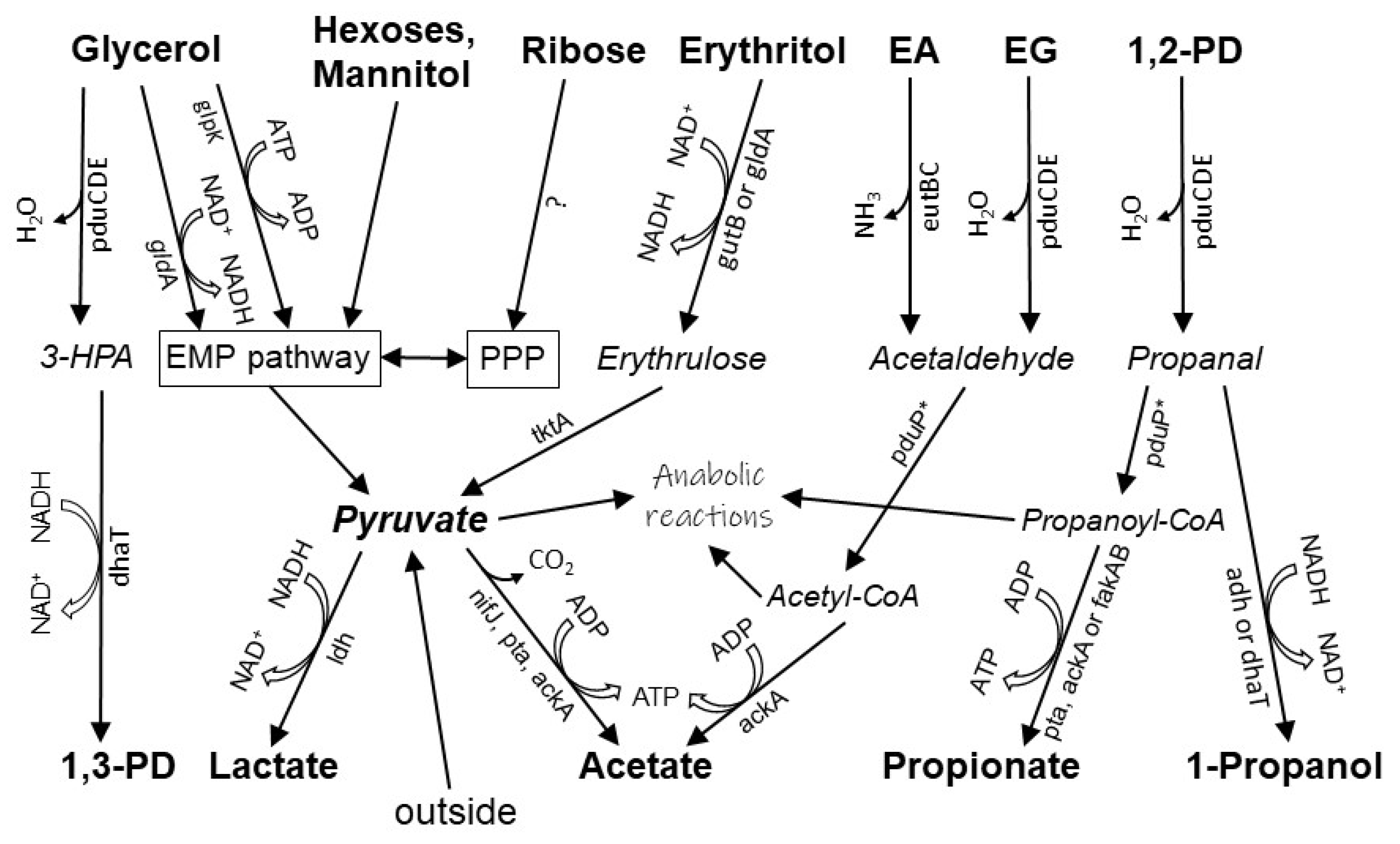
3.5. Physiology and Functional Genes
4. Discussion
5. Description of Halanaerobium polyolivorans sp. nov.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grant, W.D.; Jones, B.E. Bacteria, archaea and viruses of soda lakes. In Soda Lakes of East Africa; Schagerl, M., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 97–147. [Google Scholar] [CrossRef]
- Zhilina, T.N.; Zavarzin, G.A. Alkaliphilic anaerobic community at pH 10. Curr. Microbiol. 1994, 28, 109–112. [Google Scholar] [CrossRef]
- Sorokin, D.Y.; Banciu, H.L.; Muyzer, G. Functional microbiology of soda lakes. Curr. Opin. Microbiol. 2015, 25, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Boltyanskaya, Y.; Kevbrin, V.V. Laboratory simulation of “Proteolytic bacterium—Cyanobacterium” interaction in alkaliphilic microbial community. Paleontol. J. 2018, 52, 1179–1185. [Google Scholar] [CrossRef]
- Jennings, D.H. Polyol metabolism in fungi. In Advances in Microbial Physiology; Rose, A.H., Tempest, D.W., Eds.; Academic Press: London, UK, 1985; Volume 25, pp. 149–183. [Google Scholar] [CrossRef]
- Ônishi, H. Studies on osmophilic yeasts. Agric. Biol. Chem. 1963, 27, 543–547. [Google Scholar] [CrossRef]
- Oren, A. Glycerol metabolism in hypersaline environments. Environ. Microbiol. 2017, 19, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Ben-Amotz, A. Bioactive compounds: Glycerol production, carotenoid production, fatty acids production. In The Alga Dunaliella: Biodiversity, Physiology, Genomics and Biotechnology, 1st ed.; Ben-Amotz, A., Polle, J.E.W., Subba Rao, D.V., Eds.; Science Publishers: Enfield, NH, USA, 2009; pp. 189–208. [Google Scholar]
- Samylina, O.S.; Sapozhnikov, F.V.; Gainanova, O.Y.; Ryabova, A.V.; Nikitin, M.A.; Sorokin, D.Y. Algo-bacterial communities of the Kulunda Steppe (Altai Region, Russia) soda lakes. Microbiology 2014, 83, 849–860. [Google Scholar] [CrossRef]
- Kevbrin, V.; Boltyanskaya, Y.; Zhilina, T.; Lavrentjeva, E.; Kuznetsov, B. Proteinivorax tanatarense gen. nov., sp. nov., an anaerobic, haloalkaliphilic, proteolytic bacterium isolated from decaying algal bloom, and proposal of Proteinivoraceae fam. nov. Extremophiles 2013, 17, 747–756. [Google Scholar] [CrossRef]
- Boltyanskaya, Y.V.; Kevbrin, V.V.; Grouzdev, D.S.; Detkova, E.N.; Koziaeva, V.V.; Novikov, A.A.; Zhilina, T.N. Halonatronomonas betaini gen. nov., sp. nov., a haloalkaliphilic isolate from soda lake capable of betaine degradation and proposal of Halarsenatibacteraceae fam. nov. and Halothermotrichaceae fam. nov. within the order Halanaerobiales. Syst. Appl. Microbiol. 2023, 46, 126407. [Google Scholar] [CrossRef]
- Boronat, A.; Aguilar, J. Metabolism of L-fucose and L-rhamnose in Escherichia coli: Differences in induction of propanediol oxidoreductase. J. Bacteriol. 1981, 147, 181–185. [Google Scholar] [CrossRef]
- Tao, Y.-m.; Bu, C.-y.; Zou, L.-h.; Hu, Y.-l.; Zheng, Z.-J.; Ouyang, J. A comprehensive review on microbial production of 1,2-propanediol: Microorganisms, metabolic pathways, and metabolic engineering. Biotechnol. Biofuels 2021, 14, 216. [Google Scholar] [CrossRef]
- Blomberg, A.; Adler, L. Physiology of osmotolerance in fungi. In Advances in Microbial Physiology; Rose, A.H., Ed.; Academic Press: London, UK, 1992; Volume 33, pp. 145–212. [Google Scholar] [CrossRef]
- Gunde-Cimerman, N.; Ramos, J.; Plemenitaš, A. Halotolerant and halophilic fungi. Mycol. Res. 2009, 113, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Zeikus, J.G.; Hegge, P.W.; Thompson, T.E.; Phelps, T.J.; Langworthy, T.A. Isolation and description of Haloanaerobium praevalens gen. nov. and sp. nov., an obligately anaerobic halophile common to Great Salt Lake sediments. Curr. Microbiol. 1983, 9, 225–234. [Google Scholar] [CrossRef]
- Brown, S.D.; Begemann, M.B.; Mormile, M.R.; Wall, J.D.; Han, C.S.; Goodwin, L.A.; Pitluck, S.; Land, M.L.; Hauser, L.J.; Elias, D.A. Complete genome sequence of the haloalkaliphilic, hydrogen-producing bacterium Halanaerobium hydrogeniformans. J. Bacteriol. 2011, 193, 3682–3683. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. Halanaerobium corrig. Zeikus, Hegge, Thompson, Phelps and Langworthy 1984, 503VP. In Bergey’s Manual of Systematics of Archaea and Bacteria; In Association with Bergey’s Manual Trust; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015. [Google Scholar] [CrossRef]
- Alvarez-Coque, M.C.; Hernandez, M.J.; Camanas, R.M.; Fernandez, C. Studies on the formation and stability of isoindoles derived from amino acids, o-phthalaldehyde and N-acetyl-L-cysteine. Anal. Biochem. 1989, 180, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Trüper, H.G.; Schlegel, H.G. Sulphur metabolism in Thiorhodaceae. I. Quantitative measurements on growing cells of Chromatium okenii. Antonie van Leeuwenhoek 1964, 30, 225–238. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley & Sons, Inc.: Chichester, UK, 1991; pp. 115–175. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Chaumeil, P.-A.; Mussig, A.J.; Hugenholtz, P.; Parks, D.H. GTDB-Tk: A toolkit to classify genomes with the Genome Taxonomy Database. Bioinformatics 2020, 36, 1925–1927. [Google Scholar] [CrossRef]
- Grouzdev, D.S.; Rysina, M.S.; Bryantseva, I.A.; Gorlenko, V.M.; Gaisin, V.A. Draft genome sequences of “Candidatus Chloroploca asiatica” and “Candidatus Viridilinea mediisalina”, candidate representatives of the Chloroflexales order: Phylogenetic and taxonomic implications. Stand. Genomic Sci. 2018, 13, 24. [Google Scholar] [CrossRef] [PubMed]
- Qin, Q.-L.; Xie, B.-B.; Zhang, X.-Y.; Chen, X.-L.; Zhou, B.-C.; Zhou, J.; Oren, A.; Zhang, Y.-Z. A proposed genus boundary for the prokaryotes based on genomic insights. J. Bacteriol. 2014, 196, 2210–2215. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Sardà Carbasse, J.; Peinado-Olarte, R.L.; Göker, M. TYGS and LPSN: A database tandem for fast and reliable genome-based classification and nomenclature of prokaryotes. Nucleic Acid. Res. 2022, 50, D801–D807. [Google Scholar] [CrossRef] [PubMed]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Galperin, M.Y.; Makarova, K.S.; Wolf, Y.I.; Koonin, E.V. Expanded microbial genome coverage and improved protein family annotation in the COG database. Nucleic Acids Res. 2014, 43, D261–D269. [Google Scholar] [CrossRef]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef]
- Sequeira, J.C.; Rocha, M.; Alves, M.M.; Salvador, A.F. UPIMAPI, reCOGnizer and KEGGCharter: Bioinformatics tools for functional annotation and visualization of (meta)-omics datasets. Comput. Struct. Biotechnol. J. 2022, 20, 1798–1810. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG tools for functional characterization of genome and metagenome sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef]
- Chang, A.; Jeske, L.; Ulbrich, S.; Hofmann, J.; Koblitz, J.; Schomburg, I.; Neumann-Schaal, M.; Jahn, D.; Schomburg, D. BRENDA, the ELIXIR core data resource in 2021: New developments and updates. Nucleic Acids Res. 2021, 49, D498–D508. [Google Scholar] [CrossRef] [PubMed]
- Keshri, J.; Yousuf, B.; Mishra, A.; Jha, B. The abundance of functional genes, cbbL, nifH, amoA and apsA, and bacterial community structure of intertidal soil from Arabian Sea. Microbiol. Res. 2015, 175, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Nolla-Ardèvol, V.; Strous, M.; Tegetmeyer, H.E. Anaerobic digestion of the microalga Spirulina at extreme alkaline conditions: Biogas production, metagenome, and metatranscriptome. Front. Microbiol. 2015, 6, 597. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Klenk, H.; Göker, M. Taxonomic use of DNA G+C content and DNA–DNA hybridization in the genomic age. Int. J. Syst. Evol. Microbiol. 2014, 64, 352–356. [Google Scholar] [CrossRef]
- Chun, J.; Oren, A.; Ventosa, A.; Christensen, H.; Arahal, D.R.; da Costa, M.S.; Rooney, A.P.; Yi, H.; Xu, X.W.; De Meyer, S.; et al. Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Microbiol. 2018, 68, 461–466. [Google Scholar] [CrossRef]
- Konstantinidis, K.T.; Tiedje, J.M. Towards a genome-based taxonomy for prokaryotes. J. Bacteriol. 2005, 187, 6258–6264. [Google Scholar] [CrossRef]
- Blackwell, C.M.; Turner, J.M. Microbial metabolism of amino alcohols: Purification and properties of coenzyme B12-dependent ethanolamine ammonia-lyase of Escherichia coli. Biochem. J. 1978, 175, 555–563. [Google Scholar] [CrossRef]
- Sheppard, D.E.; Penrod, J.T.; Bobik, T.; Kofoid, E.; Roth, J.R. Evidence that a B12-adenosyl transferase is encoded within the ethanolamine operon of Salmonella enterica. J. Bacteriol. 2004, 186, 7635–7644. [Google Scholar] [CrossRef]
- Zhilina, T.N.; Zavarzin, G.A.; Bulygina, E.S.; Kevbrin, V.V.; Osipov, G.A.; Chumakov, K.M. Ecology, physiology and taxonomy studies on a new taxon of Haloanaerobiaceae, Haloincola saccharolytica gen. nov., sp. nov. Syst. Appl. Microbiol. 1992, 15, 275–284. [Google Scholar] [CrossRef]
- Ravot, G.; Magot, M.; Ollivier, B.; Patel, B.K.C.; Ageron, E.; Grimont, P.A.D.; Thomas, P.; Garcia, J.-L. Halanaerobium congolense sp. nov., an anaerobic, moderately halophilic thiosulfate- and sulfur-reducing bacterium from an African oil field. FEMS Microbiol. Lett. 1997, 147, 81–88. [Google Scholar] [CrossRef]
- Bhupathiraju, V.K.; Mclnerney, M.J.; Woese, C.R.; Tanner, R.S. Haloanaerobiurn kushneri sp. nov., an obligately halophilic, anaerobic bacterium from an oil brine. Int. J. Syst. Bacteriol. 1999, 49, 953–960. [Google Scholar] [CrossRef] [PubMed]
- Barbier, T.; Collard, F.; Zuniga-Ripa, A.; Moriyon, I.; Godard, T.; Becker, J.; Wittmann, C.; Van Schaftingen, E.; Letesson, J.J. Erythritol feeds the pentose phosphate pathway via three new isomerases leading to D-erythrose-4-phosphate in Brucella. Proc. Natl. Acad. Sci. USA 2014, 111, 17815–17820. [Google Scholar] [CrossRef] [PubMed]
- Scharschmidt, M.; Pfleiderer, G.; Metz, H.; Brummer, W. Isolierung und charakterisierung von glycerol-dehydrogenase aus Bacillus megaterium, Hoppe-Seyler’s Z. Physiol. Chem. 1983, 364, 911–921. [Google Scholar] [CrossRef]
- Sugisawa, T.; Hoshino, T. Purification and properties of membrane-bound D-sorbitol dehydrogenase from Gluconobacter suboxydans IFO 3255. Biosci. Biotechnol. Biochem. 2002, 66, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.A.; Abeles, R.H. Purification and properties of dioldehydrase, an enzyme requiring a cobamide coenzyme. J. Biol. Chem. 1963, 238, 2367–2373. [Google Scholar] [CrossRef] [PubMed]
- Toraya, T.; Honda, S.; Fukui, S. Fermentation of 1,2-propanediol and 1,2-ethanediol by some genera of Enterobacteriaceae, involving coenzyme B12-dependent diol dehydratase. J. Bacteriol. 1979, 139, 39–47. [Google Scholar] [CrossRef]
- Bobik, T.A.; Xu, Y.; Jeter, R.M.; Otto, K.E.; Roth, J.R. Propanediol utilization genes (pdu) of Salmonella typhimurium: Three genes for the propanediol dehydratase. J. Bacteriol. 1997, 179, 6633–6639. [Google Scholar] [CrossRef]
- Sampson, E.M.; Bobik, T.A. Microcompartments for B12-dependent 1,2-propanediol degradation provide protection from DNA and cellular damage by a reactive metabolic intermediate. J. Bacteriol. 2008, 190, 2966–2971. [Google Scholar] [CrossRef]
- Moore, T.C.; Escalante-Semerena, J.C. The EutQ and EutP proteins are novel acetate kinases involved in ethanolamine catabolism: Physiological implications for the function of the ethanolamine metabolosome in Salmonella enterica. Mol. Microbiol. 2016, 99, 497–511. [Google Scholar] [CrossRef]
- Vavourakis, C.D.; Andrei, A.-S.; Mehrshad, M.; Ghai, R.; Sorokin, D.Y.; Muyzer, G. A metagenomics roadmap to the uncultured genome diversity in hypersaline soda lake sediments. Microbiome 2018, 6, 168. [Google Scholar] [CrossRef]
- Leichus, B.N.; Blanchard, J.S. Isotopic analysis of the reaction catalyzed by glycerol dehydrogenase. Biochemistry 1994, 33, 14642–14649. [Google Scholar] [CrossRef] [PubMed]
- Bachochin, M.J.; Van Allen, M.; Barber, R.D. Characterization of a Rhodobacter sphaeroides primary fatty acid kinase. Arch. Microbiol. 2021, 203, 861–864. [Google Scholar] [CrossRef] [PubMed]
- Bock, A.-K.; Glasemacher, J.; Schmidt, R.; Schönheit, P. Purification and characterization of two extremely thermostable enzymes, phosphate acetyltransferase and acetate kinase, from the hyperthermophilic eubacterium Thermotoga maritima. J. Bacteriol. 1999, 181, 1861–1867. [Google Scholar] [CrossRef]
- Daniel, R.; Boenigk, R.; Gottschalk, G. Purification of 1,3-propanediol dehydrogenase from Citrobacter freundii and cloning, sequencing, and overexpression of the corresponding gene in Escherichia coli. J. Bacteriol. 1995, 177, 2151–2156. [Google Scholar] [CrossRef]
- Stewart, A.M.; Stewart, K.L.; Yeates, T.O.; Bobik, T.A. Advances in the world of bacterial microcompartments. Trends Biochem. Sci. 2021, 46, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Kaval, K.G.; Garsin, D.A. Ethanolamine utilization in bacteria. mBio 2018, 9, e00066-18. [Google Scholar] [CrossRef]
- Tanaka, S.; Sawaya, M.R.; Yeates, T.O. Structure and mechanisms of a protein-based organelle in Escherichia coli. Science 2010, 327, 81–84. [Google Scholar] [CrossRef]
- Larson, T.J.; Ehrmann, M.; Boos, W. Periplasmic glycerophosphodiester phosphodiesterase of Escherichia coli, a new enzyme of the glp regulon. J. Biol. Chem. 1983, 258, 5428–5432. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Attribute | Value | % of Total |
|---|---|---|
| Genome size (bp) | 2,522,622 | 100.0 |
| DNA coding (bp) | 2,266,477 | 89.9 |
| DNA G+C (bp) | 839,862 | 33.3 |
| DNA scaffolds | 53 | 100.0 |
| Total genes | 2457 | 100.0 |
| Protein coding genes | 2364 | 96.2 |
| RNA genes | 63 | 2.6 |
| Pseudo genes | 30 | 1.2 |
| Genes with function prediction | 2215 | 90.2 |
| Genes assigned to COGs | 1974 | 80.3 |
| Genes with Pfam domains | 2116 | 86.1 |
| Genes with signal peptides | 185 | 7.5 |
| Genes with transmembrane helices | 615 | 25.0 |
| CRISPR repeats | 2 | - |
| Species of Halanaerobium | dDDH | ANI | AAI | POCP |
|---|---|---|---|---|
| ‘H. hydrogeniformans’ SL-HP GCF_000166415.1 | 29.9 | 84.8 | 88.4 | 86.8 |
| H. congolense DSM 11287T GCF_004366375.1 | 19.2 | 77.0 | 71.1 | 69.0 |
| H. saccharolyticum DSM 6643T GCF_000350165.1 | 18.7 | 77.2 | 70.9 | 72.2 |
| H. kushneri ATCC 700103T GCF_900156285.1 | 18.5 | 77.0 | 70.9 | 64.9 |
| H. praevalens_DSM 2228T GCF_000165465.1 | 18.8 | 77.0 | 69.3 | 63.7 |
| H. salsuginis ATCC 51327T GCF_900114545.1 | 18.8 | 77.0 | 68.2 | 61.9 |
| Substrate | End Products, mol/mol Substrate | Carbon Recovery, % | ||
|---|---|---|---|---|
| Acetate | Lactate | H2 | ||
| Pyruvate | 0.96 | 0 | 0.7 | 96 |
| D-ribose | 0.46 | 0.05 | 0.6 | 51 |
| D-glucose | 0.53 | 0.56 | 0.8 | 109 |
| D-fructose | 0.22 | 0.93 | 0.4 | 115 |
| Erythritol | 0.77 | 0.05 | 0.9 | 82 |
| Mannitol | 0.38 | 0.74 | 0.9 | 111 |
| Glycerol 1 | 0.46 | 0.11 | 7.0 | 81 |
| Ethanolamine 2 | 1.12 | 0 | 3.3 | 112 |
| Z-7514T | 1 H. kushneri VS-751T | 2 H. saccharolyticum Z-7787T | 3 H. praevalens GSLT | |
|---|---|---|---|---|
| Cell size, µm | 0.4–0.8 × 1.0–2.0 | 0.7 × 2.0–3.3 | 0.5–0.7 × 1.0–1.5 | 0.9–11.0 × 2.0–2.6 |
| Na+, M, range/opt | 0.6–3.9/1.6–2.1 | 1.5–3.1/2.1 | 0.5–5.1/1.7 | 0.3–5.1/2.1 |
| pH, range/opt | 6.7–10.1/8.0–8.5 | 6.0–8.0/6.5–7.5 | 6.0–8.0/7.5 | 6.0–9.0/7.0–7.4 |
| T, °C, range/opt | 14–51/31–35 | 20–45/40 | 15–47/37–40 | 5–50/37 |
| Utilization of | ||||
| L-arabinose | – | + | + | ND |
| Cellobiose | + | + | + | − |
| D-galactose | +/− | + | + | − |
| Glycerol | + | 4 var | + | − |
| Lactose | − | + | + | − |
| Pyruvate | + | + | + | − |
| Starch | − | − | ND | − |
| Sucrose | + | + | + | − |
| D-xylose | +/− | − | + | − |
| Trypticase | − | + | ND | + |
| Amino acids | − | − | − | + |
| N-acetylglucosamine | + | ND | − | + |
| Fermentation products from sugars | Acetate, lactate, H2, CO2 | Acetate, formate, ethanol, H2, CO2 | Acetate, H2, CO2 | Acetate, formate, lactate, ethanol, H2, CO2 |
| G+C, mol % from genome | 33.3 | 34.2 | 34.8 | 30.3 |
| Habitat | Alkaline sediments of a collector at Tanatar III soda lake, Russia | Hypersaline petroleum reservoir fluid, OK, USA | Hypersaline lagoons of Sivash Lake, Russia | Deep bottom sediment of Great Salt Lake, Utah, USA. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boltyanskaya, Y.; Zhilina, T.; Grouzdev, D.; Detkova, E.; Pimenov, N.; Kevbrin, V. Halanaerobium polyolivorans sp. nov.—A Novel Halophilic Alkalitolerant Bacterium Capable of Polyol Degradation: Physiological Properties and Genomic Insights. Microorganisms 2023, 11, 2325. https://doi.org/10.3390/microorganisms11092325
Boltyanskaya Y, Zhilina T, Grouzdev D, Detkova E, Pimenov N, Kevbrin V. Halanaerobium polyolivorans sp. nov.—A Novel Halophilic Alkalitolerant Bacterium Capable of Polyol Degradation: Physiological Properties and Genomic Insights. Microorganisms. 2023; 11(9):2325. https://doi.org/10.3390/microorganisms11092325
Chicago/Turabian StyleBoltyanskaya, Yulia, Tatjana Zhilina, Denis Grouzdev, Ekaterina Detkova, Nikolay Pimenov, and Vadim Kevbrin. 2023. "Halanaerobium polyolivorans sp. nov.—A Novel Halophilic Alkalitolerant Bacterium Capable of Polyol Degradation: Physiological Properties and Genomic Insights" Microorganisms 11, no. 9: 2325. https://doi.org/10.3390/microorganisms11092325
APA StyleBoltyanskaya, Y., Zhilina, T., Grouzdev, D., Detkova, E., Pimenov, N., & Kevbrin, V. (2023). Halanaerobium polyolivorans sp. nov.—A Novel Halophilic Alkalitolerant Bacterium Capable of Polyol Degradation: Physiological Properties and Genomic Insights. Microorganisms, 11(9), 2325. https://doi.org/10.3390/microorganisms11092325