
The Probiotic Properties and Safety of Limosilactobacillus mucosae NK41 and Bifidobacterium longum NK46


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Cultivation Conditions
2.2. Cell Culture
2.3. Resistance to Simulated Gastrointestinal Environments
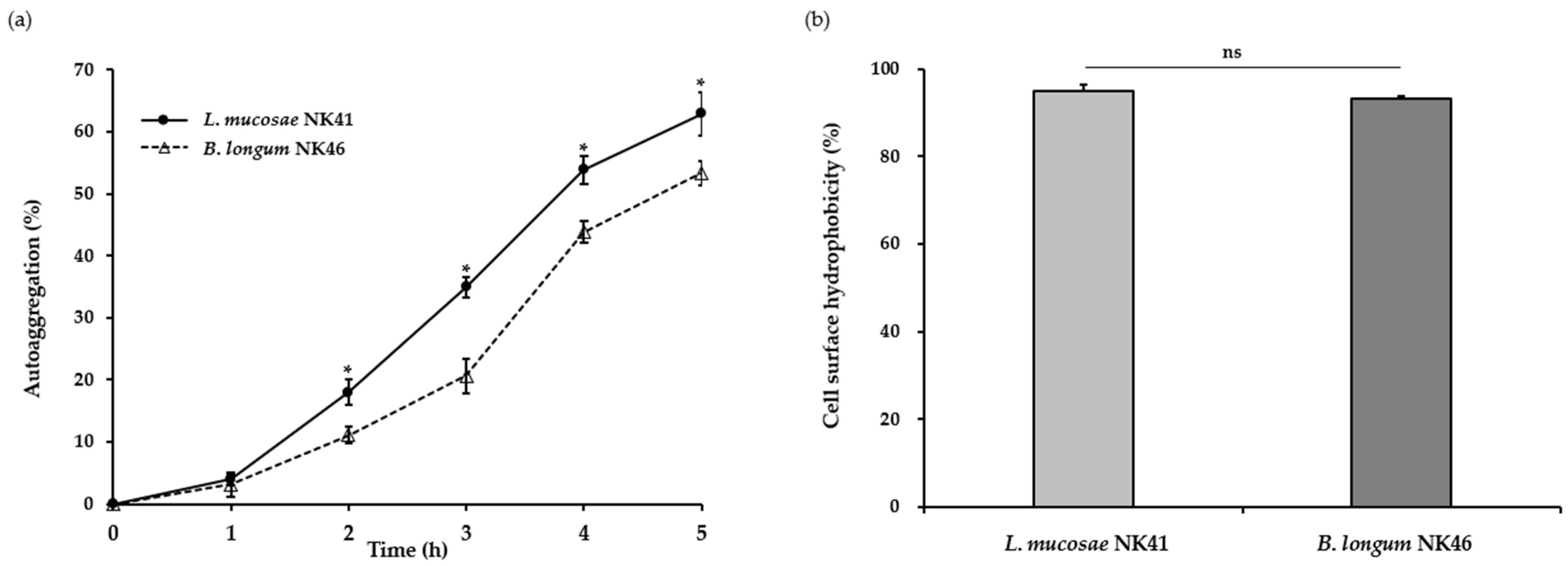
2.4. Auto-Aggregation
2.5. Cell Surface Hydrophobicity
2.6. Cell Cytotoxicity Assay
2.7. Adhesion to Caco-2 Cell Line
2.8. Enzymatic Activity Profile
2.9. Hemolytic Activity
2.10. Gelatinase Activity
2.11. Determination of Minimum Inhibitory Concentration
2.12. Determination of D(−)-Lactate and L(+)-Lactate
2.13. Detection of Genes for Virulence Factors Production
2.14. Statistical Analysis
3. Results and Discussion
3.1. Stability of the Probiotic Strains under Simulated Gastrointestinal Conditions
3.2. Auto-Aggregation and Cell Surface Hydrophobicity of the Probiotic Strains
3.3. Adhesion Ability of the Probiotic Strains
3.4. Profile of Enzyme Activities of the Probiotic Strains
3.5. Hemolytic and Gelatinase Activities of L. mucosae NK41 and B. longum NK46
3.6. Antibiotic Susceptibility of the Probiotic Strains
3.7. Lactate Production of the Probiotic Strains
3.8. Detection of Virulence Factor Genes in the Probiotic Strains
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kechagia, M.; Basoulis, D.; Konstantopoulou, S.; Dimitriadi, D.; Gyftopoulou, K.; Skarmoutsou, N.; Fakiri, E.M. Health benefits of probiotics: A review. ISRN Nutr. 2013, 2013, 481651. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Lee, A.; Yoo, H.J.; Kim, M.; Noh, G.M.; Lee, J.H. Supplementation with the probiotic strain Weissella cibaria JW15 enhances natural killer cell activity in nondiabetic subjects. J. Funct. Foods 2018, 48, 153–158. [Google Scholar] [CrossRef]
- Kang, M.G.; Han, S.W.; Kang, H.R.; Hong, S.J.; Kim, D.H.; Choi, J.H. Probiotic NVP-1703 Alleviates Allergic Rhinitis by inducing IL-10 Expression: A Four-week Clinical Trial. Nutrients 2020, 12, 1427. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.E.; Kim, M.S.; Shim, K.W.; Kim, Y.-I.; Chu, J.; Kim, B.-K.; Choi, I.S.; Kim, J.Y. Effects of Lactobacillus plantarum Q180 on postprandial lipid levels and intestinal environment: A double-blind, randomized, placebo-controlled, parallel trial. Nutrients 2020, 12, 255. [Google Scholar] [CrossRef] [PubMed]
- Lim, E.Y.; Lee, S.-Y.; Shin, H.S.; Lee, J.; Nam, Y.-D.; Lee, D.O.; Lee, J.Y.; Yeon, S.H.; Son, R.H.; Park, C.L. The effect of Lactobacillus acidophilus YT1 (MENOLACTO) on improving menopausal symptoms: A randomized, double-blinded, placebo-controlled clinical trial. J. Clin. Med. 2020, 9, 2173. [Google Scholar] [CrossRef]
- Sharpe, M.; Shah, V.; Freire-Lizama, T.; Cates, E.C.; McGrath, K.; David, I.; Cowan, S.; Letkeman, J.; Stewart-Wilson, E. Effectiveness of oral intake of Lactobacillus rhamnosus GR-1 and Lactobacillus reuteri RC-14 on Group B Streptococcus colonization during pregnancy: A midwifery-led double-blind randomized controlled pilot trial. J. Matern.-Fetal Neonatal Med. 2021, 34, 1814–1821. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.-P.; Lee, K.-M.; Kang, J.-H.; Yun, S.-I.; Park, H.-O.; Moon, Y.; Kim, J.-Y. Effect of Lactobacillus gasseri BNR17 on overweight and obese adults: A randomized, double-blind clinical trial. Korean J. Fam. Med. 2013, 34, 80–89. [Google Scholar] [CrossRef]
- Ng, S.Y.; Koon, S.S.; Padam, B.S.; Chye, F.Y. Evaluation of probiotic potential of lactic acid bacteria isolated from traditional Malaysian fermented Bambangan (Mangifera pajang). CYTA J. Food 2015, 13, 563–572. [Google Scholar] [CrossRef]
- Araya, M.; Morelli, L.; Reid, G.; Sanders, M.; Stanton, C.; Pineiro, M.; Ben Embarek, P. Joint FAO/WHO Working Group Report on Drafting Guidelines for the Evaluation of Probiotics in Food; World Health Organization, Food and Agriculture Organization of the United Nations: London, ON, Canada, 2002. [Google Scholar]
- Lee, B.S.; Ban, O.-H.; Bang, W.Y.; Chae, S.A.; Oh, S.; Park, C.; Lee, M.; Kim, S.-J.; Yang, J.; Jung, Y.H. Safety assessment of Lactobacillus reuteri IDCC 3701 based on phenotypic and genomic analysis. Ann. Microbiol. 2021, 71, 10. [Google Scholar] [CrossRef]
- Royan, M.; Seighalani, R.; Mortezaei, F.; Pourebrahim, M. In vitro assessment of safety and functional probiotic properties of Lactobacillus mucosae strains isolated from Iranian native ruminants intestine. Ital. J. Anim. Sci. 2021, 20, 1187–1200. [Google Scholar] [CrossRef]
- Somashekaraiah, R.; Shruthi, B.; Deepthi, B.; Sreenivasa, M. Probiotic properties of lactic acid bacteria isolated from neera: A naturally fermenting coconut palm nectar. Front. Microbiol. 2019, 10, 1382. [Google Scholar] [CrossRef]
- Monteagudo-Mera, A.; Rastall, R.A.; Gibson, G.R.; Charalampopoulos, D.; Chatzifragkou, A. Adhesion mechanisms mediated by probiotics and prebiotics and their potential impact on human health. Appl. Microbiol. Biotechnol. 2019, 103, 6463–6472. [Google Scholar] [CrossRef] [PubMed]
- Plavec, T.V.; Berlec, A. Safety aspects of genetically modified lactic acid bacteria. Microorganisms 2020, 8, 297. [Google Scholar] [CrossRef] [PubMed]
- Kamaliya, D.; Javia, B.; Gadhvi, M.; Dudhagara, D. Isolation and characterization of probiotic bacteria from fruit pulp, screening of probiotic properties and production of bacteriocin and probiotic curd. Open J. Biol. Sci. 2023, 8, 033–042. [Google Scholar] [CrossRef]
- Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Hilbert, F. Update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA 15: Suitability of taxonomic units notified to EFSA until September 2021. EFSA J. 2022, 20, e07045. [Google Scholar] [CrossRef]
- Kwon, Y.J.; Chun, B.H.; Jung, H.S.; Chu, J.; Joung, H.; Park, S.Y.; Kim, B.K.; Jeon, C.O. Safety assessment of Lactiplantibacillus (formerly Lactobacillus) plantarum Q180. J. Microbiol. Biotechnol. 2021, 31, 1420–1429. [Google Scholar] [CrossRef]
- Perin, L.M.; Miranda, R.O.; Todorov, S.D.; de Melo Franco, B.D.G.; Nero, L.A. Virulence, antibiotic resistance and biogenic amines of bacteriocinogenic lactococci and enterococci isolated from goat milk. Int. J. Food Microbiol. 2014, 185, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Alayande, K.A.; Aiyegoro, O.A.; Nengwekhulu, T.M.; Katata-Seru, L.; Ateba, C.N. Integrated genome-based probiotic relevance and safety evaluation of Lactobacillus reuteri PNW1. PLoS ONE 2020, 15, e0235873. [Google Scholar] [CrossRef]
- Han, S.-K.; Kim, D.H. Lactobacillus mucosae and Bifidobacterium longum synergistically alleviate immobilization stress-induced anxiety/depression in mice by suppressing gut dysbiosis. J. Microbiol. Biotechnol. 2019, 29, 1369–1374. [Google Scholar] [CrossRef]
- Ma, X.; Kim, J.-K.; Shin, Y.-J.; Son, Y.-H.; Lee, D.-Y.; Park, H.-S.; Kim, D.-H. Alleviation of cognitive impairment-like behaviors, neuroinflammation, colitis, and gut dysbiosis in 5xFAD transgenic and aged mice by Lactobacillus mucosae and Bifidobacterium longum. Nutrients 2023, 15, 3381. [Google Scholar] [CrossRef]
- Lee, J.; Kim, S.; Kang, C.-H. Screening and probiotic properties of lactic acid bacteria with potential immunostimulatory activity isolated from kimchi. Fermentation 2022, 9, 4. [Google Scholar] [CrossRef]
- Rastogi, S.; Mittal, V.; Singh, A. In vitro evaluation of probiotic potential and safety assessment of Lactobacillus mucosae strains isolated from donkey’s lactation. Probiotics Antimicrob. Proteins 2020, 12, 1045–1056. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, H.C.; de Sousa Melo, D.; Ramos, C.L.; Dias, D.R.; Schwan, R.F. Probiotic properties of lactobacilli and their ability to inhibit the adhesion of enteropathogenic bacteria to Caco-2 and HT-29 cells. Probiotics Antimicrob. Proteins 2021, 13, 102–112. [Google Scholar] [CrossRef]
- Veljović, K.; Popović, N.; Miljković, M.; Tolinački, M.; Terzić-Vidojević, A.; Kojić, M. Novel aggregation promoting factor AggE contributes to the probiotic properties of Enterococcus faecium BGGO9-28. Front. Microbiol. 2017, 8, 1843. [Google Scholar] [CrossRef]
- Clinical Laboratory Standards Institute (CLSI). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 11th ed.; Approved Standard; CLSI Document M07-A; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Rychen, G.; Aquilina, G.; Azimonti, G.; Bampidis, V.; de Lourdes Bastos, M.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; Flachowsky, G. Guidance on the characterisation of microorganisms used as feed additives or as production organisms. EFSA J. 2018, 16, e05206. [Google Scholar] [CrossRef] [PubMed]
- Bengoa, A.A.; Zavala, L.; Carasi, P.; Trejo, S.A.; Bronsoms, S.; de los Ángeles Serradell, M.; Garrote, G.L.; Abraham, A.G. Simulated gastrointestinal conditions increase adhesion ability of Lactobacillus paracasei strains isolated from kefir to Caco-2 cells and mucin. Food Res. Int. 2018, 103, 462–467. [Google Scholar] [CrossRef] [PubMed]
- de Moraes, G.M.D.; de Abreu, L.R.; do Egito, A.S.; Salles, H.O.; da Silva, L.M.F.; Nero, L.A.; Todorov, S.D.; Dos Santos, K.M.O. Functional properties of Lactobacillus mucosae strains isolated from Brazilian goat milk. Probiotics Antimicrob. Proteins 2017, 9, 235–245. [Google Scholar] [CrossRef]
- Fajardo-Cavazos, P.; Nicholson, W.L. Shelf life and simulated gastrointestinal tract survival of selected commercial probiotics during a simulated round-trip journey to Mars. Front. Microbiol. 2021, 12, 748950. [Google Scholar] [CrossRef]
- da Silva, M.N.; Tagliapietra, B.L.; do Amaral Flores, V.; dos Santos Richards, N.S.P. In vitro test to evaluate survival in the gastrointestinal tract of commercial probiotics. Curr. Res. Food Sci. 2021, 4, 320–325. [Google Scholar] [CrossRef]
- Stasiak-Różańska, L.; Berthold-Pluta, A.; Pluta, A.S.; Dasiewicz, K.; Garbowska, M. Effect of simulated gastrointestinal tract conditions on survivability of probiotic bacteria present in commercial preparations. Int. J. Environ. Res. Public Health 2021, 18, 1108. [Google Scholar] [CrossRef]
- Darmastuti, A.; Hasan, P.N.; Wikandari, R.; Utami, T.; Rahayu, E.S.; Suroto, D.A. Adhesion properties of Lactobacillus plantarum Dad-13 and Lactobacillus plantarum Mut-7 on Sprague Dawley rat intestine. Microorganisms 2021, 9, 2336. [Google Scholar] [CrossRef]
- García-Cayuela, T.; Korany, A.M.; Bustos, I.; de Cadiñanos, L.P.G.; Requena, T.; Peláez, C.; Martínez-Cuesta, M.C. Adhesion abilities of dairy Lactobacillus plantarum strains showing an aggregation phenotype. Food Res. Int. 2014, 57, 44–50. [Google Scholar] [CrossRef]
- Tuo, Y.; Yu, H.; Ai, L.; Wu, Z.; Guo, B.; Chen, W. Aggregation and adhesion properties of 22 Lactobacillus strains. J. Dairy Sci. 2013, 96, 4252–4257. [Google Scholar] [CrossRef] [PubMed]
- Krausova, G.; Hyrslova, I.; Hynstova, I. In vitro evaluation of adhesion capacity, hydrophobicity, and auto-aggregation of newly isolated potential probiotic strains. Fermentation 2019, 5, 100. [Google Scholar] [CrossRef]
- Purkayastha, S.D.; Bhattacharya, M.K.; Prasad, H.K.; Bhattacharjee, M.J.; De Mandal, S.; Mathipi, V.; Kumar, N.S. Probiotic and Cytotoxic Potential of Vaginal Lactobacillus Isolated from Healthy Northeast Indian Women. J. Pure Appl. Microbiol. 2020, 14, 205–214. [Google Scholar] [CrossRef]
- Xu, H.; Jeong, H.; Lee, H.; Ahn, J. Assessment of cell surface properties and adhesion potential of selected probiotic strains. Lett. Appl. Microbiol. 2009, 49, 434–442. [Google Scholar] [CrossRef]
- Li, M.; Wang, Y.; Cui, H.; Li, Y.; Sun, Y.; Qiu, H.-J. Characterization of lactic acid bacteria isolated from the gastrointestinal tract of a wild boar as potential probiotics. Front. Vet. Sci. 2020, 7, 49. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Kang, C.-H. Probiotics alleviate oxidative stress in H2O2-exposed hepatocytes and t-BHP-induced C57BL/6 mice. Microorganisms 2022, 10, 234. [Google Scholar] [CrossRef]
- Patrone, V.; Al-Surrayai, T.; Romaniello, F.; Fontana, A.; Milani, G.; Sagheddu, V.; Puglisi, E.; Callegari, M.L.; Al-Mansour, H.; Kishk, M.W. Integrated phenotypic-genotypic analysis of candidate probiotic Weissella cibaria strains isolated from dairy cows in Kuwait. Probiotics Antimicrob. Proteins 2021, 13, 809–823. [Google Scholar] [CrossRef] [PubMed]
- Begunova, A.V.; Rozhkova, I.V.; Glazunova, O.A.; Moiseenko, K.V.; Savinova, O.S.; Fedorova, T.V. Fermentation profile and probiotic-related characteristics of Bifidobacterium longum MC-42. Fermentation 2021, 7, 101. [Google Scholar] [CrossRef]
- Sulemankhil, I.; Parent, M.; Jones, M.L.; Feng, Z.; Labbé, A.; Prakash, S. In vitro and in vivo characterization and strain safety of Lactobacillus reuteri NCIMB 30253 for probiotic applications. Can. J. Microbiol. 2012, 58, 776–787. [Google Scholar] [CrossRef]
- Bang, W.Y.; Chae, S.A.; Ban, O.-H.; Oh, S.; Park, C.; Lee, M.; Shin, M.; Yang, J.; Jung, Y.H. The in vitro and in vivo Safety Evaluation of Lactobacillus acidophilus IDCC 3302. Food Microbiol. 2021, 49, 39–44. [Google Scholar] [CrossRef]
- Gueimonde, M.; Sánchez, B.G.; de los Reyes-Gavilán, C.; Margolles, A. Antibiotic resistance in probiotic bacteria. Front. Microbiol. 2013, 4, 202. [Google Scholar] [CrossRef]
- Astó, E.; Huedo, P.; Altadill, T.; Aguilo Garcia, M.; Sticco, M.; Perez, M.; Espadaler-Mazo, J. Probiotic properties of Bifidobacterium longum KABP042 and Pediococcus pentosaceus KABP041 show potential to counteract functional gastrointestinal disorders in an observational pilot trial in infants. Front. Microbiol. 2022, 12, 741391. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.; Paek, J.J.; Lee, Y. Antimicrobial Resistance of Seventy Lactic Acid Bacteria Isolated from Commercial Probiotics in Korea. J. Microbiol. Biotechnol. 2023, 33, 500. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Tomar, S.K.; Goswami, P.; Sangwan, V.; Singh, R. Antibiotic resistance among commercially available probiotics. Food Res. Int. 2014, 57, 176–195. [Google Scholar] [CrossRef]
- Choi, I.Y.; Kim, J.; Kim, S.-H.; Ban, O.-H.; Yang, J.; Park, M.-K. Safety evaluation of Bifidobacterium breve IDCC4401 isolated from infant feces for use as a commercial probiotic. J. Microbiol. Biotechnol. 2021, 31, 949–955. [Google Scholar] [CrossRef] [PubMed]
- Song, K.-Y. Preliminary data on the ratio of D (–)-lactate and L (+)-lactate levels in various lactic acid bacteria as evaluated using an enzymatic method. J. Dairy Sci. Biotechnol. 2022, 40, 15–22. [Google Scholar] [CrossRef]
- Lee, Y.-R.; Bang, W.Y.; Baek, K.-R.; Kim, G.-H.; Kang, M.-J.; Yang, J.; Seo, S.-O. Safety evaluation by phenotypic and genomic characterization of four Lactobacilli strains with probiotic properties. Microorganisms 2022, 10, 2218. [Google Scholar] [CrossRef] [PubMed]
- Moreno Muñoz, J.A.; Chenoll, E.; Casinos, B.; Bataller, E.; Ramón, D.; Genovés, S.; Montava, R.; Ribes, J.M.; Buesa, J.; Fabrega, J. Novel probiotic Bifidobacterium longum subsp. infantis CECT 7210 strain active against rotavirus infections. Appl. Environ. Microbiol. 2011, 77, 8775–8783. [Google Scholar] [CrossRef] [PubMed]
- FDA. GRAS Notice Inventory. GRN No. 856. Available online: https://www.fda.gov/food/generally-recognized-safe-gras/gras-notice-inventory (accessed on 10 August 2023).
- Kiruthiga, A.; Padmavathy, K.; Shabana, P.; Naveenkumar, V.; Gnanadesikan, S.; Malaiyan, J. Improved detection of esp, hyl, asa1, gelE, cylA virulence genes among clinical isolates of Enterococci. BMC Res. Notes 2020, 13, 170. [Google Scholar] [CrossRef]
- Todorov, S.D.; Perin, L.M.; Carneiro, B.M.; Rahal, P.; Holzapfel, W.; Nero, L.A. Safety of Lactobacillus plantarum ST8Sh and its bacteriocin. Probiotics Antimicrob. Proteins 2017, 9, 334–344. [Google Scholar] [CrossRef] [PubMed]
- Chajęcka-Wierzchowska, W.; Zadernowska, A.; Łaniewska-Trokenheim, Ł. Virulence factors of Enterococcus spp. presented in food. LWT Food Sci. Technol. 2017, 75, 670–676. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Virulence Factor | Primers (5′ to 3′) | Product Size (bp) | References |
|---|---|---|---|---|
| gelE | Gelatinase | TATGACAATGCTTTTTGGGAT AGATGCACCCGAAATAATATA | 213 | [11] |
| ace | Adhesion of collagen | GAATTGACAAAAGTTCAATCG GTCTGTCTTTTCACTTGTTTC | 1008 | |
| efaA | Endocarditis antigen | GCCAATTGGGACAGACCCTC CGCCTTCTGTTCCTTCTTTGGC | 688 | |
| esp | Enterococcal surface protein | AGATTTCATCTTTGATTCTTGG AATTGATTCTTTAGCATCTGG | 510 |
| Strains | Initial Counts (log CFU/mL) | SGF a (log CFU/mL) | SIF b (log CFU/mL) | Survival Rate (%) |
|---|---|---|---|---|
| L. mucosae NK41 | 7.65 ± 0.04 | 5.97 ± 0.03 * | 5.46 ± 0.08 * | 71.45 ± 0.76 |
| B. longum NK46 | 7.84 ± 0.04 | 3.24 ± 0.11 * | 3.20 ± 0.13 * | 40.82 ± 1.41 |
| Enzyme | L. mucosae NK41 | B. longum NK46 |
|---|---|---|
| Control (Negative) | 0 | 0 |
| Alkaline phosphatase | 0 | 0 |
| Esterase (C4) | 3 | 2 |
| Esterase Lipase (C8) | 2 | 1 |
| Lipase (C14) | 0 | 0 |
| Leucine arylamidase | 3 | 4 |
| Valine arylamidase | 0 | 0 |
| Cystine arylamidase | 0 | 0 |
| Trypsin | 0 | 0 |
| α-Chymotrypsin | 0 | 0 |
| Acid phosphatase | 1 | 0 |
| Naphtol-AS-BI-phosphohydrolase | 2 | 2 |
| α-Galactosidase | 4 | 5 |
| β-Galactosidase | 5 | 5 |
| β-Glucuronidase | 0 | 0 |
| α-Glucosidase | 2 | 4 |
| β-Glucosidase | 5 | 0 |
| N-acetyl-β-glucosaminidase | 0 | 0 |
| α-Mannosidase | 0 | 0 |
| α-Fucosidase | 0 | 0 |
| Antibiotics a | L. mucosae NK41 | B. longum NK46 | ||||
|---|---|---|---|---|---|---|
| Cut-Off Value (μg/mL) | MIC b (μg/mL) | Susceptibility (S/R) | Cut-Off Value (μg/mL) | MIC (μg/mL) | Susceptibility (S/R) | |
| AMP | 2 | 0.5 | S | 2 | 0.125 | S |
| VAN | n.r. c | 256 | - | 2 | 0.5 | S |
| GEN | 16 | 8 | S | 64 | 16 | S |
| KAN | 64 | 64 | S | n.r. | 64 | - |
| STR | 64 | 64 | S | 128 | 8 | S |
| ERY | 1 | <0.125 | S | 1 | <0.125 | S |
| CLI | 4 | 0.125 | S | 1 | 0.125 | S |
| TET | 8 | 4 | S | 8 | 2 | S |
| CHL | 4 | 4 | S | 4 | 0.5 | S |
| Strains | D-lactate (mM) | L-lactate (mM) | Ratio of Isomers (%) | |
|---|---|---|---|---|
| D-Form | L-Form | |||
| L. mucosae NK41 | 6.13 ± 0.23 | 15.62 ± 1.08 | 28.20 ± 0.68 | 71.80 ± 0.68 |
| B. longum NK46 | 0.43 ± 0.09 | 19.22 ± 0.50 | 2.20 ± 0.40 | 97.80 ± 0.40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.; Jo, J.; Seo, H.; Han, S.-W.; Kim, D.-H. The Probiotic Properties and Safety of Limosilactobacillus mucosae NK41 and Bifidobacterium longum NK46. Microorganisms 2024, 12, 776. https://doi.org/10.3390/microorganisms12040776
Lee J, Jo J, Seo H, Han S-W, Kim D-H. The Probiotic Properties and Safety of Limosilactobacillus mucosae NK41 and Bifidobacterium longum NK46. Microorganisms. 2024; 12(4):776. https://doi.org/10.3390/microorganisms12040776
Chicago/Turabian StyleLee, Jaekoo, Jaehyun Jo, Hanseul Seo, Seung-Won Han, and Dong-Hyun Kim. 2024. "The Probiotic Properties and Safety of Limosilactobacillus mucosae NK41 and Bifidobacterium longum NK46" Microorganisms 12, no. 4: 776. https://doi.org/10.3390/microorganisms12040776
APA StyleLee, J., Jo, J., Seo, H., Han, S. -W., & Kim, D. -H. (2024). The Probiotic Properties and Safety of Limosilactobacillus mucosae NK41 and Bifidobacterium longum NK46. Microorganisms, 12(4), 776. https://doi.org/10.3390/microorganisms12040776