


The Staphylococcus aureus ArlS Kinase Inhibitor Tilmicosin Has Potent Anti-Biofilm Activity in Both Static and Flow Conditions


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Plasmids
2.2. Growth Media and Chemical Agents
2.3. Construction of the arlRS Deletion Mutants
2.4. Semi-Quantitative Detection of Static Biofilms Formation
2.5. Purification of the Catalytic Domain of ArlS (ArlSHK)
2.6. ATPase Assay for Screening ArlSHK Kinase Activity Inhibitors
2.7. Promoter-Florescence Reporter Assay
2.8. Visualization of Three-Dimensional Structure of Biofilms by Confocal Laser Scanning Microscopy (CLSM)
2.9. Antimicrobial Susceptibility Test
2.10. Detection of Biofilm Formation in Flow Conditions by a BioFlux System
2.11. Statistical Analysis
3. Results
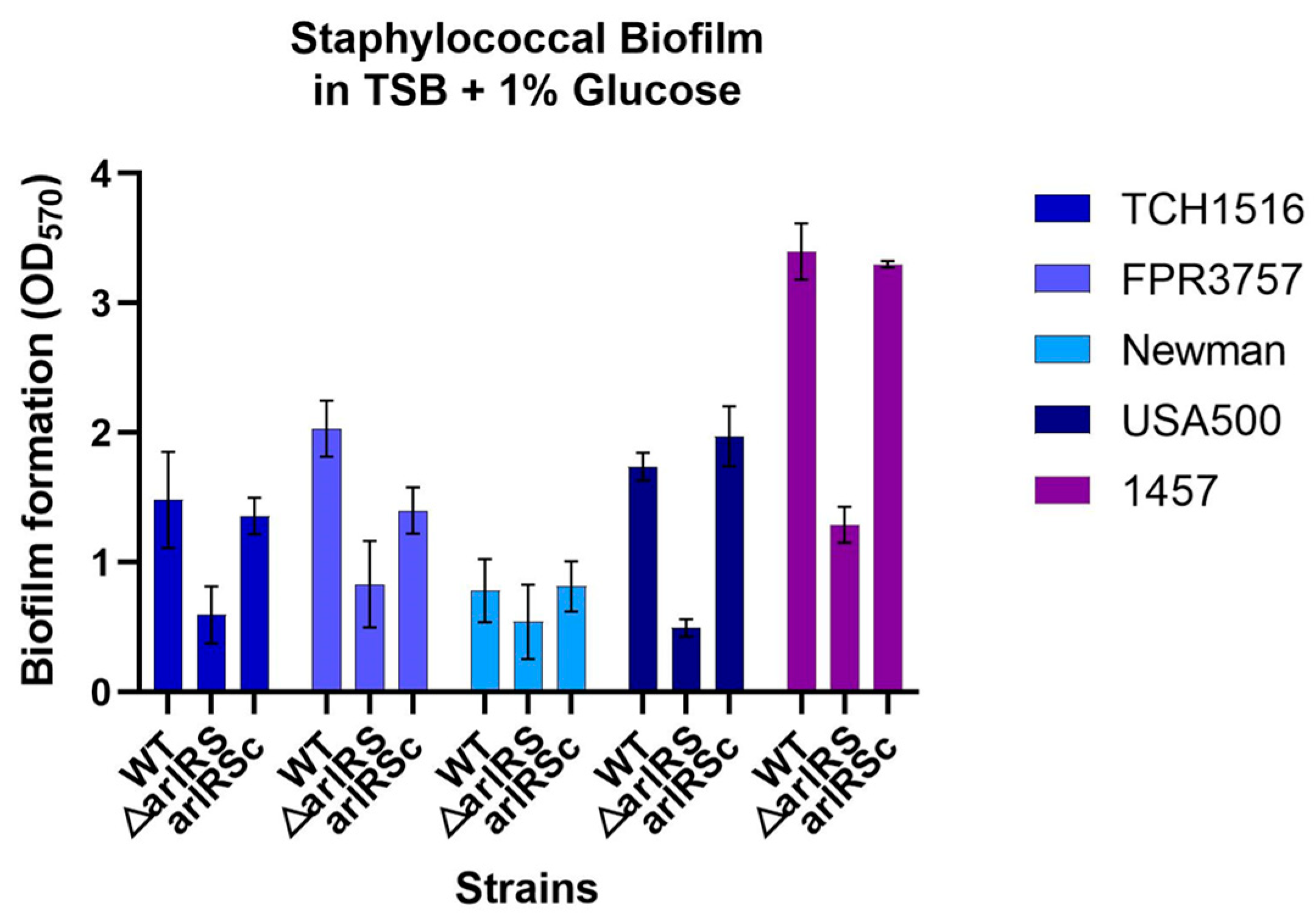
3.1. Effect of arlRS Knockout in Biofilm Formation in Multiple Staphylococcal Strains
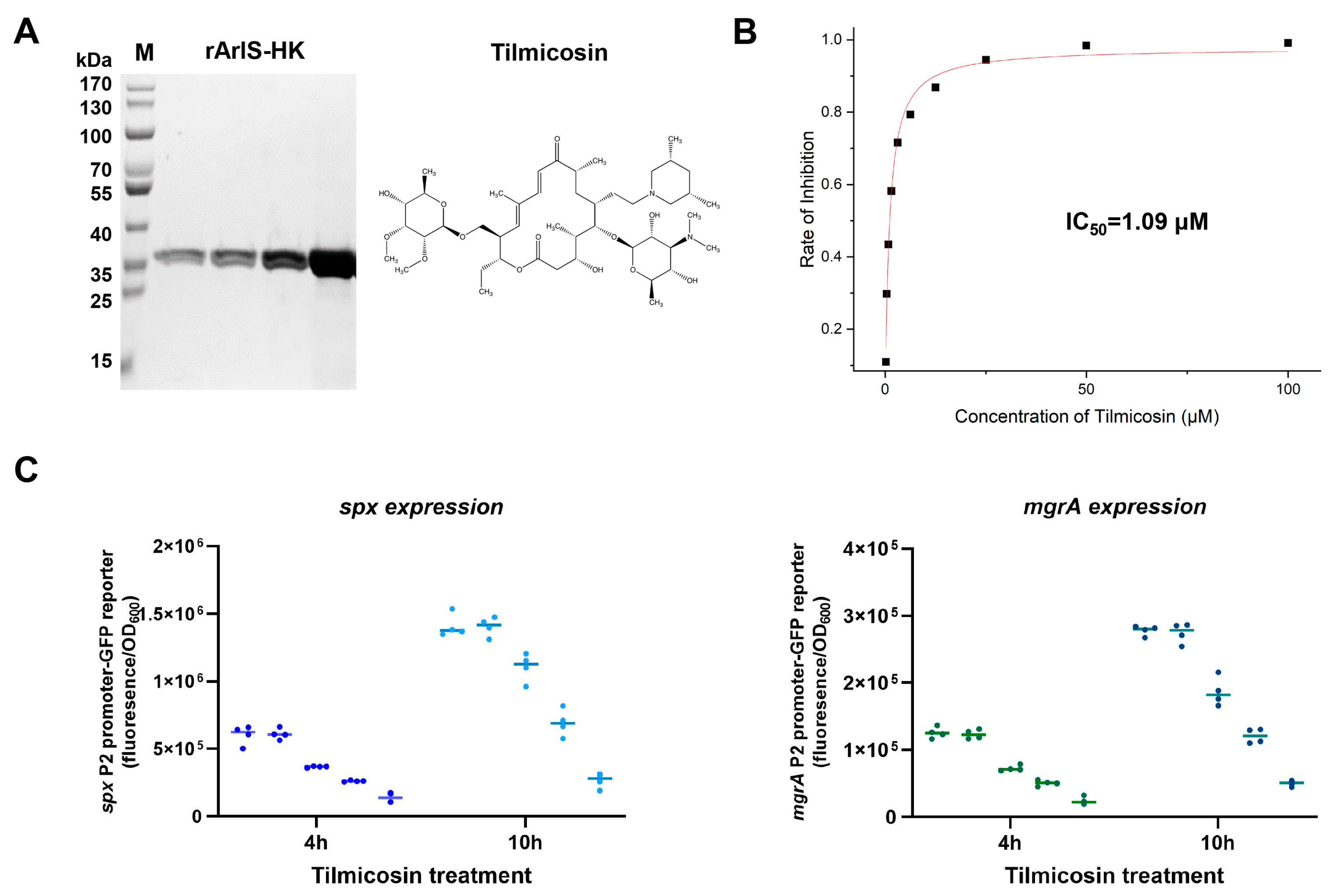
3.2. Inhibitory Effect of Tilmicosin on ArlS Activity
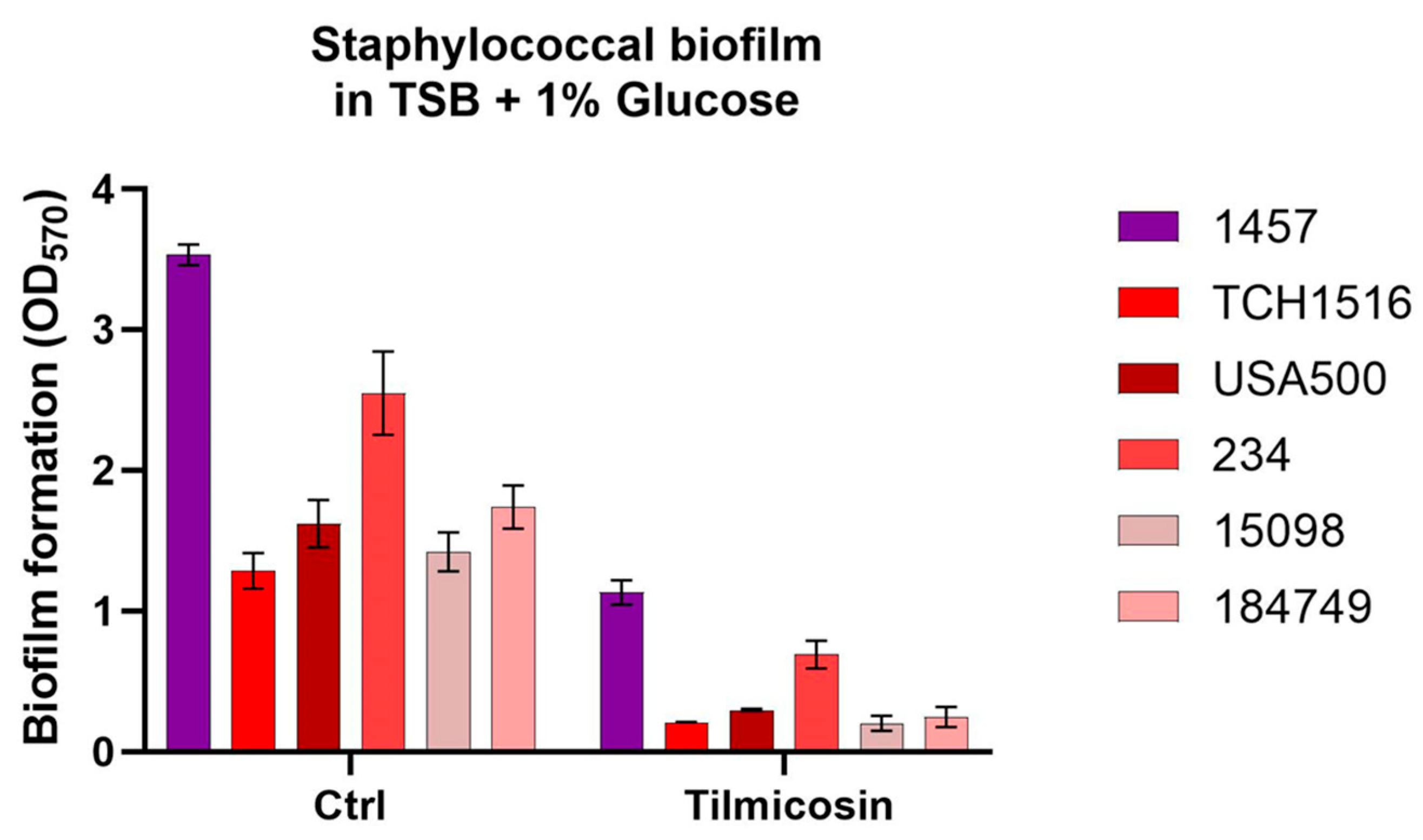
3.3. Effect of Tilmicosin on Static Biofilms Formation of the Staphylococcal Strains
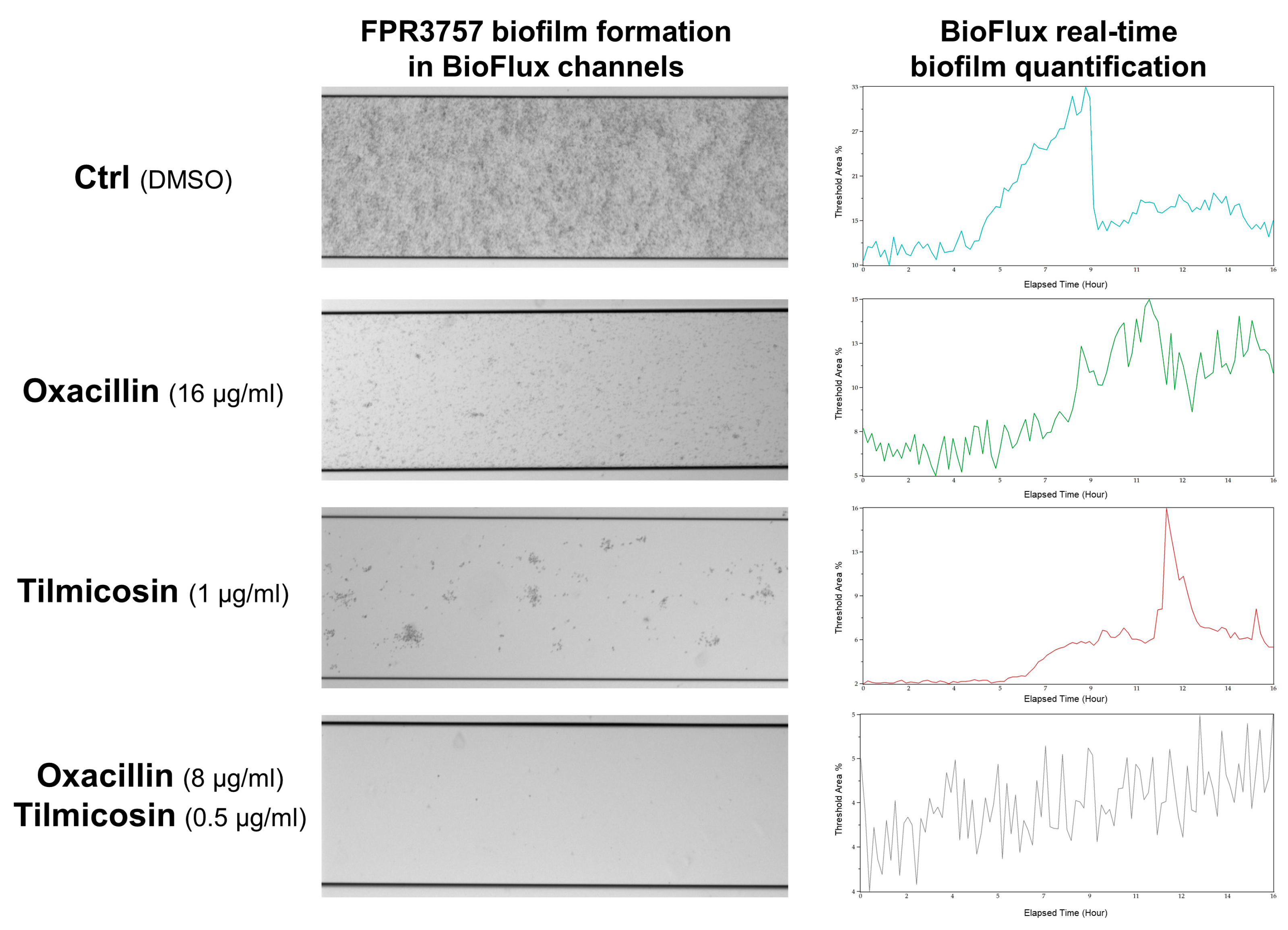
3.4. Effect of Tilmicosin on MRSA Biofilm Formation in a Flow Condition
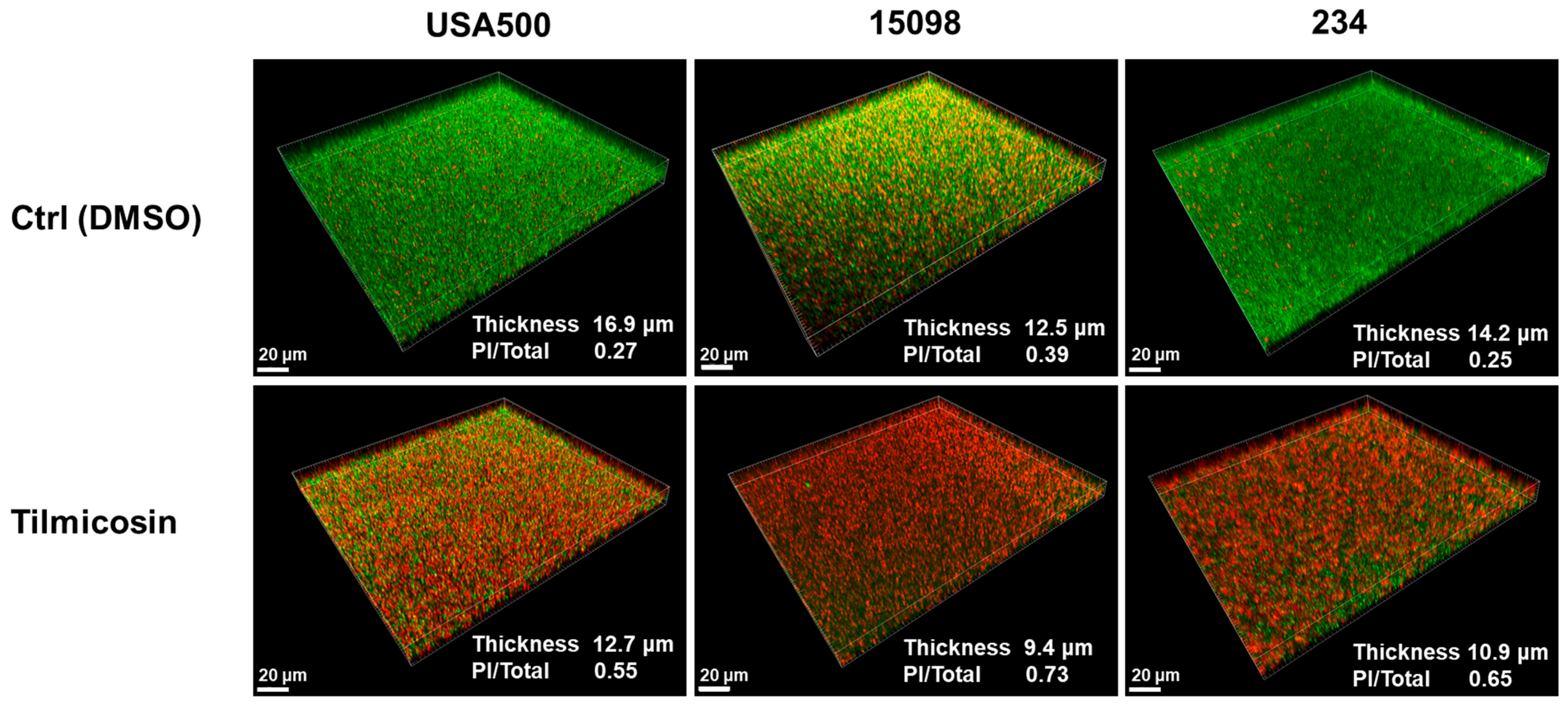
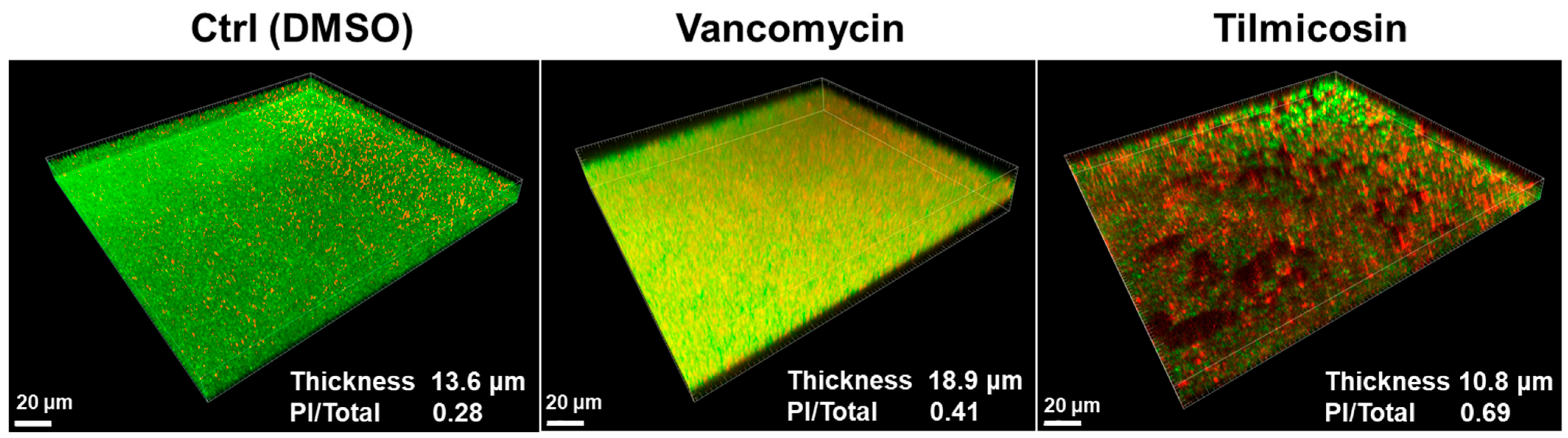
3.5. Effect of Tilmicosin on MRSA Mature Biofilms and Embedded Cells Viability
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tong, S.Y.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Community-associated MRSA: What makes them special? Int. J. Med. Microbiol. 2013, 303, 324–330. [Google Scholar] [CrossRef]
- Rybak, M.J.; Akins, R.L. Emergence of methicillin-resistant Staphylococcus aureus with intermediate glycopeptide resistance: Clinical significance and treatment options. Drugs 2001, 61, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Aires-de-Sousa, M. Methicillin-resistant Staphylococcus aureus among animals: Current overview. Clin. Microbiol. Infect. 2017, 23, 373–380. [Google Scholar] [CrossRef]
- Otto, M. Staphylococcal infections: Mechanisms of biofilm maturation and detachment as critical determinants of pathogenicity. Annu. Rev. Med. 2013, 64, 175–188. [Google Scholar] [CrossRef]
- Seethalakshmi, P.S.; Rajeev, R.; Kiran, G.S.; Selvin, J. Promising treatment strategies to combat Staphylococcus aureus biofilm infections: An updated review. Biofouling 2020, 36, 1159–1181. [Google Scholar]
- Liu, Y.; Zhang, J.; Ji, Y. Environmental factors modulate biofilm formation by Staphylococcus aureus. Sci. Prog. 2020, 103, 36850419898659. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Sun, X.; Wang, Z.; Zhang, Y. Biofilm-forming capacity of Staphylococcus epidermidis, Staphylococcus aureus, and Pseudomonas aeruginosa from ocular infections. Invest. Ophthalmol. Vis. Sci. 2012, 53, 5624–5631. [Google Scholar] [CrossRef]
- Darouiche, R.O. Treatment of infections associated with surgical implants. N. Engl. J. Med. 2004, 350, 1422–1429. [Google Scholar] [CrossRef]
- Qi, M.; Liu, Q.; Liu, Y.; Yan, H.; Zhang, Y.; Yuan, Y. Staphylococcus aureus biofilm inhibition by high voltage prick electrostatic field (HVPEF) and the mechanism investigation. Int. J. Food Microbiol. 2022, 362, 109499. [Google Scholar] [CrossRef]
- Pouget, C.; Dunyach-Remy, C.; Magnan, C.; Pantel, A.; Sotto, A.; Lavigne, J.P. Polymicrobial Biofilm Organization of Staphylococcus aureus and Pseudomonas aeruginosa in a Chronic Wound Environment. Int. J. Mol. Sci. 2022, 23, 10761. [Google Scholar] [CrossRef]
- Nasser, A.; Dallal, M.M.S.; Jahanbakhshi, S.; Azimi, T.; Nikouei, L. Staphylococcus aureus: Biofilm Formation and Strategies Against it. Curr. Pharm. Biotechnol. 2022, 23, 664–678. [Google Scholar] [CrossRef]
- Latorre, M.C.; Alonso, B.; Cruces, R.; Sanz, A.; Muñoz, P.; Guembe, M. The classification of Staphylococcus aureus strains by biofilm production differs depending on the method used. Enfermedades Infecc. Y Microbiol. Clínica 2022, 40, 134–137. [Google Scholar] [CrossRef]
- Abdullahi, U.F.; Igwenagu, E.; Mu’azu, A.; Aliyu, S.; Umar, M.I. Intrigues of biofilm: A perspective in veterinary medicine. Vet. World 2016, 9, 12–18. [Google Scholar] [CrossRef]
- Nesse, L.L.; Osland, A.M.; Vestby, L.K. The Role of Biofilms in the Pathogenesis of Animal Bacterial Infections. Microorganisms 2023, 11, 608. [Google Scholar] [CrossRef]
- Zhai, Y.; Minnick, P.J.; Pribis, J.P.; Garcia-Villada, L.; Hastings, P.J.; Herman, C.; Rosenberg, S.M. ppGpp and RNA-polymerase backtracking guide antibiotic-induced mutable gambler cells. Mol. Cell 2023, 83, 1298–1310.e4. [Google Scholar] [CrossRef]
- Sato, A.; Yamaguchi, T.; Hamada, M.; Ono, D.; Sonoda, S.; Oshiro, T.; Nagashima, M.; Kato, K.; Okazumi, S.; Katoh, R.; et al. Morphological and Biological Characteristics of Staphylococcus aureus Biofilm Formed in the Presence of Plasma. Microb. Drug Resist. 2019, 25, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Mah, T.F.; O’Toole, G.A. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.; Cho, C.Y.; Ho, A.; Huang, J.Y.; Martin, B.; Gilbert, E.S. 4-Ethoxybenzoic acid inhibits Staphylococcus aureus biofilm formation and potentiates biofilm sensitivity to vancomycin. Int. J. Antimicrob. Agents 2020, 56, 106086. [Google Scholar] [CrossRef] [PubMed]
- Frassinetti, S.; Falleni, A.; Del Carratore, R. Effect of itraconazole on Staphylococcus aureus biofilm and extracellular vesicles formation. Microb. Pathog. 2020, 147, 104267. [Google Scholar] [CrossRef] [PubMed]
- Gries, C.M.; Biddle, T.; Bose, J.L.; Kielian, T.; Lo, D.D. Staphylococcus aureus Fibronectin Binding Protein A Mediates Biofilm Development and Infection. Infect. Immun. 2020, 88, e00859-19. [Google Scholar] [CrossRef]
- Li, Y.; Xiao, P.; Wang, Y.; Hao, Y. Mechanisms and Control Measures of Mature Biofilm Resistance to Antimicrobial Agents in the Clinical Context. ACS Omega 2020, 5, 22684–22690. [Google Scholar] [CrossRef]
- Kwiecinski, J.M.; Jacobsson, G.; Horswill, A.R.; Josefsson, E.; Jin, T. Biofilm formation by Staphylococcus aureus clinical isolates correlates with the infection type. Infect. Dis. 2019, 51, 446–451. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, W.; Zhu, X.; Zhao, H.; Lu, Y.; Zhang, C.; Lu, Z. Surfactin effectively inhibits Staphylococcus aureus adhesion and biofilm formation on surfaces. Appl. Microbiol. Biotechnol. 2019, 103, 4565–4574. [Google Scholar] [CrossRef] [PubMed]
- Idrees, M.; Sawant, S.; Karodia, N.; Rahman, A. Staphylococcus aureus Biofilm: Morphology, Genetics, Pathogenesis and Treatment Strategies. Int. J. Env. Res. Public. Health 2021, 18, 7602. [Google Scholar] [CrossRef] [PubMed]
- Kamble, E.; Pardesi, K. Antibiotic Tolerance in Biofilm and Stationary-Phase Planktonic Cells of Staphylococcus aureus. Microb. Drug Resist. 2021, 27, 3–12. [Google Scholar] [CrossRef]
- Kaźmierczak, N.; Grygorcewicz, B.; Piechowicz, L. Biofilm Formation and Prevalence of Biofilm-Related Genes Among Clinical Strains of Multidrug-Resistant Staphylococcus aureus. Microb. Drug Resist. 2021, 27, 956–964. [Google Scholar] [CrossRef] [PubMed]
- Keogh, R.A.; Zapf, R.L.; Frey, A.; Marino, E.C.; Null, G.G.; Wiemels, R.E.; Holzschu, D.L.; Shaw, L.N.; Carroll, R.K. Staphylococcus aureus Trigger Factor Is Involved in Biofilm Formation and Cooperates with the Chaperone PpiB. J. Bacteriol. 2021, 203, e00681-20. [Google Scholar] [CrossRef]
- Pusparajah, P.; Letchumanan, V.; Law, J.W.; Ab Mutalib, N.S.; Ong, Y.S.; Goh, B.H.; Tan, L.T.; Lee, L.H. Streptomyces sp.-A Treasure Trove of Weapons to Combat Methicillin-Resistant Staphylococcus aureus Biofilm Associated with Biomedical Devices. Int. J. Mol. Sci. 2021, 22, 9360. [Google Scholar] [CrossRef]
- Qian, Y.; Xia, L.; Wei, L.; Li, D.; Jiang, W. Artesunate inhibits Staphylococcus aureus biofilm formation by reducing alpha-toxin synthesis. Arch. Microbiol. 2021, 203, 707–717. [Google Scholar] [CrossRef]
- Vudhya Gowrisankar, Y.; Manne Mudhu, S.; Pasupuleti, S.K.; Suthi, S.; Chaudhury, A.; Sarma, P. Staphylococcus aureus grown in anaerobic conditions exhibits elevated glutamine biosynthesis and biofilm units. Can. J. Microbiol. 2021, 67, 323–331. [Google Scholar] [CrossRef]
- Hirakawa, H.; Kurushima, J.; Hashimoto, Y.; Tomita, H. Progress Overview of Bacterial Two-Component Regulatory Systems as Potential Targets for Antimicrobial Chemotherapy. Antibiotics 2020, 9, 635. [Google Scholar] [CrossRef]
- Memmi, G.; Nair, D.R.; Cheung, A. Role of ArlRS in autolysis in methicillin-sensitive and methicillin-resistant Staphylococcus aureus strains. J. Bacteriol. 2012, 194, 759–767. [Google Scholar] [CrossRef]
- Ji, Y.; Yu, C.; Liang, X. Transcriptomic analysis of ArlRS two-component signaling regulon, a global regulator, in Staphylococcus aureus. Methods Enzym. 2007, 423, 502–513. [Google Scholar] [CrossRef]
- Luong, T.T.; Lee, C.Y. The arl locus positively regulates Staphylococcus aureus type 5 capsule via an mgrA-dependent pathway. Microbiology 2006, 152, 3123–3131. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zheng, L.; Landwehr, C.; Lunsford, D.; Holmes, D.; Ji, Y. Global regulation of gene expression by ArlRS, a two-component signal transduction regulatory system of Staphylococcus aureus. J. Bacteriol. 2005, 187, 5486–5492. [Google Scholar] [CrossRef] [PubMed]
- Fournier, B.; Klier, A. Protein A gene expression is regulated by DNA supercoiling which is modified by the ArlS-ArlR two-component system of Staphylococcus aureus. Microbiology 2004, 150, 3807–3819. [Google Scholar] [CrossRef] [PubMed]
- Fournier, B.; Hooper, D.C. A new two-component regulatory system involved in adhesion, autolysis, and extracellular proteolytic activity of Staphylococcus aureus. J. Bacteriol. 2000, 182, 3955–3964. [Google Scholar] [CrossRef] [PubMed]
- Fournier, B.; Klier, A.; Rapoport, G. The two-component system ArlS-ArlR is a regulator of virulence gene expression in Staphylococcus aureus. Mol. Microbiol. 2001, 41, 247–261. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, J.; Jiang, J.; Hu, J.; Xu, T.; Wang, J.; Qu, D. Role of the two-component regulatory system arlRS in ica operon and aap positive but non-biofilm-forming Staphylococcus epidermidis isolates from hospitalized patients. Microb. Pathog. 2014, 76, 89–98. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, J.; Xu, T.; Liu, J.; Yu, W.; Lou, Q.; Zhu, T.; He, N.; Ben, H.; Hu, J.; et al. The two-component signal transduction system ArlRS regulates Staphylococcus epidermidis biofilm formation in an ica-dependent manner. PLoS ONE 2012, 7, e40041. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Jiang, Q.; Fang, B.; Sun, B. The ArlR-MgrA regulatory cascade regulates PIA-dependent and protein-mediated biofilm formation in Rbf-dependent and Rbf-independent pathways. Int. J. Med. Microbiol. 2019, 309, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Burgui, S.; Gil, C.; Solano, C.; Lasa, I.; Valle, J. A Systematic Evaluation of the Two-Component Systems Network Reveals That ArlRS Is a Key Regulator of Catheter Colonization by Staphylococcus aureus. Front. Microbiol. 2018, 9, 342. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Zhu, X.; Zhao, K.; Yan, Y.; Xu, T.; Wang, J.; Zheng, J.; Huang, W.; Shi, L.; Shang, Y.; et al. The role of ArlRS in regulating oxacillin susceptibility in methicillin-resistant Staphylococcus aureus indicates it is a potential target for antimicrobial resistance breakers. Emerg. Microbes Infect. 2019, 8, 503–515. [Google Scholar] [CrossRef]
- Theis, T.J.; Daubert, T.A.; Kluthe, K.E.; Brodd, K.L.; Nuxoll, A.S. Staphylococcus aureus persisters are associated with reduced clearance in a catheter-associated biofilm infection. Front. Cell Infect. Microbiol. 2023, 13, 1178526. [Google Scholar] [CrossRef] [PubMed]
- Long, D.R.; Penewit, K.; Lo, H.Y.; Almazan, J.; Holmes, E.A.; Bryan, A.B.; Wolter, D.J.; Lewis, J.D.; Waalkes, A.; Salipante, S.J. In Vitro Selection Identifies Staphylococcus aureus Genes Influencing Biofilm Formation. Infect. Immun. 2023, 91, e0053822. [Google Scholar] [CrossRef]
- Bem, A.E.; Velikova, N.; Pellicer, M.T.; Baarlen, P.; Marina, A.; Wells, J.M. Bacterial histidine kinases as novel antibacterial drug targets. ACS Chem. Biol. 2015, 10, 213–224. [Google Scholar] [CrossRef]
- Sato, T.; Uno, T.; Kawamura, M.; Fujimura, S. In vitro Tolerability of Biofilm-Forming Trimethoprim-/Sulfamethoxazole-Resistant Small Colony Variants of Staphylococcus aureus Against Various Antimicrobial Agents. Microb. Drug Resist. 2021, 27, 1282–1289. [Google Scholar] [CrossRef]
- Shukla, S.K.; Rao, T.S. Targeting hydrophobicity in biofilm-associated protein (Bap) as a novel antibiofilm strategy against Staphylococcus aureus biofilm. Biophys. Chem. 2022, 289, 106860. [Google Scholar] [CrossRef]
- Yang, Y.; Yuan, L.; Li, J.; Muhammad, I.; Cheng, P.; Xiao, T.; Zhang, X. Preparation and evaluation of tilmicosin microspheres and lung-targeting studies in rabbits. Vet. J. 2019, 246, 27–34. [Google Scholar] [CrossRef]
- Roberts, R.B.; Chung, M.; de Lencastre, H.; Hargrave, J.; Tomasz, A.; Nicolau, D.P.; John, J.F., Jr.; Korzeniowski, O. Distribution of methicillin-resistant Staphylococcus aureus clones among health care facilities in Connecticut, New Jersey, and Pennsylvania. Microb. Drug Resist. 2000, 6, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, B.E.; Martinez-Aguilar, G.; Hulten, K.G.; Hammerman, W.A.; Coss-Bu, J.; Avalos-Mishaan, A.; Mason, E.O., Jr.; Kaplan, S.L. Severe Staphylococcal sepsis in adolescents in the era of community-acquired methicillin-resistant Staphylococcus aureus. Pediatrics 2005, 115, 642–648. [Google Scholar] [CrossRef] [PubMed]
- Diep, B.A.; Gill, S.R.; Chang, R.F.; Phan, T.H.; Chen, J.H.; Davidson, M.G.; Lin, F.; Lin, J.; Carleton, H.A.; Mongodin, E.F.; et al. Complete genome sequence of USA300, an epidemic clone of community-acquired meticillin-resistant Staphylococcus aureus. Lancet 2006, 367, 731–739. [Google Scholar] [CrossRef]
- Galac, M.R.; Stam, J.; Maybank, R.; Hinkle, M.; Mack, D.; Rohde, H.; Roth, A.L.; Fey, P.D. Complete Genome Sequence of Staphylococcus epidermidis 1457. Genome Announc 2017, 5, e00450-17. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Bae, T.; Schneewind, O.; Takeuchi, F.; Hiramatsu, K. Genome sequence of Staphylococcus aureus strain Newman and comparative analysis of staphylococcal genomes: Polymorphism and evolution of two major pathogenicity islands. J. Bacteriol. 2008, 190, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Charpentier, E.; Anton, A.I.; Barry, P.; Alfonso, B.; Fang, Y.; Novick, R.P. Novel cassette-based shuttle vector system for gram-positive bacteria. Appl. Environ. Microbiol. 2004, 70, 6076–6085. [Google Scholar] [CrossRef]
- Pang, Y.Y.; Schwartz, J.; Thoendel, M.; Ackermann, L.W.; Horswill, A.R.; Nauseef, W.M. agr-Dependent interactions of Staphylococcus aureus USA300 with human polymorphonuclear neutrophils. J. Innate Immun. 2010, 2, 546–559. [Google Scholar] [CrossRef]
- Li, M.; Yu, J.; Guo, G.; Shen, H. Interactions between Macrophages and Biofilm during Staphylococcus aureus-Associated Implant Infection: Difficulties and Solutions. J. Innate Immun. 2023, 15, 499–515. [Google Scholar] [CrossRef]
- Ghosh, C.; Das, M.C.; Acharjee, S.; Bhattacharjee, S.; Sandhu, P.; Kumari, M.; Bhowmik, J.; Ghosh, R.; Banerjee, B.; De, U.C.; et al. Combating Staphylococcus aureus biofilm formation: The inhibitory potential of tormentic acid and 23-hydroxycorosolic acid. Arch. Microbiol. 2023, 206, 25. [Google Scholar] [CrossRef]
- Bae, T.; Schneewind, O. Allelic replacement in Staphylococcus aureus with inducible counter-selection. Plasmid 2006, 55, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Xu, T.; Zhu, T.; Lou, Q.; Wang, X.; Wu, Y.; Huang, R.; Liu, J.; Liu, H.; Yu, F.; et al. Monoclonal antibodies against accumulation-associated protein affect EPS biosynthesis and enhance bacterial accumulation of Staphylococcus epidermidis. PLoS ONE 2011, 6, e20918. [Google Scholar] [CrossRef]
- Bai, X.; Shen, Y.; Zhang, T.; Meng, R.; Zhang, Y.; Deng, Y.; Guo, N. Anti-biofilm activity of biochanin A against Staphylococcus aureus. Appl. Microbiol. Biotechnol. 2023, 107, 867–879. [Google Scholar] [CrossRef] [PubMed]
- Demirci, M.; Yigin, A.; Demir, C. Efficacy of antimicrobial peptide LL-37 against biofilm forming Staphylococcus aureus strains obtained from chronic wound infections. Microb. Pathog. 2022, 162, 105368. [Google Scholar] [CrossRef]
- Dutta, K.; Karmakar, A.; Jana, D.; Ballav, S.; Shityakov, S.; Panda, A.K.; Ghosh, C. Benzyl isocyanate isolated from the leaves of Psidium guajava inhibits Staphylococcus aureus biofilm formation. Biofouling 2020, 36, 1000–1017. [Google Scholar] [CrossRef] [PubMed]
- Gonik, E.; Rodríguez Sartori, D.; David Gara, P.; Miñán, A.; Fernández Lorenzo de Mele, M.; Gonzalez, M.C. Staphylococcus aureus biofilm eradication by the synergistic effect exerted by PEG-coated silicon dots immobilized in silica films and light irradiation. Nanotechnology 2021, 32, 095105. [Google Scholar] [CrossRef] [PubMed]
- Alabdullatif, M.; Alzahrani, A. Expression of biofilm-associated genes in Staphylococcus aureus during storage of platelet concentrates. Transfus. Apher. Sci. 2022, 61, 103456. [Google Scholar] [CrossRef]
- Zheng, J.; Shang, Y.; Wu, Y.; Wu, J.; Chen, J.; Wang, Z.; Sun, X.; Xu, G.; Deng, Q.; Qu, D.; et al. Diclazuril Inhibits Biofilm Formation and Hemolysis of Staphylococcus aureus. ACS Infect. Dis. 2021, 7, 1690–1701. [Google Scholar] [CrossRef]
- Khasawneh, A.I.; Himsawi, N.; Abu-Raideh, J.; Salameh, M.A.; Al-Tamimi, M.; Al Haj Mahmoud, S.; Saleh, T. Status of Biofilm-Forming Genes among Jordanian Nasal Carriers of Methicillin-Sensitive and Methicillin-Resistant Staphylococcus aureus. Iran. Biomed. J. 2020, 24, 386–398. [Google Scholar] [CrossRef]
- Kim, Y.J.; Yu, H.H.; Park, Y.J.; Lee, N.K.; Paik, H.D. Anti-Biofilm Activity of Cell-Free Supernatant of Saccharomyces cerevisiae against Staphylococcus aureus. J. Microbiol. Biotechnol. 2020, 30, 1854–1861. [Google Scholar] [CrossRef]
- Kırmusaoğlu, S.; Kaşıkçı, H. Identification of ica-dependent biofilm production by Staphylococcus aureus clinical isolates and antibiofilm effects of ascorbic acid against biofilm production. J. Clin. Pathol. 2020, 73, 261–266. [Google Scholar] [CrossRef]
- Babushkina, I.V.; Mamonova, I.A.; Ulyanov, V.Y.; Gladkova, E.V.; Shpinyak, S.P. Antibiotic Susceptibility of Staphylococcus aureus Plankton and Biofilm Forms Isolated in Implant-Associated Infection. Bull. Exp. Biol. Med. 2021, 172, 46–48. [Google Scholar] [CrossRef]
- Chen, A.; Dellos-Nolan, S.; Lu, Y.; West, J.S.; Wozniak, D.J.; Mitton-Fry, M.J. Dioxane-Linked Novel Bacterial Topoisomerase Inhibitors Exhibit Bactericidal Activity against Planktonic and Biofilm Staphylococcus aureus In Vitro. Microbiol. Spectr. 2022, 10, e0205622. [Google Scholar] [CrossRef] [PubMed]
- Dash, P.; Rana, K.; Turuk, J.; Palo, S.K.; Pati, S. Antimicrobial Resistance and Biofilm Formation of Staphylococcus aureus Isolates from Febrile Cases: Findings from a Rural Cohort of Odisha, India. Pol. J. Microbiol. 2023, 72, 209–214. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, D.; Chang, J.; Yan, L.; Zhao, F.; Wu, Y.; Xu, T.; Gong, T.; Chen, L.; He, N.; et al. Efficacy of novel antibacterial compounds targeting histidine kinase YycG protein. Appl. Microbiol. Biotechnol. 2014, 98, 6003–6013. [Google Scholar] [CrossRef]
- Martinez-Irujo, J.J.; Villahermosa, M.L.; Alberdi, E.; Santiago, E. A checkerboard method to evaluate interactions between drugs. Biochem. Pharmacol. 1996, 51, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Kolenda, C.; Josse, J.; Medina, M.; Fevre, C.; Lustig, S.; Ferry, T.; Laurent, F. Evaluation of the Activity of a Combination of Three Bacteriophages Alone or in Association with Antibiotics on Staphylococcus aureus Embedded in Biofilm or Internalized in Osteoblasts. Antimicrob. Agents Chemother. 2020, 64, e02231-19. [Google Scholar] [CrossRef]
- Lin, Q.; Sun, H.; Yao, K.; Cai, J.; Ren, Y.; Chi, Y. The Prevalence, Antibiotic Resistance and Biofilm Formation of Staphylococcus aureus in Bulk Ready-To-Eat Foods. Biomolecules 2019, 9, 524. [Google Scholar] [CrossRef]
- Matias, R.R.; Sepúlveda, A.M.G.; Batista, B.N.; de Lucena, J.; Albuquerque, P.M. Degradation of Staphylococcus aureus Biofilm Using Hydrolytic Enzymes Produced by Amazonian Endophytic Fungi. Appl. Biochem. Biotechnol. 2021, 193, 2145–2161. [Google Scholar] [CrossRef] [PubMed]
- Moreno, A.; Dos Santos, D.M.; Lamartine de Moraes Melo Neto, C.; Luiz de Melo Moreno, A.; de Magalhães Bertoz, A.P.; Goiato, M.C. In vitro evaluation of the effect of different disinfectants on the biofilm of Staphylococcus epidermidis and Staphylococcus aureus formed on acrylic ocular prostheses. PLoS ONE 2020, 15, e0240116. [Google Scholar] [CrossRef]
- Orazi, G.; Ruoff, K.L.; O’Toole, G.A. Pseudomonas aeruginosa Increases the Sensitivity of Biofilm-Grown Staphylococcus aureus to Membrane-Targeting Antiseptics and Antibiotics. mBio 2019, 10, e01501-19. [Google Scholar] [CrossRef]
- Passos, T.F.; Souza, M.T.; Zanotto, E.D.; de Souza, C.W.O. Bactericidal activity and biofilm inhibition of F18 bioactive glass against Staphylococcus aureus. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 118, 111475. [Google Scholar] [CrossRef]
- Kwiecinski, J.M.; Kratofil, R.M.; Parlet, C.P.; Surewaard, B.G.J.; Kubes, P.; Horswill, A.R. Staphylococcus aureus uses the ArlRS and MgrA cascade to regulate immune evasion during skin infection. Cell Rep. 2021, 36, 109462. [Google Scholar] [CrossRef]
- Kwiecinski, J.M.; Crosby, H.A.; Valotteau, C.; Hippensteel, J.A.; Nayak, M.K.; Chauhan, A.K.; Schmidt, E.P.; Dufrene, Y.F.; Horswill, A.R. Staphylococcus aureus adhesion in endovascular infections is controlled by the ArlRS-MgrA signaling cascade. PLoS Pathog. 2019, 15, e1007800. [Google Scholar] [CrossRef] [PubMed]
- Crosby, H.A.; Schlievert, P.M.; Merriman, J.A.; King, J.M.; Salgado-Pabon, W.; Horswill, A.R. The Staphylococcus aureus Global Regulator MgrA Modulates Clumping and Virulence by Controlling Surface Protein Expression. PLoS Pathog. 2016, 12, e1005604. [Google Scholar] [CrossRef] [PubMed]
- Crosby, H.A.; Tiwari, N.; Kwiecinski, J.M.; Xu, Z.; Dykstra, A.; Jenul, C.; Fuentes, E.J.; Horswill, A.R. The Staphylococcus aureus ArlRS two-component system regulates virulence factor expression through MgrA. Mol. Microbiol. 2020, 113, 103–122. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Zhao, D.; Chang, J.; Liu, H.; Wang, X.; Zheng, J.; Huang, R.; Lin, Z.; Shang, Y.; Ye, L.; et al. Anti-bacterial and Anti-biofilm Evaluation of Thiazolopyrimidinone Derivatives Targeting the Histidine Kinase YycG Protein of Staphylococcus epidermidis. Front. Microbiol. 2017, 8, 549. [Google Scholar] [CrossRef]
- Cai, M.G.; Wu, Y.; Chang, J. Synthesis and biological evaluation of 2-arylimino-3-pyridin-thiazolineone derivatives as antibacterial agents. Bioorg Med. Chem. Lett. 2016, 26, 2517–2520. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, Y.; Zhao, D.; Gong, T.; Wu, Y.; Han, H.; Xu, T.; Peschel, A.; Han, S.; Qu, D. Antibacterial and anti-biofilm activities of thiazolidione derivatives against clinical staphylococcus strains. Emerg. Microbes Infect. 2015, 4, e1. [Google Scholar] [CrossRef]
- Zhao, D.; Liu, H.; Zheng, L.; He, G.; Qu, D.; Han, S. Synthesis of novel 4-thiazolidione derivatives as antibacterial agents against drug-resistant Staphylococcus epidermidis. Med. Chem. Res. 2012, 22, 3743–3750. [Google Scholar] [CrossRef]
- Huang, R.Z.; Zheng, L.K.; Liu, H.Y.; Pan, B.; Hu, J.; Zhu, T.; Wang, W.; Jiang, D.B.; Wu, Y.; Wu, Y.C.; et al. Thiazolidione derivatives targeting the histidine kinase YycG are effective against both planktonic and biofilm-associated Staphylococcus epidermidis. Acta Pharmacol. Sin. 2012, 33, 418–425. [Google Scholar] [CrossRef]
- Pan, B.; Huang, R.; Zheng, L.; Chen, C.; Han, S.; Qu, D.; Zhu, M.; Wei, P. Thiazolidione derivatives as novel antibiofilm agents: Design, synthesis, biological evaluation, and structure-activity relationships. Eur. J. Med. Chem. 2011, 46, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.; Huang, R.-Z.; Han, S.-Q.; Qu, D.; Zhu, M.-L.; Wei, P.; Ying, H.-J. Design, synthesis, and antibiofilm activity of 2-arylimino-3-aryl-thiazolidine-4-ones. Bioorganic Med. Chem. Lett. 2010, 20, 2461–2464. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.; Lee, B.; Yang, L.; Zhang, J.; Yang, X.; Qu, D.; Jiang, H.; Molin, S. Antimicrobial activities of YycG histidine kinase inhibitors against Staphylococcus epidermidis biofilms. FEMS Microbiol. Lett. 2007, 273, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, S.; Marsh, L.; Knight, K.; Ali, L.; Gomez, A.; Sunderhaus, A.; Abdel Aziz, M.H. NH125 Sensitizes Staphylococcus aureus to Cell Wall-Targeting Antibiotics through the Inhibition of the VraS Sensor Histidine Kinase. Microbiol. Spectr. 2023, 11, e0486122. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Quan, C.; Zhang, X.; Xiong, W.; Fan, S. Proteoliposome-based model for screening inhibitors targeting histidine kinase AgrC. Chem. Biol. Drug Des. 2019, 93, 712–723. [Google Scholar] [CrossRef]
- Trizna, E.Y.; Yarullina, M.N.; Baidamshina, D.R.; Mironova, A.V.; Akhatova, F.S.; Rozhina, E.V.; Fakhrullin, R.F.; Khabibrakhmanova, A.M.; Kurbangalieva, A.R.; Bogachev, M.I.; et al. Bidirectional alterations in antibiotics susceptibility in Staphylococcus aureus-Pseudomonas aeruginosa dual-species biofilm. Sci. Rep. 2020, 10, 14849. [Google Scholar] [CrossRef]
- Zhang, L.; Jin, M.; Sun, M. Inhibition characteristics of biofilm structure of Staphylococcus aureus. Cell. Mol. Biol. 2020, 66, 204–211. [Google Scholar] [CrossRef]
- Lyu, Z.; Shang, Y.; Wang, X.; Wu, Y.; Zheng, J.; Liu, H.; Gong, T.; Ye, L.; Qu, D. Monoclonal Antibodies Specific to the Extracellular Domain of Histidine Kinase YycG of Staphylococcus epidermidis Inhibit Biofilm Formation. Front. Microbiol. 2020, 11, 1839. [Google Scholar] [CrossRef]
- Gotoh, Y.; Doi, A.; Furuta, E.; Dubrac, S.; Ishizaki, Y.; Okada, M.; Igarashi, M.; Misawa, N.; Yoshikawa, H.; Okajima, T.; et al. Novel antibacterial compounds specifically targeting the essential WalR response regulator. J. Antibiot. 2010, 63, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Wei, Y.; Hou, S.; Lai, P.M.; Liu, H.; Tai, S.S.C.; Tang, V.Y.M.; Prakash, P.H.; Sze, K.H.; Chen, J.H.K.; et al. SaeR as a novel target for antivirulence therapy against Staphylococcus aureus. Emerg. Microbes Infect. 2023, 12, 2254415. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, M.; Roch, M.; Lasa, I.; Renzoni, A.; Kelley, W.L. The Role of ArlRS and VraSR in Regulating Ceftaroline Hypersusceptibility in Methicillin-Resistant Staphylococcus aureus. Antibiotics 2021, 10, 821. [Google Scholar] [CrossRef] [PubMed]
- Kwiecinski, J.M.; Jelani, D.A.; Fuentes, E.J.; Horswill, A.R. Therapeutic Inhibition of Staphylococcus aureus ArlRS Two-Component Regulatory System Blocks Virulence. Antimicrob. Agents Chemother. 2022, 66, e0018722. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Bhunia, A.K. Animal-Use Antibiotics Induce Cross-Resistance in Bacterial Pathogens to Human Therapeutic Antibiotics. Curr. Microbiol. 2019, 76, 1112–1117. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Staphylococcal Strains/Plasmids | Information |
|---|---|
| Strains | |
| FPR3757ΔarlRS | an arlRS knockout strain of the MRSA USA300 FPR3757 |
| NewmanΔarlRS | an arlRS knockout strain of the MSSA Newman |
| arlRSc (FPR3757) | an arlRS complementation strain of the FPR3757ΔarlRS |
| arlRSc (USA500) | an arlRS complementation strain of the USA500ΔarlRS |
| arlRSc (Newman) | an arlRS complementation strain of the NewmanΔarlRS |
| FPR3757-PmgrA-P2 | a mgrA promoter-reporter strain by transforming the USA300 FPR3757 with the plasmid pCM29-mgrA-P2 |
| FPR3757-Pspx-P2 | a spx promoter-reporter strain by transforming the USA300 FPR3757 with the plasmid pCM29-spx-P2 |
| ΔarlRS::PmgrA-P2 | FPR3757ΔarlRS harboring pCM-mgrA-P2 |
| ΔarlRS::Pspx-P2 | FPR3757ΔarlRS harboring pCM-spx-P2 |
| Plasmids | |
| pCM-spx-P2 | pCM29 modified by replacing its GFP promoter with spx P2 promoter |
| pCM-mgrA-P2 | pCM29 modified by replacing its GFP promoter with mgrA P2 promoter |
| pRB-500arlRS | pRB475 carrying the arlR and arlS genes of USA500 |
| pRB-NMarlRS | pRB475 carrying the arlR and arlS genes of Newman |
| pETMG-arlSHK | the arlS gene histidine kinase region cloned into pETMG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Wang, H.; Bai, J.; Cai, S.; Qu, D.; Xie, Y.; Wu, Y. The Staphylococcus aureus ArlS Kinase Inhibitor Tilmicosin Has Potent Anti-Biofilm Activity in Both Static and Flow Conditions. Microorganisms 2024, 12, 256. https://doi.org/10.3390/microorganisms12020256
Wang Z, Wang H, Bai J, Cai S, Qu D, Xie Y, Wu Y. The Staphylococcus aureus ArlS Kinase Inhibitor Tilmicosin Has Potent Anti-Biofilm Activity in Both Static and Flow Conditions. Microorganisms. 2024; 12(2):256. https://doi.org/10.3390/microorganisms12020256
Chicago/Turabian StyleWang, Zihui, Haoran Wang, Jinna Bai, Shen Cai, Di Qu, Youhua Xie, and Yang Wu. 2024. "The Staphylococcus aureus ArlS Kinase Inhibitor Tilmicosin Has Potent Anti-Biofilm Activity in Both Static and Flow Conditions" Microorganisms 12, no. 2: 256. https://doi.org/10.3390/microorganisms12020256
APA StyleWang, Z., Wang, H., Bai, J., Cai, S., Qu, D., Xie, Y., & Wu, Y. (2024). The Staphylococcus aureus ArlS Kinase Inhibitor Tilmicosin Has Potent Anti-Biofilm Activity in Both Static and Flow Conditions. Microorganisms, 12(2), 256. https://doi.org/10.3390/microorganisms12020256