
Insights on Klebsiella pneumoniae Biofilms Assembled on Different Surfaces Using Phenotypic and Genotypic Approaches



Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Biofilms
2.2.1. Biofilm Assembly on Silicone
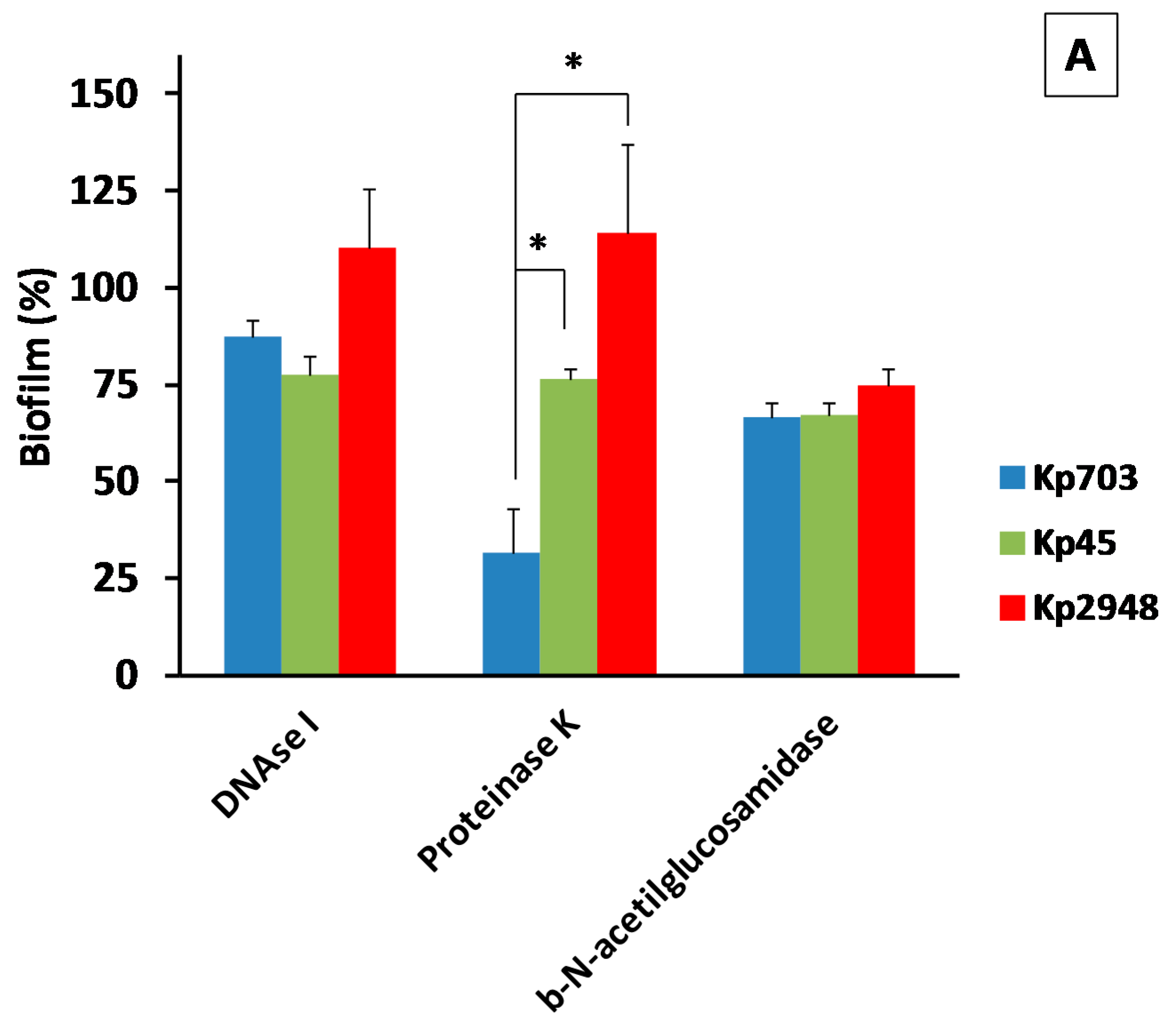
2.2.2. Effect of Enzymatic Treatment on Biofilm Formation
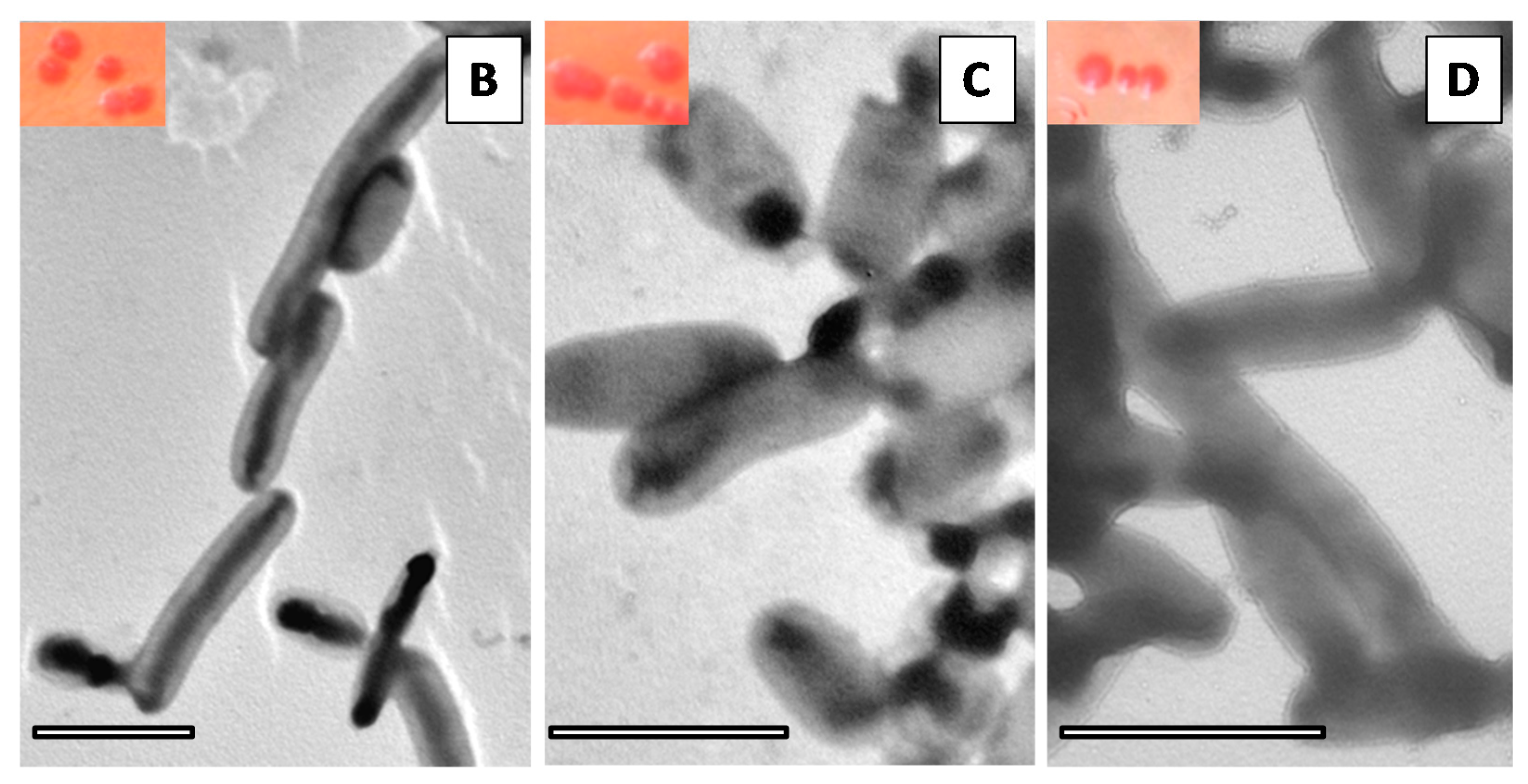
2.2.3. Congo Red-Base Colony Morphology and Transmission Electron Microscopy (TEM)
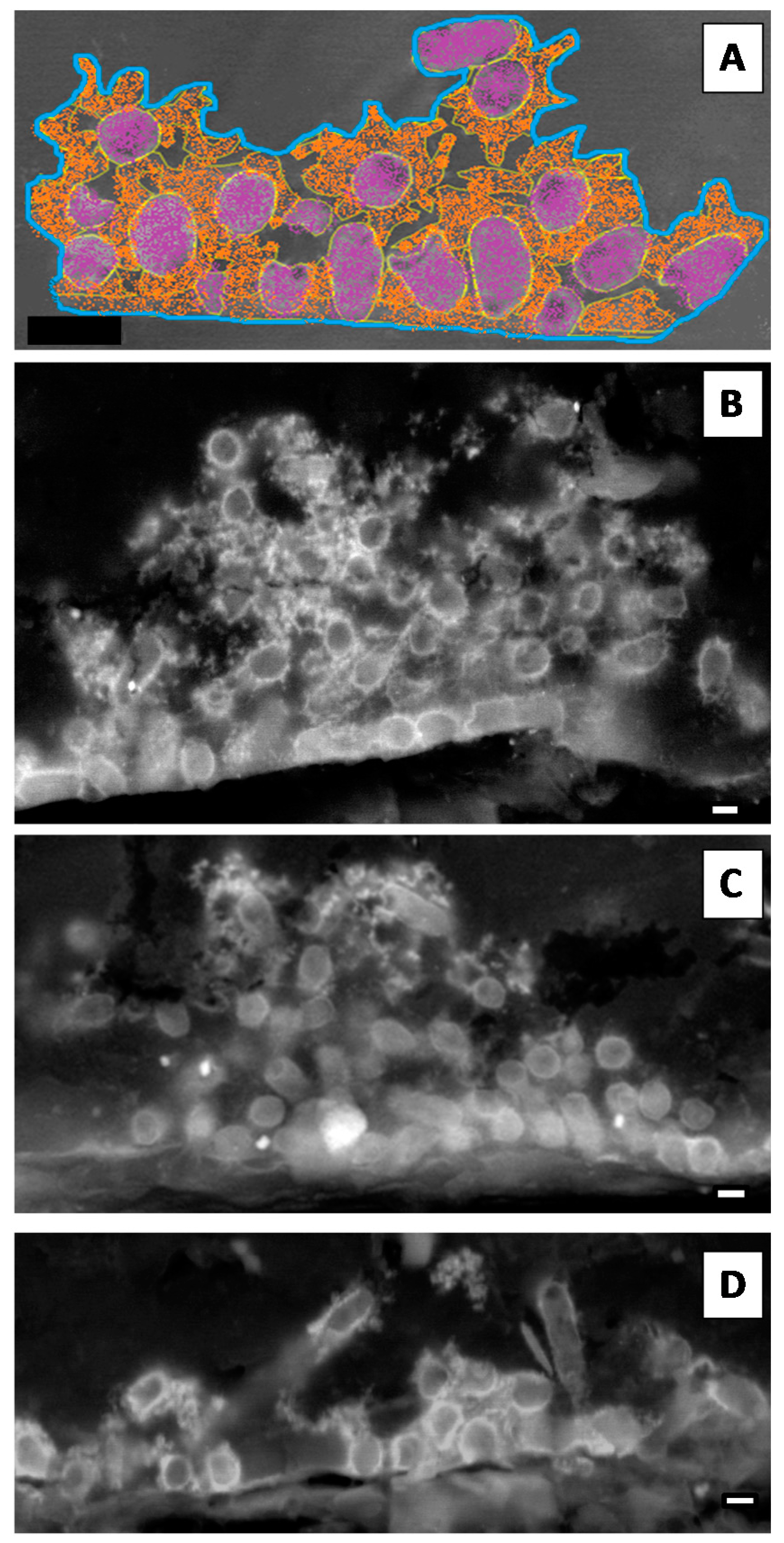
2.2.4. Scanning Laser Confocal Microscopy
2.2.5. Scanning Electron Microscopy (SEM)
2.3. Statistical Analysis
2.4. Whole-Genome Sequencing and Comparative Analyses
2.5. Nucleotide Sequence Accession Numbers
3. Results
3.1. Biofilm Assembly on Polystyrene and Silicone
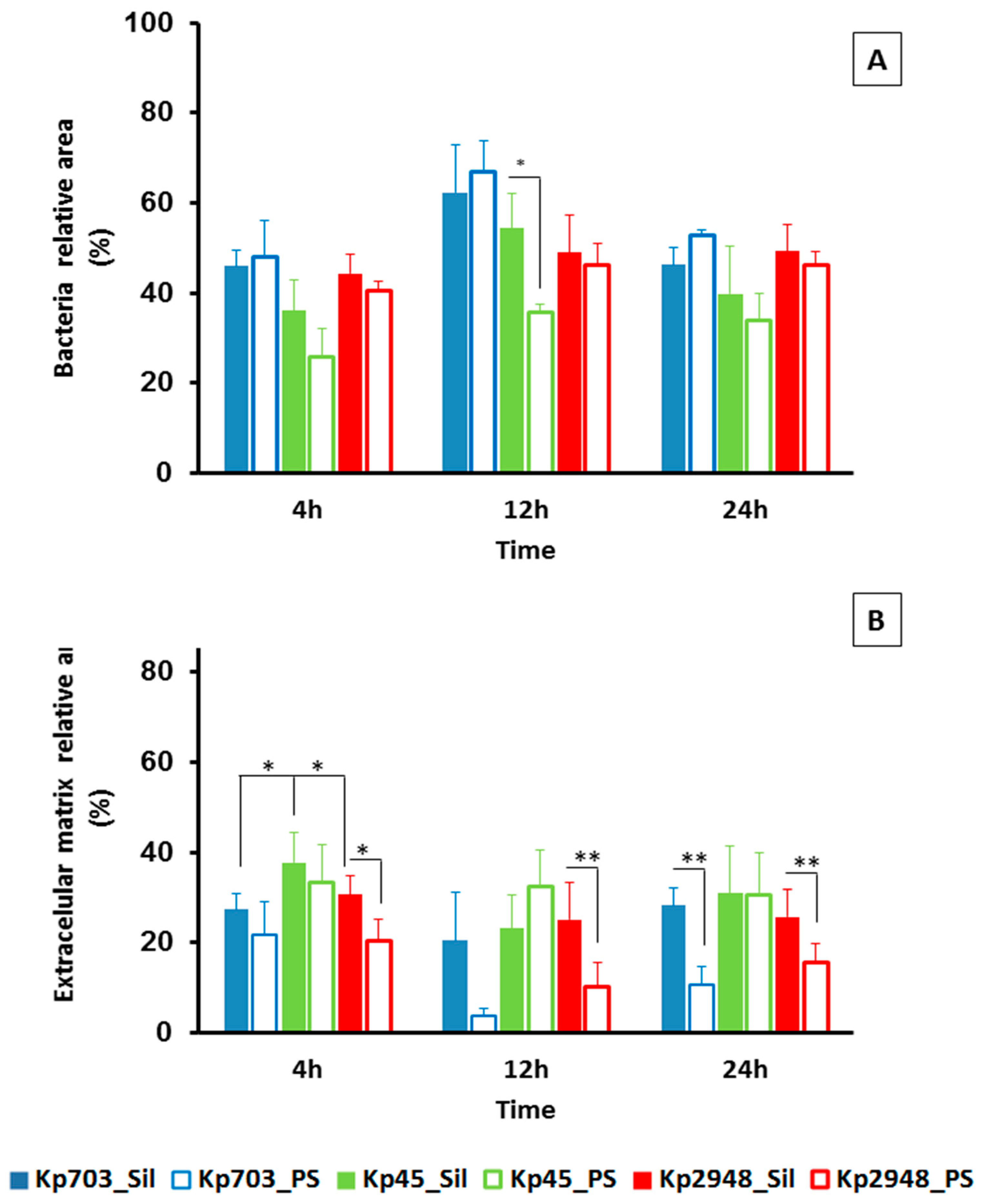
3.2. The Extracellular Polymeric Matrix (EPS)
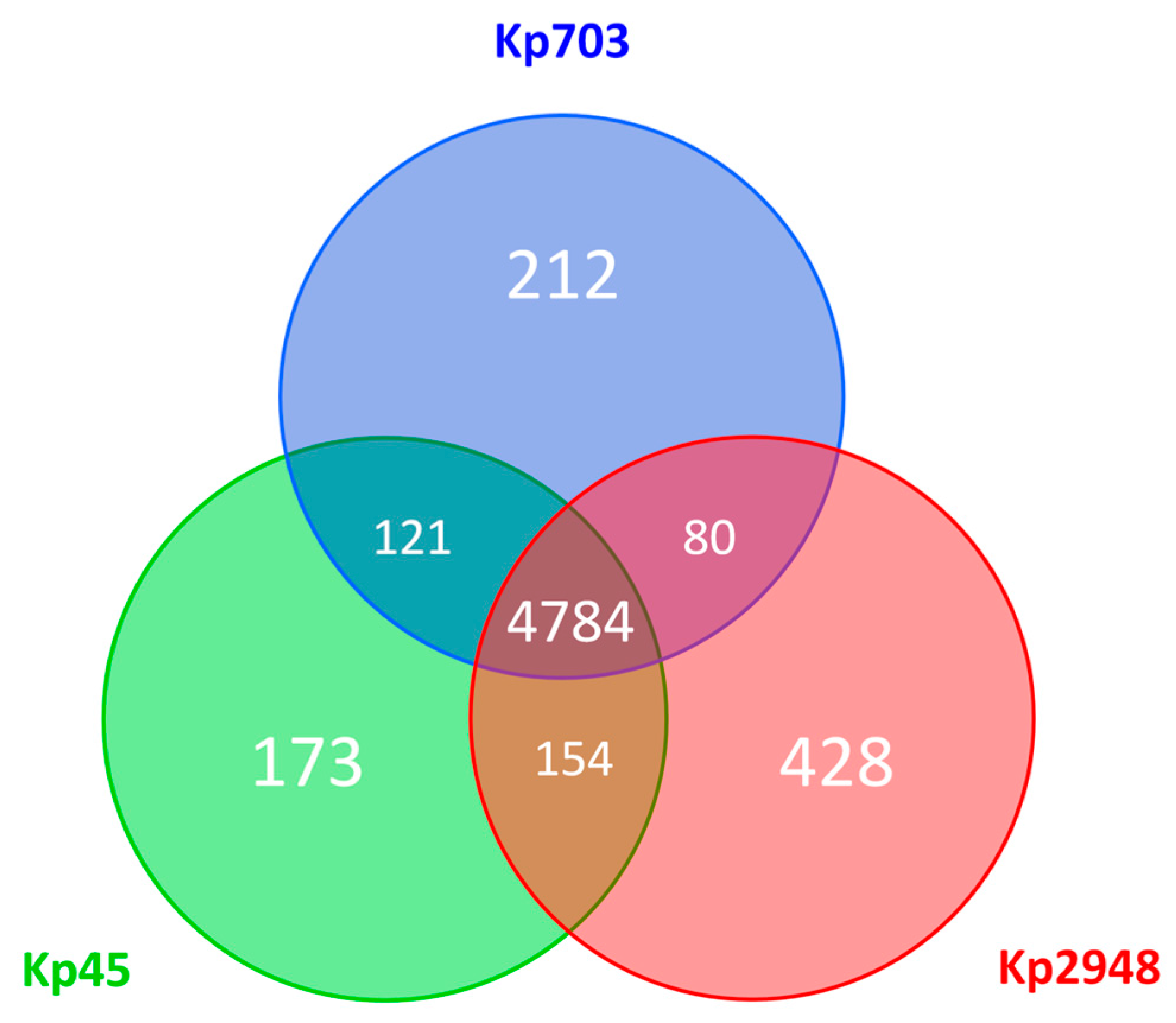
3.3. Whole-Genome Analysis of K. pneumoniae Isolates Displaying Different Biofilm Phenotypes
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Vuotto, C.; Longo, F.; Balice, M.P.; Donelli, G.; Varaldo, P.E. Antibiotic resistance related to biofilm formation in Klebsiella pneumoniae. Pathogens 2014, 3, 743–758. [Google Scholar] [CrossRef] [PubMed]
- Podschun, R.; Ullmann, U. Klebsiella spp. as nosocomial pathogens: Epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin. Microbiol. Rev. 1998, 11, 589–603. [Google Scholar] [PubMed]
- Percival, S.L.; Suleman, L.; Vuotto, C.; Donelli, G. Healthcare-associated infections, medical devices and biofilms: Risk, tolerance and control. J. Med. Microbiol. 2015, 64, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Machado, P.; Silva, A.; Lito, L.; Melo-Cristino, J.; Duarte, A. Emergence of Klebsiella pneumoniae ST11-producing KPC-3 carbapenemase at a Lisbon hospital. Clin. Microbiol. Infect. 2010, 16, S28. [Google Scholar]
- Francolini, I.; Donelli, G. Prevention and control of biofilm-based medical-device-related infections. FEMS Immunol. Med. Microbiol. 2010, 59, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Bryers, J.D. Medical biofilms. Biotechnol. Bioeng. 2008, 100, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Wilksch, J.J.; Yang, J.; Clements, A.; Gabbe, J.L.; Short, K.R.; Cao, H.; Cavaliere, R.; James, C.E.; Whitchurch, C.B.; Schembri, M.A.; et al. MrkH, a novel c-di-GMP-dependent transcriptional activator, controls Klebsiella pneumoniae biofilm formation by regulating type 3 fimbriae expression. PLoS Pathog. 2011, 7, e1002204. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.-H.; Chen, F.-J.; Huang, Y.-J.; Chen, S.-Y.; Liu, K.-L.; Wang, Z.-C.; Peng, H.-L.; Yew, T.-R.; Liu, C.-H.; Liou, G.-G.; et al. Identification of protein domains on major pilin MrkA that affects the mechanical properties of Klebsiella pneumoniae type 3 fimbriae. Langmuir 2012, 28, 7428–7435. [Google Scholar] [CrossRef] [PubMed]
- Bandeira, M.; Carvalho, P.A.; Duarte, A.; Jordao, L. Exploring dangerous connections between Klebsiella pneumoniae biofilms and healthcare-associated infections. Pathogens 2014, 3, 720–731. [Google Scholar] [CrossRef] [PubMed]
- Pomakova, D.; Hsiao, C.; Beanan, J.; Olson, R.; MacDonald, U.; Keynan, Y.; Russo, T. Clinical and phenotypic differences between classic and hypervirulent Klebsiella pneumoniae: An emerging and under-recognized pathogenic variant. Eur. J. Clin. Microbiol. 2012, 31, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Fodah, R.A.; Scott, J.B.; Tam, H.-H.; Yan, P.; Pfeffer, T.L.; Bundschuh, R.; Warawa, J.M. Correlation of Klebsiella pneumoniae Comparative Genetic Analyses with Virulence Profiles in a Murine Respiratory Disease Model. PLoS ONE 2014, 9, e107394. [Google Scholar] [CrossRef] [PubMed]
- Al-Fattani, M.A.; Douglas, L.J. Biofilm matrix of Candida albicans and Candida tropicalis: Chemical composition and role in drug resistance. J. Med. Microbiol. 2006, 55, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Charlebois, A.; Jacques, M.; Archambault, M. Biofilm formation of Clostridium perfringens and its exposure to low-dose antimicrobials. Front. Microbiol. 2014, 5, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yan, Y.; Rong, D.; Wang, J.; Wang, H.; Liu, Z.; Wang, J.; Yang, R.; Han, Y. Increased biofilm formation ability in Klebsiella pneumoniae after short-term exposure to a simulated microgravity environment. MicrobiologyOpen 2016, 5, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Bialek-Davenet, S.; Criscuolo, A.; Ailloud, F.; Passet, V.; Jones, L.; Delannoy-Vieillard, A.-S.; Garin, B.; Le Hello, S.; Arlet, G.; Nicolas-Chanoine, M.-H.; et al. Genomic definition of hypervirulent and multidrug-resistant Klebsiella pneumoniae clonal groups. Emerg. Infect. Dis. 2014, 20, 1812–1820. [Google Scholar] [CrossRef] [PubMed]
- Diancourt, L.; Passet, V.; Verhoef, J.; Grimont, P.A.; Brisse, S. Multilocus sequence typing of Klebsiella pneumoniae nosocomial isolates. J. Clin. Microbiol. 2005, 43, 4178–4182. [Google Scholar] [CrossRef] [PubMed]
- Holt, K.E.; Wertheim, H.; Zadoks, R.N.; Baker, S.; Whitehouse, C.A.; Dance, D.; Jenney, A.; Connor, T.R.; Hsu, L.Y.; Severin, J.; et al. Genomic analysis of diversity, population structure, virulence, and antimicrobial resistance in Klebsiella pneumoniae, an urgent threat to public health. Proc. Natl. Acad. Sci. USA 2015, 112, E3574–E3581. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.-J.; Lin, T.-L.; Chen, C.-T.; Chen, Y.-Y.; Hsieh, P.-F.; Hsu, C.-R.; Wu, M.-C.; Wang, J.-T. Genetic analysis of capsular polysaccharide synthesis gene clusters in 79 capsular types of Klebsiella spp. Sci. Rep. 2015, 5, 15573. [Google Scholar] [CrossRef] [PubMed]
- Hancock, V.; Witsø, I.L.; Klemm, P. Biofilm formation as a function of adhesin, growth medium, substratum and strain type. Int. J. Med. Microbiol. 2011, 301, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Van Laar, T.A.; Chen, T.; You, T.; Leung, K.P. Sublethal concentrations of carbapenems alter cell morphology and genomic expression of Klebsiella pneumoniae biofilms. Antimicrob. Agents Chemother. 2015, 59, 1707–1717. [Google Scholar] [CrossRef] [PubMed]
- Abee, T.; Kovács, Á.T.; Kuipers, O.P.; Van der Veen, S. Biofilm formation and dispersal in Gram-positive bacteria. Curr. Opin Biotechnol. 2011, 22, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Bandeira, M.; Carvalho, P.A.; Duarte, A.; Jordao, L. Bacterial biofilms, antibiotic resistance and healthcare-associated infections: A dangerous connection. Microsc. Microanal. 2015, 21, 38–39. [Google Scholar] [CrossRef]
- Donlan, R.M. Biofilms: Microbial life on surfaces. Emerg. Infect. Dis. 2002, 8, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, K.K. What drives bacteria to produce a biofilm? FEMS Microbiol. Lett. 2004, 236, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Koczan, J.M.; Lenneman, B.R.; McGrath, M.J.; Sundin, G.W. Cell surface attachment structures contribute to biofilm formation and xylem colonization by Erwinia amylovora. Appl. Environ. Microb. 2011, 77, 7031–7039. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed]
- Bales, P.M.; Renke, E.M.; May, S.L.; Shen, Y.; Nelson, D.C. Purification and characterization of biofilm-associated EPS exopolysaccharides from ESKAPE organisms and other pathogens. PLoS ONE 2013, 8, e67950. [Google Scholar] [CrossRef] [PubMed]
- Zogaj, X.; Bokranz, W.; Nimtz, M.; Römling, U. Production of cellulose and curli fimbriae by members of the family Enterobacteriaceae isolated from the human gastrointestinal tract. Infect. Immun. 2003, 71, 4151–4158. [Google Scholar] [CrossRef] [PubMed]
- Loferer, H.; Hammar, M.; Normark, S. Availability of the fibre subunit CsgA and the nucleator protein CsgB during assembly of fibronectin-binding curli is limited by the intracellular concentration of the novel lipoprotein CsgG. Mol. Microbiol. 1997, 26, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Römling, U.; Galperin, M.Y. Bacterial cellulose biosynthesis: Diversity of operons, subunits, products and functions. Trends Microbiol. 2015, 23, 545–557. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.N.; Mortensen, M.S.; Krogfelt, K.A.; Clegg, S. Role of Klebsiella pneumoniae type 1 and type 3 fimbriae in colonizing silicone tubes implanted into the bladders of mice as a model of catheter-associated urinary tract infections. Infect. Immun. 2013, 81, 3009–3017. [Google Scholar] [CrossRef] [PubMed]
- Duguid, J.P. Fimbriae and adhesive properties in Klebsiella strains. J. Gen. Microbiol. 1959, 21, 271–286. [Google Scholar] [CrossRef] [PubMed]
- Craig, L.; Pique, M.E.; Tainer, J.A. Type IV pilus structure and bacterial pathogenicity. Nat. Rev. Microbiol. 2004, 2, 363–378. [Google Scholar] [CrossRef] [PubMed]
- Guilhen, C.; Charbonnel, N.; Parisot, N.; Gueguen, N.; Iltis, A.; Forestier, C.; Balestrino, D. Transcriptional profiling of Klebsiella pneumoniae defines signatures for planktonic, sessile and biofilm-dispersed cells. BMC Genom. 2016, 17, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Astaneh, S.D.A.; Rasooli, I.; Gargari, S.L.M. The role of filamentous hemagglutinin adhesin in adherence and biofilm formation in Acinetobacter baumannii ATCC19606 T. Microb. Pathog. 2014, 74, 42–49. [Google Scholar] [CrossRef] [PubMed]
- García-Contreras, R.; Zhang, X.-S.; Kim, Y.; Wood, T.K. Protein translation and cell death: The role of rare tRNAs in biofilm formation and in activating dormant phage killer genes. PLoS ONE 2008, 3, e2394. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J.; Kim, S.C.; Kim, J.; Do, A.; Han, S.Y.; Lee, B.D.; Lee, H.H.; Lee, M.C.; Lee, S.H.; Oh, T.; et al. Synergistic inhibition of Streptococcal biofilm by ribose and xylitol. Arch. Oral Biol. 2015, 60, 304–312. [Google Scholar] [CrossRef] [PubMed]
- James, D.; Shao, H.; Lamont, R.J.; Demuth, D.R. The Actinobacillus actinomycetemcomitans ribose binding protein RbsB interacts with cognate and heterologous autoinducer 2 signals. Infect. Immun. 2006, 74, 4021–4029. [Google Scholar] [CrossRef] [PubMed]
- Calisto, F.; Caneiras, C.; Cerqueira, S.; Lito, L.; Melo-Cristino, J.; Duarte, A. Klebsiella pneumoniae producing carbapenemase KPC-3 identified in hospital wards. Rev. Port. Doença Infecc. 2012, 8, 127–134. [Google Scholar]
- Novais, A.; Rodrigues, C.; Branquinho, R.; Antunes, P.; Grosso, F.; Boaventura, L.; Ribeiro, G.; Peixe, L. Spread of an OmpK36-modified ST15 Klebsiella pneumoniae variant during an outbreak involving multiple carbapenem-resistant Enterobacteriaceae species and clones. Eur. J. Clin. Microbiol. 2012, 31, 3057–3063. [Google Scholar] [CrossRef] [PubMed]
- Österblad, M.; Kirveskari, J.; Hakanen, A.J.; Tissari, P.; Vaara, M.; Jalava, J. Carbapenemase-producing Enterobacteriaceae in Finland: The first years (2008–11). J. Antimicrob. Chemother. 2012, 67, 2860–2864. [Google Scholar] [CrossRef] [PubMed]
- Stillwell, T.; Green, M.; Barbadora, K.; Ferrelli, J.G.; Roberts, T.L.; Weissman, S.J.; Nowalk, A. Outbreak of KPC-3 producing carbapenem-resistant Klebsiella pneumoniae in a US pediatric hospital. J. Pediatr. Infect. Dis. Soc. 2014, 4, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Naparstek, L.; Carmeli, Y.; Navon-Venezia, S.; Banin, E. Biofilm formation and susceptibility to gentamicin and colistin of extremely drug-resistant KPC-producing Klebsiella pneumoniae. J. Antimicrob. Chemother. 2014, 69, 1027–1034. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genomic Region 1 | Approximate Length (Kbp) | Isolate | Relevant Information 2,3 | ||
|---|---|---|---|---|---|
| Kp703 | Kp45 | Kp2948 | |||
| 1 | 45 | √ | --- | --- | Intact prophage (predicted by PHAST) |
| 2 | 41 | √ | --- | --- | Intact prophage (predicted by PHAST) |
| 3 | 4 | √ | --- | --- | Includes predicted AMR genes: tetracycline resistance genes tet(A/G) and tetR |
| 4 | 6 | √ | --- | --- | Includes Ribose ABC-transporter system proteins (permease RbsC, ATP-binding RbsA) |
| 5 | 11 | √ | --- | --- | Includes genes encoding a Filamentous hemagglutinin and a Hemolysin transporter protein |
| 6 | 57 | √ | --- | --- | Includes genes encoding a Haemolysin expression modulating protein, the YdeA protein and a Programmed cell death antitoxin MazEF |
| 7 | 12 | √ | --- | --- | Putative prophage elements: predicted annotation includes several hypothetical proteins, a purine NTPase and a prophage CP4-57 integrase |
| 8 | 2 | √ | √ | --- | Aerotaxis sensor receptor protein |
| 9 | 13 | √ | √ | --- | Includes an additional Urea ABC transport system (another similar gene cluster is carried by all three isolates) |
| 10 4 | 36 | √ | --- | --- | Incomplete prophage (predicted by PHAST) carrying a strain-specific Glycosaminoglycan attachment site encoding gene |
| 11 4 | 36 | --- | √ | --- | Incomplete prophage (predicted by PHAST) carrying a strain-specific Transcription activator gutR gene |
| 12 | 12 | --- | √ | √ | Putative plasmid fragments carrying the virulence-associated proteins VagC and VagD |
| 13 | 37 | --- | √ | --- | Intact prophage (predicted by PHAST) |
| 14 | 43 | --- | √ | --- | Intact prophage (predicted by PHAST) |
| 15 | 57 | √ | --- | √ | Includes genes encoding Type IV pili-related proteins, a TonB-dependent protein, Iron aquisition yersiniabactin synthesis enzymes; and a predicted AMR gene likely associated with resistance to tetracycline and aminoglycosides (fyuA) |
| 16 | 2 | √ | --- | √ | Includes a predicted AMR gene: blaTEM-1A—class A beta-lactamase |
| 17 | 13 | --- | √ | √ | Includes genes encoding Type I secretion related proteins (LapC, ATPase LapB, LapE, agglutinin RTX) |
| 18 | 7 | --- | √ | √ | Includes predicted AMR genes: Aminoglycoside 3′-phosphotransferase strA and strB [or APH(3″)-Ib and APH(6)-Id, respectively] likely associated with resistance to aminoglycosides |
| 19 | 11 | --- | √ | --- | Predicted annotation includes several hypothetical proteins and putative phage-like elements. |
| 20 | 39 | --- | √ | --- | Intact prophage (predicted by PHAST) |
| 21 | 17 | --- | --- | √ | Questionable prophage (predicted by PHAST) |
| 22 | 11 | --- | --- | √ | Incomplete prophage (predicted by PHAST) |
| 23 | 40 | --- | --- | √ | Intact prophage (predicted by PHAST) |
| 24 | 25 | --- | --- | √ | Questionable prophage (predicted by PHAST) |
| 25 | 37 | --- | --- | √ | Questionable prophage (predicted by PHAST) |
| 26 | 12 | --- | --- | √ | Incomplete prophage (predicted by PHAST) |
| 27 | 5 | --- | --- | √ | Includes genes encoding a putative serine protease (from the Peptidase S8 Subtilisin superfamily) and a cell division protein FtsH |
| 28 | 27 | --- | --- | √ | Putative fragment of a conjugative plasmid including genes encoding Type IV pili-related proteins, a Programmed cell death antitoxin MazE and a zinc metalloproteinase Mpr protein |
| 29 | 1 | --- | --- | √ | Includes a predicted AMR gene: trimetroprim resistance gene dfrA14 coding for a Dihydrofolate reductase |
| 30 | 40 | --- | --- | √ | Intact prophage (predicted by PHAST) |
| 31 | 4 | --- | --- | √ | Includes predicted AMR genes: tetracycline resistance genes tet(A/G) and tetR; and proteins from the Glutathione-dependent pathway of formaldehyde detoxification |
| 32 | 17 | --- | --- | √ | Includes a predicted AMR gene (KPC-3 beta-lactam resistance gene) and gene encoding a Chromate resistance protein ChrB |
| 33 | 22 | --- | --- | √ | Includes genes coding for a arsenic resistance operon and a outer membrane protein or related peptidoglycan-associated (lipo)protein |
| 34 | 5 | --- | --- | √ | Includes genes coding for a phosphonate ABC transport system and the RuBisCO operon transcriptional regulator CbbR |
| 35 | 1 | --- | --- | √ | Includes a predicted AMR gene: SHV-161 beta-lactam resistance gene |
| 36 | 3 | --- | --- | √ | Includes a predicted AMR gene: AC(6′)-Ib putative fluoroquinolone resistance gene |
| 37 | 2 | --- | --- | √ | Includes a predicted AMR gene: putative Sulphonamide resistance gene sul1 coding for a Dihydropteroate synthase |
| 38 | 1 | --- | --- | √ | Includes a predicted AMR gene: putative Sulphonamide resistance gene sul2 coding for a Dihydropteroate synthase |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bandeira, M.; Borges, V.; Gomes, J.P.; Duarte, A.; Jordao, L. Insights on Klebsiella pneumoniae Biofilms Assembled on Different Surfaces Using Phenotypic and Genotypic Approaches. Microorganisms 2017, 5, 16. https://doi.org/10.3390/microorganisms5020016
Bandeira M, Borges V, Gomes JP, Duarte A, Jordao L. Insights on Klebsiella pneumoniae Biofilms Assembled on Different Surfaces Using Phenotypic and Genotypic Approaches. Microorganisms. 2017; 5(2):16. https://doi.org/10.3390/microorganisms5020016
Chicago/Turabian StyleBandeira, Maria, Vítor Borges, João P. Gomes, Aida Duarte, and Luisa Jordao. 2017. "Insights on Klebsiella pneumoniae Biofilms Assembled on Different Surfaces Using Phenotypic and Genotypic Approaches" Microorganisms 5, no. 2: 16. https://doi.org/10.3390/microorganisms5020016
APA StyleBandeira, M., Borges, V., Gomes, J. P., Duarte, A., & Jordao, L. (2017). Insights on Klebsiella pneumoniae Biofilms Assembled on Different Surfaces Using Phenotypic and Genotypic Approaches. Microorganisms, 5(2), 16. https://doi.org/10.3390/microorganisms5020016