Inoculation with Azospirillum sp. and Herbaspirillum sp. Bacteria Increases the Tolerance of Maize to Drought Stress

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Statistical Analysis
2.2. Bacterial Growth and Inoculation
2.3. Plant Growth Conditions
2.4. Measures
2.4.1. Symbiotic Development
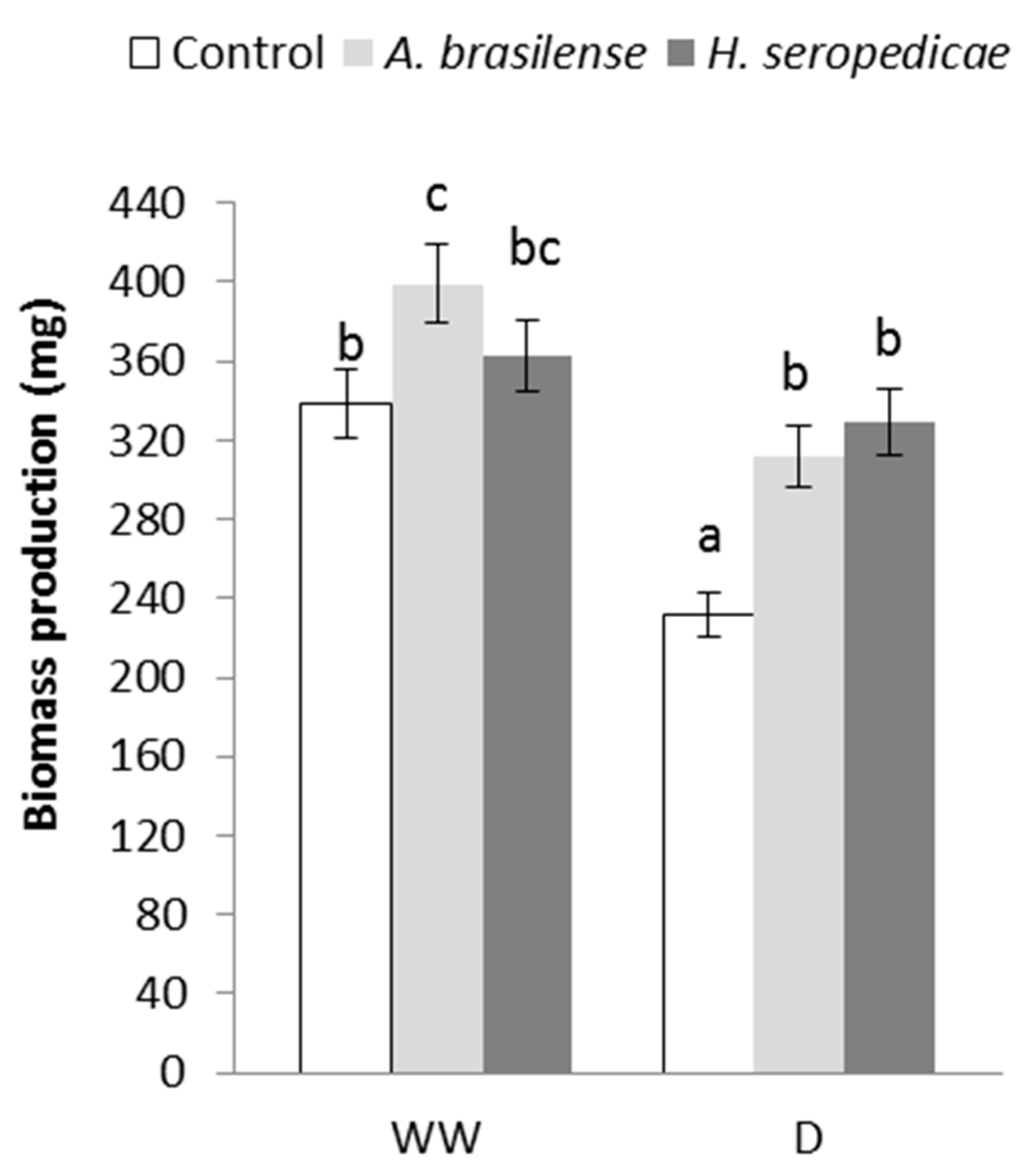
2.4.2. Total Biomass (TB) Production
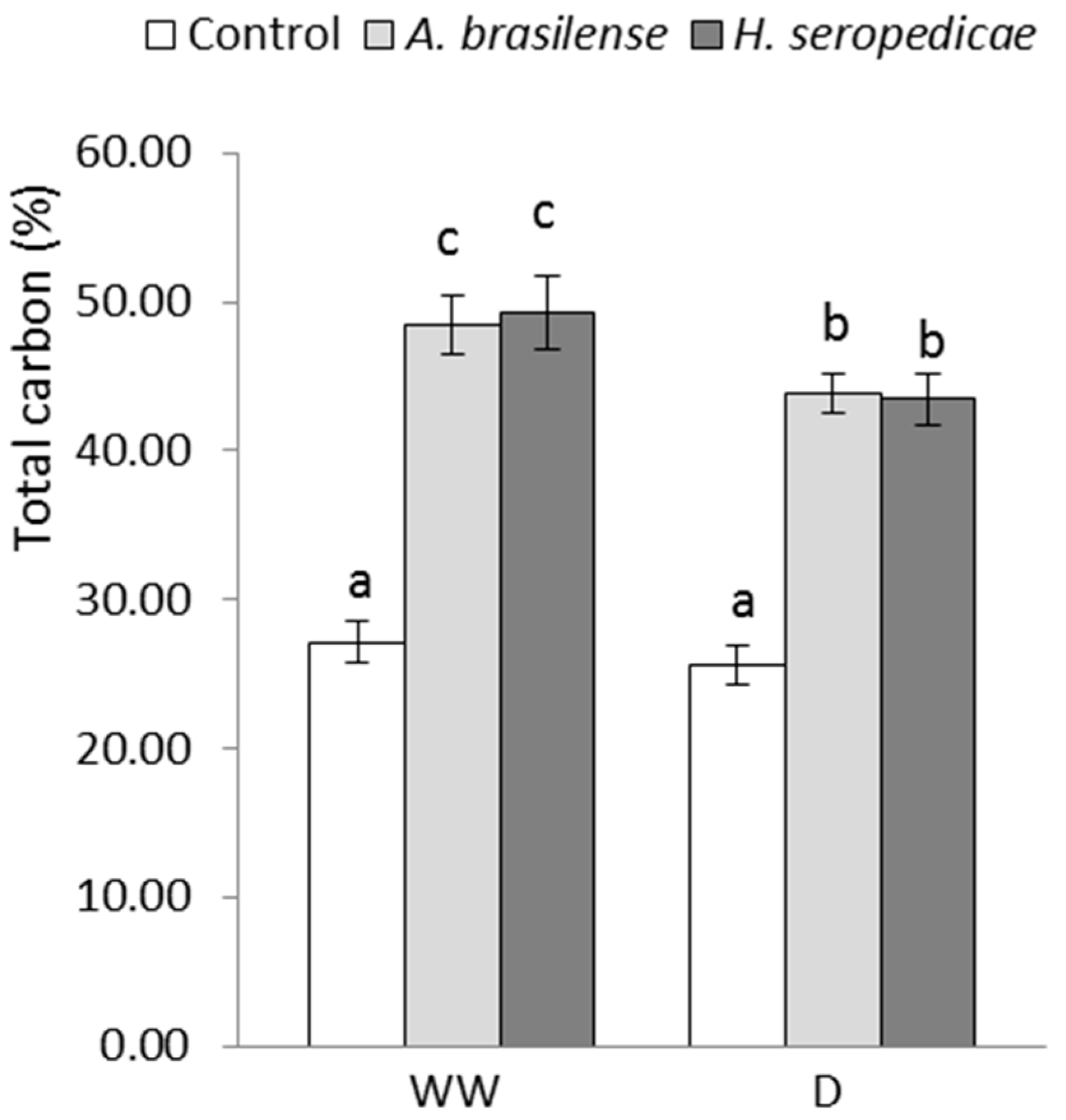
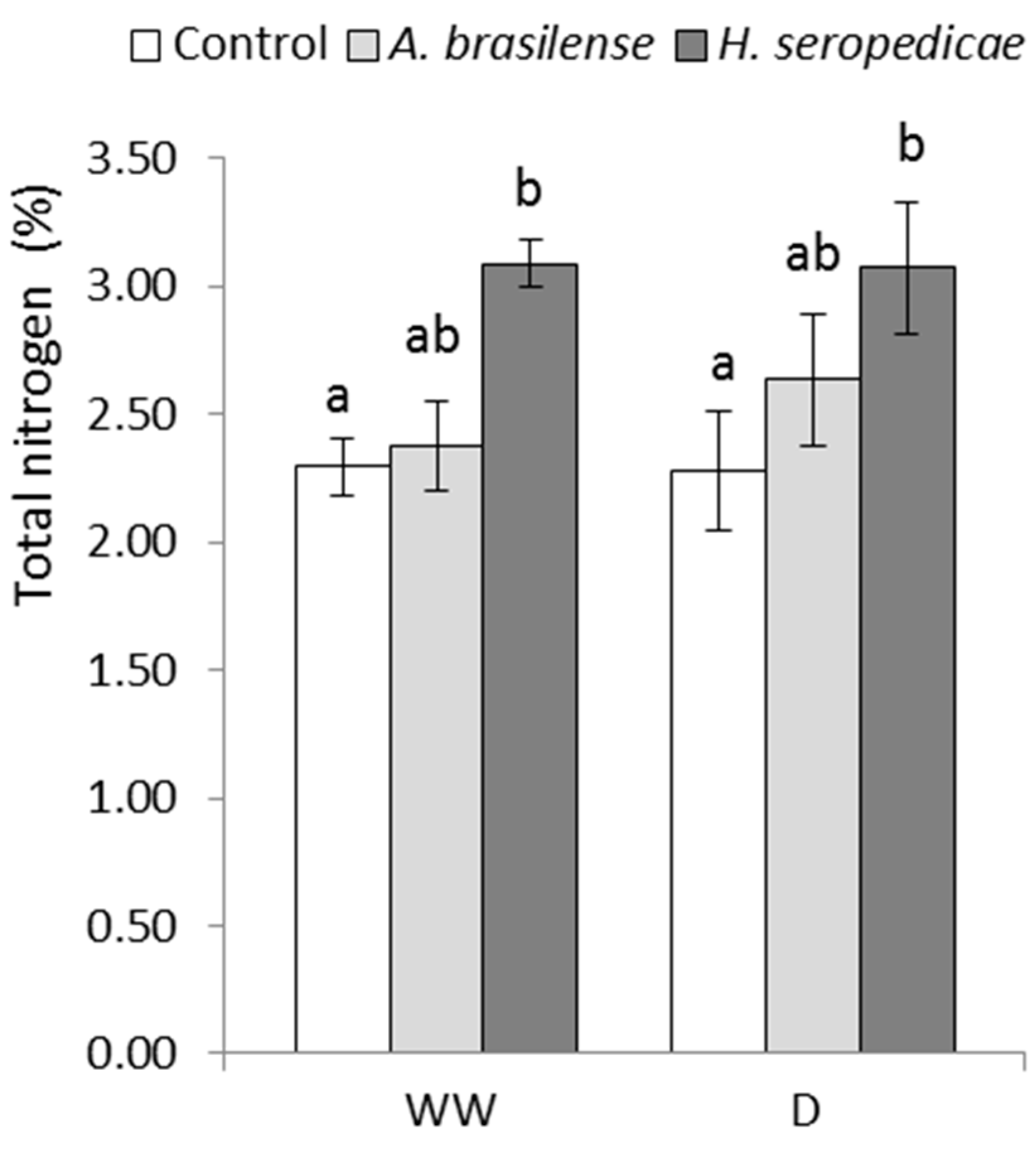
2.4.3. Total Carbon (TC) and Total Nitrogen (TN) Content
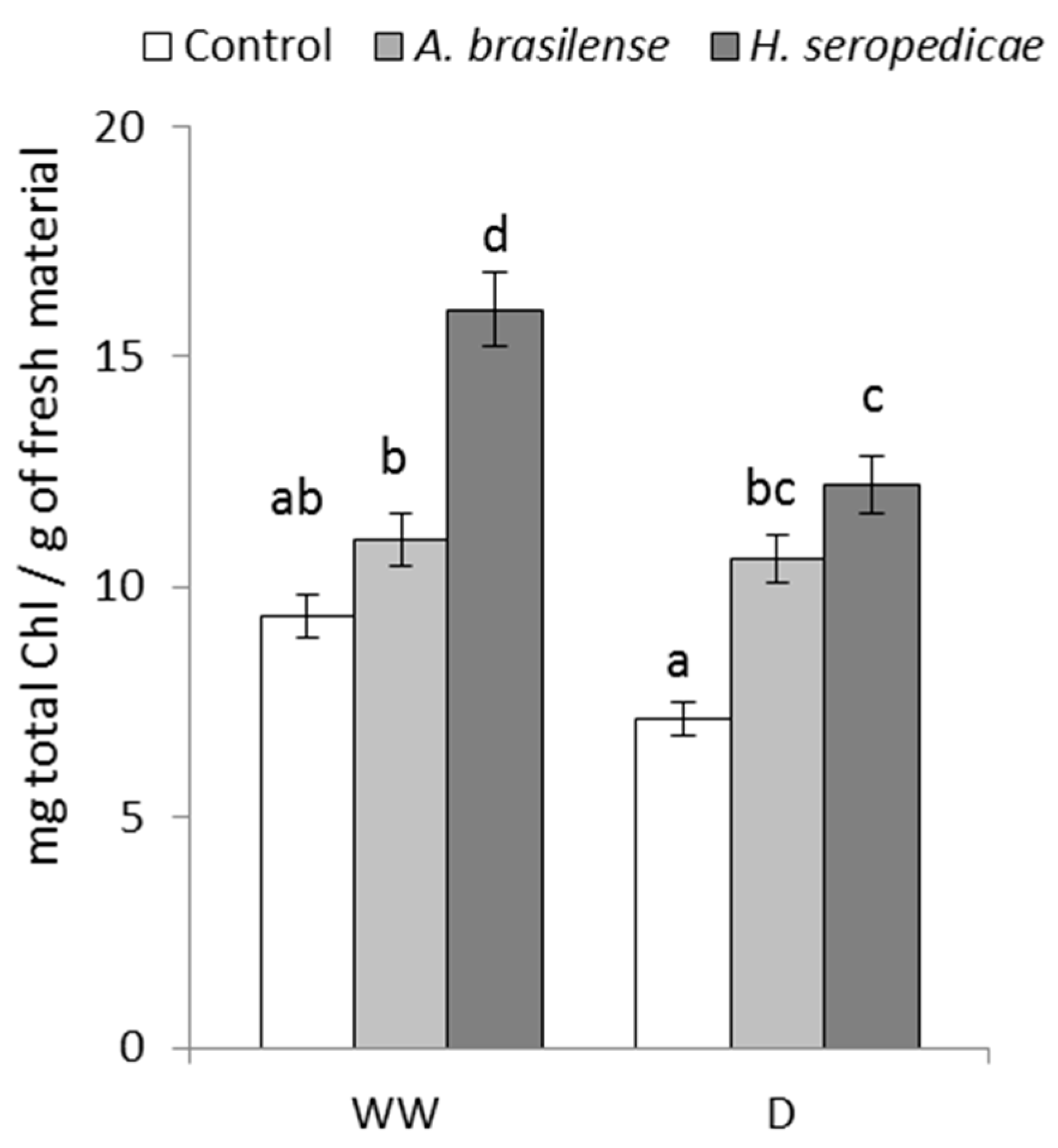
2.4.4. Total Chlorophyll (TChl) Content
2.4.5. ABA and Ethylene Content
2.4.6. Relative Water Content (RWC)
2.4.7. Malondialdehyde (MDA) Content
2.4.8. Proline Content
2.4.9. Differential Expression of the ZmVP14 Gene
3. Results
3.1. Symbiotic Development
3.2. TB Production
3.3. TC and TN Content
3.4. TChl Content
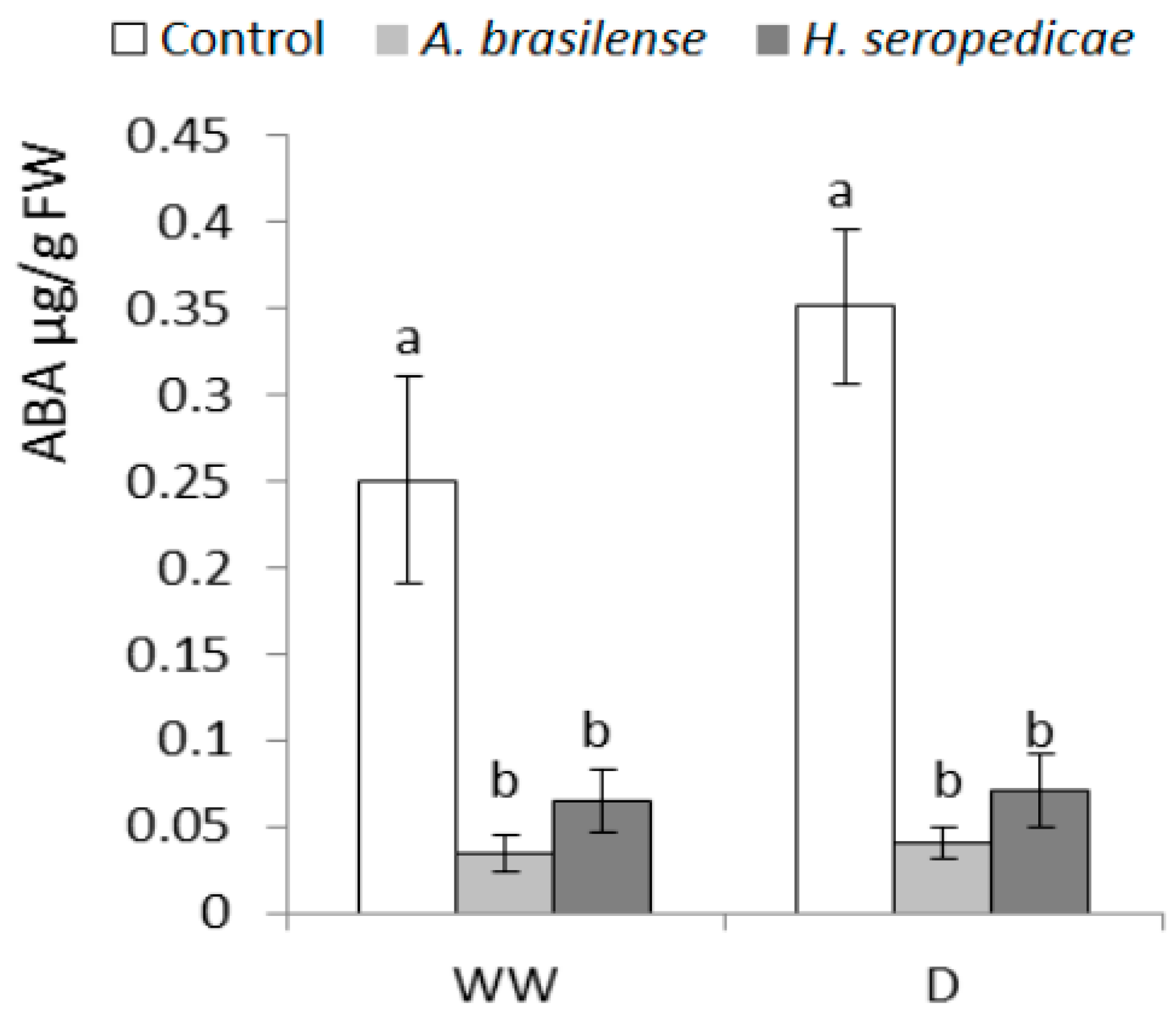
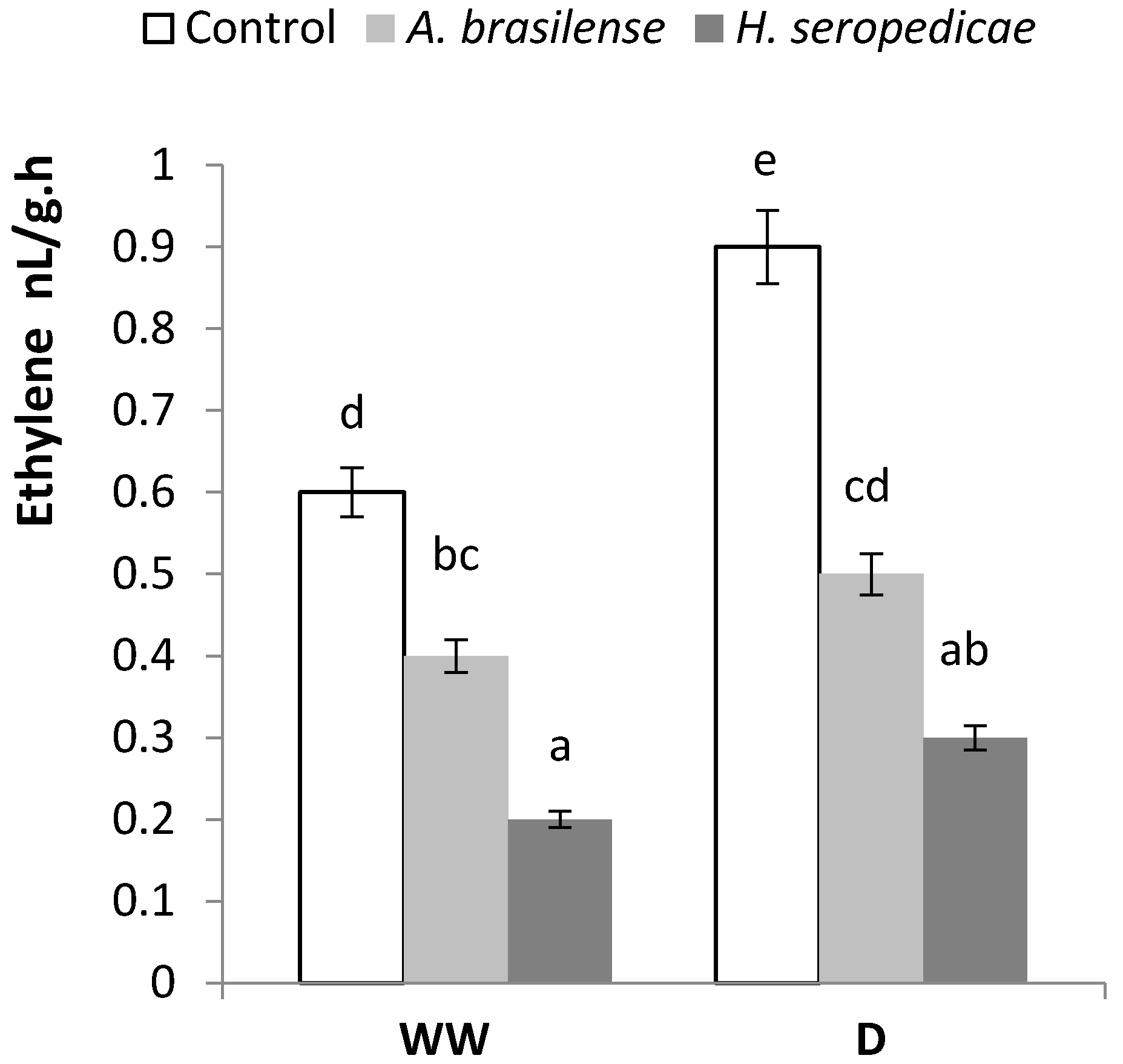
3.5. ABA and Ethylene Content
3.6. RWC
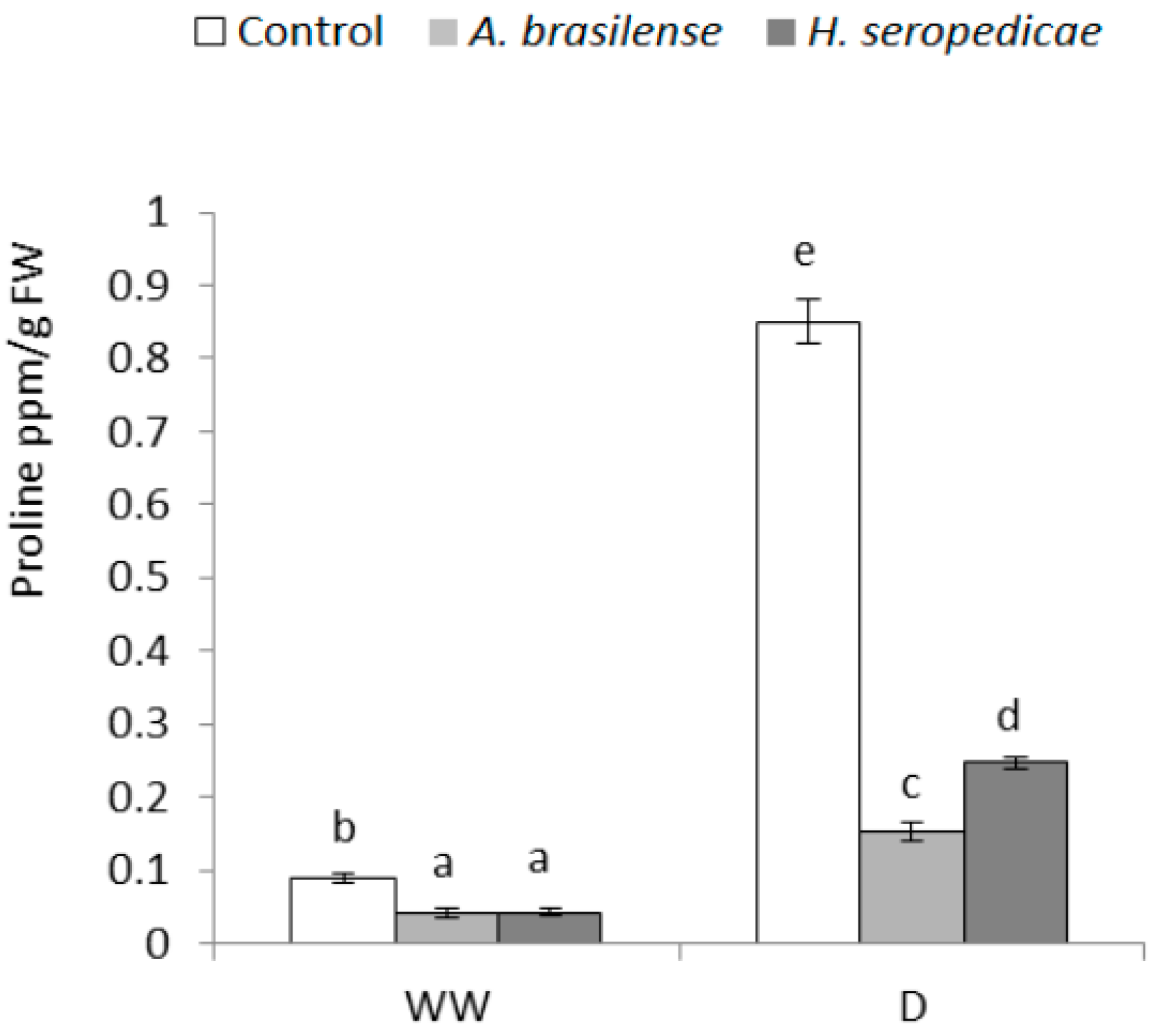
3.7. Proline Content
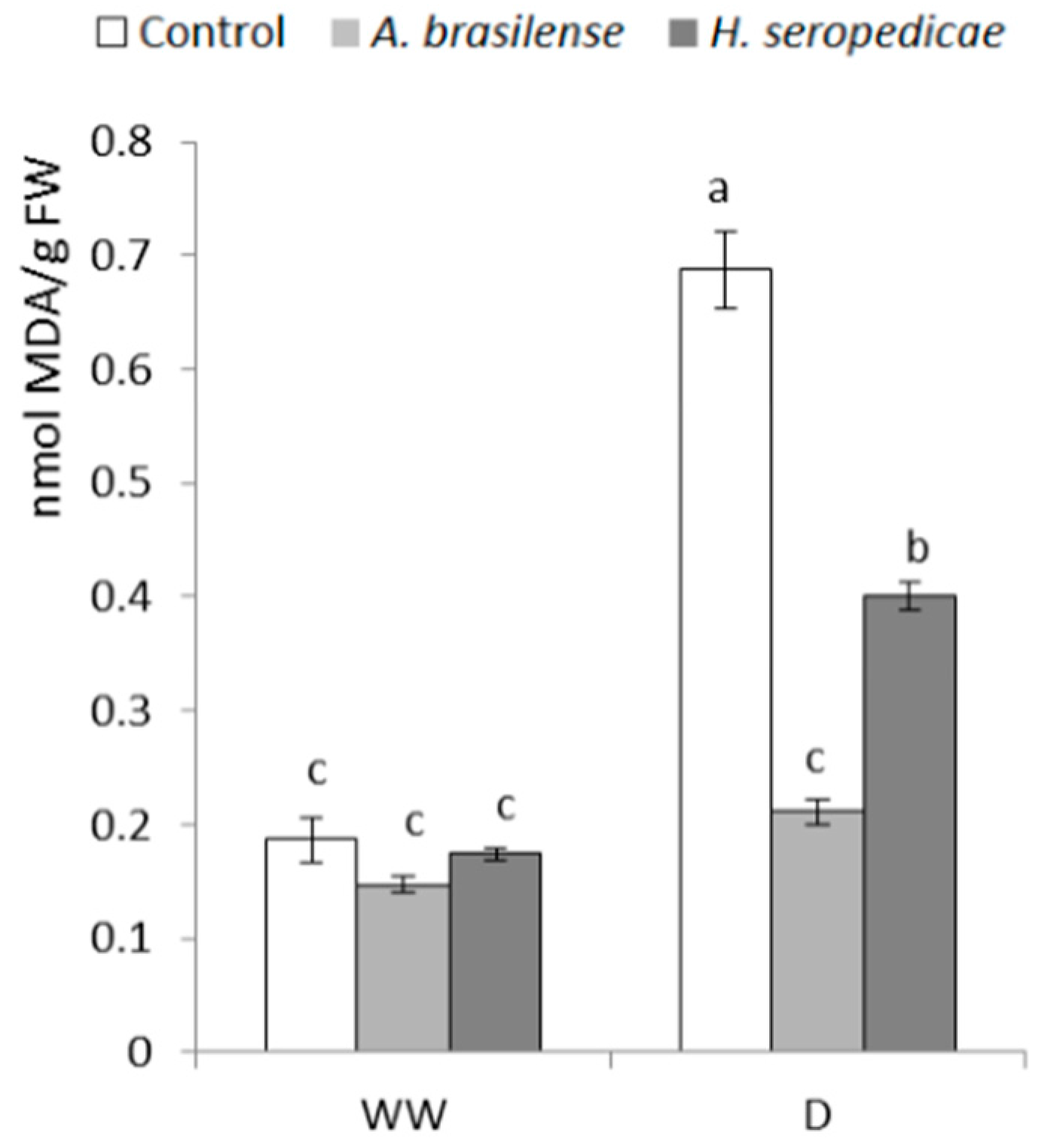
3.8. MDA Content
3.9. Expression of the ZmVP14 Gene
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- FAOSTAT. 2013. Available online: http://faostat3.fao.org/browse/Q/QC/S (accessed on 1 October 2016).
- Daryanto, S.; Wang, L.; Jacinthe, P.A. Global Synthesis of Drought Effects on Maize and Wheat Production. PLoS ONE 2016, 11, e0156362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heiniger, R.W. Irrigation and Drought Management; North Carolina State University: Raleigh, NC, USA, 2000. [Google Scholar]
- Edmeades, G.O. Drought tolerance in maize: An emerging reality. A feature. In Global Status of Commercialized Biotech/GM Crops; James, C., Ed.; ISAAA Brief No. 39; ISAAA: Ithaca, NY, USA, 2008; p. 275. [Google Scholar]
- Lata, C.; Muthamilarasan, M.; Prasad, M. Drought stress responses and signal transduction in plants. In Elucidation of Abiotic Stress Signaling in Plants; Pandey, G.K., Ed.; Springer: New York, NY, USA, 2015; pp. 195–225. [Google Scholar]
- Lata, C.; Jha, S.; Dixit, V.; Sreenivasulu, N.; Prasad, M. Differential antioxidative responses to dehydration-induced oxidative stress in core set of foxtail millet cultivars (Setaria italica L.). Protoplasma 2011, 248, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Silveira, J.A.G.; Costa, R.C.L.; Oliveira, J.T.A. Drought-induced effects and recovery of nitrate assimilation and nodule activity in cowpea plants inoculated with Bradyrhizobium spp. under moderate nitrate level. Braz. J. Microbiol. 2001, 32, 187–194. [Google Scholar] [CrossRef]
- Riera, M.; Valon, C.; Fenzi, F.; Giraudat, J.; Leung, J. The genetics of adaptive responses to drought stress: Abscisic acid-dependent and abscisic acid-independent signalling components. Physiol. Plant. 2005, 123, 111–119. [Google Scholar] [CrossRef]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Molecular responses to dehydration and low temperature: Differences and cross-talk between two stress signaling pathways. Curr. Opin. Plant Biol. 2000, 3, 217–223. [Google Scholar] [CrossRef]
- Finkelstein, R.R.; Gampala, S.S.; Rock, C.D. Abscisic acid signaling in seeds and seedlings. Plant Cell 2002, 14, S15–S45. [Google Scholar] [PubMed]
- Xiong, L.; Schumaker, K.; Zhu, J.K. Cell signaling during cold, drought and salt stress. Plant Cell 2002, 14, S165–S183. [Google Scholar] [CrossRef] [PubMed]
- Taiz, L.; Zeiger, E. Responses and Adaptations to Abiotic Stress. In Plant Physiology, 5th ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 2012; pp. 755–778. [Google Scholar]
- Zeevaart, J.A.D. Abscisic acid metabolism and its regulation. In Biochemistry and Molecular Biology of Plant Hormones; Hooykaas, P.J., Hall, M.A., Libbenga, R., Eds.; Elsevier: Amsterdam, The Netherlands, 1999; pp. 189–207. [Google Scholar]
- Xiong, L.; Zhu, J.K. Regulation of Abscisic acid biosynthesis. Plant Physiol. 2003, 133, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.J.; Jackson, A.C.; Symonds, R.C.; Mulholland, B.J.; Dadswell, A.R.; Blake, P.S.; Burbidge, A.; Taylor, I.B. Ectopic expression of a tomato 9-cis-epoxycarotenoid dioxygenase gene causes over-production of abscisic acid. Plant J. 2000, 23, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.C.; Schwartz, S.H.; Zeevaart, J.A.D.; McCarty, D.R. Genetic control of abscisic acid biosynthesis in maize. Proc. Natl. Acad. Sci. USA 1997, 94, 12235–12240. [Google Scholar] [CrossRef] [PubMed]
- Hardoim, P.R.; van Overbeek, L.S.; van Elsas, J.D. Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol. 2008, 16, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. Modulation of plant ethylene levels by the bacterial enzyme ACC deaminase. FEMS Microbiol. Lett. 2005, 251, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Inzé, D.; van Montagu, M. Oxidative stress in plants. Curr. Opin. Biotechnol. 1995, 6, 153–158. [Google Scholar] [CrossRef]
- Storz, G.; Imalay, J.A. Oxidative stress. Curr. Opin. Microbiol. 1999, 2, 188–194. [Google Scholar] [CrossRef]
- Chugh, V.; Kaur, N.; Gupta, A.K. Evaluation of oxidative stress tolerance in maize (Zea mays L.) seedlings in response to drought. Indian J. Biochem. Biophys. 2011, 48, 47–53. [Google Scholar] [PubMed]
- Chimenti, C.A.; Marcantonio, M.; Hall, A.J. Divergent selection for osmotic adjustment results in improved drought tolerance in maize (Zea mays L.) in both early growth and flowering phases. Field Crop. Res. 2005, 95, 305–315. [Google Scholar] [CrossRef]
- Parra-Quezada, R.A.; Rodríguez-Ontiveros, J.L.; González-Hernández, V.A. Transpiración, potencial hídrico y prolina en zarzamora bajo déficit hídrico. Terra Latinoam. 1999, 17, 125–130. [Google Scholar]
- Kloepper, J.W.; Schroth, M.N. Plant growth promoting rhizobacteria on radishes. In Proceedings of the 4th International Conference on Plant Pathogenic Bacteria, Angers, France, August 1978; I.N.R.A.: Angers, France; Volume 2, pp. 879–882. [Google Scholar]
- Glick, B.R. The enhancement of plant growth by free-living bacteria. Can. J. Microbiol. 1995, 41, 109–117. [Google Scholar] [CrossRef]
- Bashan, Y.; Holguin, G. Proposal for the division plant growth promoting rhizobacteria into two classifications: Biocontrol-PGPB (plant growth-promoting bacteria) and PGPB. Soil Biol. Biochem. 1998, 30, 1225–1228. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Rodriguez-Ubana, R.; Zehnder, G.W.; Murphy, J.F.; Sikora, E.; Fernández, C. Plant root-bacterial interactions in biological control of soilborne diseases and potencial extensión to systemic and foliar diseases. Australas. Plant Pathol. 1999, 28, 21–26. [Google Scholar] [CrossRef]
- Bashan, J.; Holguin, G. Root-to-root travel of the benefitial bacterium Azospirillum brasilense. Appl. Environ. Microbiol. 1997, 60, 2120–2131. [Google Scholar]
- Cassán, F.; Bottini, R.; Schneider, G.; Piccoli, P. Azospirillum brasilense and Azospirillum lipoferum hydrolize conjugates of GA20 and metabolize the resultant aglycones to GA1 in seedlings of rice dwarf mutants. Plant Physiol. 2001, 125, 2053–2058. [Google Scholar] [CrossRef] [PubMed]
- Bottini, R.; Cassán, F.; Picolli, P. Gibberellin production by bacteria and its involvement in plant growth promotion and yield increase. Appl. Microbiol. Biotechnol. 2004, 65, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Ribaudo, C.M.; Rondanini, D.P.; Trinchero, G.D.; Curá, J.A. Effect of Herbaspirillum seropedicae inoculation on maize nitrogen metabolism. Maydica 2006, 51, 481–485. [Google Scholar]
- De Salamone, I.G.; Döbereiner, J. Maize genotype effects on the response to Azospirillum inoculation. Biol. Fertil. Soils 1996, 21, 193–196. [Google Scholar] [CrossRef]
- Ribaudo, C.M.; Paccusse, N.A.; Rondanini, D.P.; Curá, J.A.; Fraschina, A.A. Maize-Azospirillum association: Effects on the dry matter yield and nitrate reductase activity. Agric. Trop. Subtrop. 1998, 31, 61–70. [Google Scholar]
- Ribaudo, C.M.; Krumpholz, E.M.; Cassán, F.D.; Bottini, R.; Cantore, M.L.; Curá, J.A. Azospirillum sp. promotes growth of tomato plants by modulating the levels of root-produced ethylene. J. Plant Growth Regul. 2006, 24, 175–185. [Google Scholar] [CrossRef]
- Nico, M.; Ribaudo, C.M.; Gori, J.I.; Cantore, M.L.; Curá, J.A. Uptake of phosphate and promotion of vegetative growth in glucose-exuding rice plants (Oryza sativa) inoculated with plant growth-promoting bacteria. Appl. Soil Ecol. 2012, 65, 190–195. [Google Scholar] [CrossRef]
- Nielsen, T.H.; Sorensen, D.; Tobiasen, C.; Andersen, J.B.; Christeophersen, C.; Givskov, M.; Sorensen, J. Antibiotic and biosurfactant properties of cyclic lipopeptides produced by fluorescent Pseudomonas spp. from the sugar beet rhizosphere. Appl. Environ. Microbiol. 2002, 63, 3416–3423. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Vlami, M.; de Souza, J.T. Antibiotic production by bacteria biocontrol agents. Antonie Van Leeuwenhoek 2002, 81, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Whipps, J.M. Microbial interactions and biocontrol in the rhizosphere. J. Exp. Bot. 2001, 52, 487–511. [Google Scholar] [CrossRef] [PubMed]
- Frankowski, J.; Lorito, M.; Scala, F.; Schmidt, R.; Berg, G.; Bahl, H. Purification and properties of two chitinolytic enzymes of Serratia plymuthica HRO-C48. Arch. Microbiol. 2001, 176, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.H.; Zhang, X.F.; Xu, J.L.; Zhang, L.H. Insecticidal Bacillus thuringiensis silences Erwinia carotovora virulence by a new form of microbial antagonism, signal interference. Appl. Environ. Microbiol. 2004, 70, 954–960. [Google Scholar] [CrossRef] [PubMed]
- Kloepper, J.W. Plant growth-promoting rhizobacteria (other systems). In Azospirillum/Plant Associations; Okon, Y., Ed.; CRC Press: Boca Raton, FL, USA, 1995; pp. 137–166. [Google Scholar]
- Bashan, Y.; Holguin, G. Azospirillum—Plant relationships: Environmental and physiological advances (1990–1996). Can. J. Microbiol. 1997, 43, 103–121. [Google Scholar] [CrossRef]
- Holguin, G.; Patten, C.L.; Glick, B.R. Genetics and molecular biology of Azospirillum. Biol. Fertil. Soils 1999, 29, 10–23. [Google Scholar] [CrossRef]
- Steenhoudt, O.; Vanderleyden, J. Azospirillum, a free-living nitrogen fixing bacterium closely associated grasses: Genetic, biochemical and ecological aspects. FEMS Microbiol. Rev. 2000, 24, 487–506. [Google Scholar] [CrossRef] [PubMed]
- Okon, Y.; Labandera-González, C.A. Agronomic applications of Azospirillum: An evaluation of 20 years worldwide field inoculation. Soil Biol. Biochem. 1994, 26, 1591–1601. [Google Scholar] [CrossRef]
- Boddey, R.M.; de Oliveira, O.C.; Urquiaga, S.; Reis, V.M.; Olivares, F.L.; Baldani, V.L.D.; Döbereiner, J. Biological nitrogen fixation associated with sugar cane and rice: Contributions and prospects for improvement. Plant Soil 1995, 174, 195–209. [Google Scholar] [CrossRef]
- James, E.K. Nitrogen fixation in endophytic and associative symbiosis. Field Crop Res. 2000, 65, 197–209. [Google Scholar] [CrossRef]
- Elbeltagy, A.; Nishioka, K.; Sato, T. Endophytic colonization and in plant nitrogen fixation by a Herbaspirillum sp. isolated from wild rice species. Appl. Environ. Microbiol. 2001, 67, 5285–5293. [Google Scholar] [CrossRef] [PubMed]
- James, E.K.; Gyaneshwar, P.; Mathan, N.; Barraquio, W.L.; Reddy, P.M.; Iannetta, P.P.M.; Olivares, F.L.; Ladha, J.K. Infection and colonization of rice seedlings by the plant growth promotion bacterium Herbaspirillum seropedicae Z67. Mol. Plant Microbe Interact. 2002, 15, 894–906. [Google Scholar] [CrossRef] [PubMed]
- Casanovas, E.M.; Barassi, C.A.; Sueldo, R.J. Azospirillum inoculation mitigates water stress effects in maize seedlings. Cereal Res. Commun. 2002, 30, 343–350. [Google Scholar]
- Marulanda, A.; Barea, J.M.; Azcón, R. Stimulation of plant growth and drought tolerance by native microorganisms (AM fungi and bacteria) from dry environments mechanisms related to bacterial effectiveness. J. Plant Growth Regul. 2009, 28, 115–124. [Google Scholar] [CrossRef]
- Yang, J.; Kloepper, J.W.; Ryu, C.M. Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 2009, 14, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Ruíz-Sánchez, M.; Armada, E.; Muñoz, Y.; García de Salamone, I.E.; Aroca, R.; Ruíz-Lozano, J.M.; Azcón, R. Azospirillum and arbuscular mycorrhizal colonization enhance rice growth and physiological traits under well-watered and drought conditions. J. Plant Physiol. 2011, 168, 1031–1037. [Google Scholar] [CrossRef] [PubMed]
- Nautiyal, C.S.; Srivastava, S.; Chauhan, P.S.; Seem, K.; Mishra, A.; Sopory, S.K. Plant growth-promoting bacteria Bacillus amyloliquefaciens NBRISN13 modulates gene expression profile of leaf and rhizosphere community in rice during salt stress. Plant Physiol. Biochem. 2013, 66, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Naveed, M.; Mitter, B.; Reichenauer, T.G.; Wieczorek, K.; Sessitsch, A. Increased drought stress resilience of maize through endophytic colonization by Burkholderia phytofirmans PsJN and Enterobacter sp. FD17. Environ. Exp. Bot. 2014, 97, 30–39. [Google Scholar] [CrossRef]
- Cohen, A.C.; Bottini, R.; Pontin, M.; Berli, F.J.; Moreno, D.; Boccanlandro, H.; Travaglia, C.N.; Piccoli, P.N. Azospirillum brasilense ameliorates the response of Arabidopsis thaliana to drought mainly via enhancement of ABA levels. Physiol. Plant. 2015, 153, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Lata, C.; Chauhan, P.S.; Nautiyal, C.S. Pseudomonas putida attunes morphophysiological, biochemical and molecular responses in Cicer arietinum L. during drought stress and recovery. Plant Physiol. Biochem. 2016, 99, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol. Res. 2016, 184, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Döbereiner, J.; Marriel, I.E.; Nery, M. Ecological distribution of Spirillum lipoferum Beijerinck. Can. J. Microbiol. 1976, 22, 1464–1473. [Google Scholar] [CrossRef] [PubMed]
- FCA-UNC. Facultad de Ciencias Agropecuarias. Universidad Nacional de Córdoba. Available online: http://www.infostat.com.ar/index.php (accessed on 1 October 2016).
- Baldani, V.L.D.; Baldani, J.I.; Olivares, F.L.; Döbereiner, J. Identification and ecology of Herbaspirillum seropedicae and closely related Pseudomonas rubrisubalbicans. Symbiosis 1992, 13, 65–73. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plantarum 1962, 18, 100–127. [Google Scholar] [CrossRef]
- Rodríguez-Cáceres, E. Improved medium for isolation of Azospirillum spp. Appl. Environ. Microbiol. 1982, 44, 990–991. [Google Scholar]
- Walkley, A.; Black, I. An examination of the Degtjareff method and a proposed modification of the chromic matter and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 34, 29–38. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists. Official Methods of Analysis, 12th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1975.
- Inskeep, W.P.; Bloom, P.R. Extinction coefficients of chlorophyll a and b in N,N-Dimethylformamide and 80% Acetone. Plant Physiol. 1985, 77, 483–485. [Google Scholar] [CrossRef] [PubMed]
- Kelen, M.; Cubuk Demiralay, E.; Sen, S.; Ozkan, G. Separation of Abscisic Acid, Indole-3-Acetic Acid, Gibberellic Acid in 99 R (Vitis berlandieri x Vitis rupestris) and Rose Oil (Rosa damascena Mill.) by Reversed Phase Liquid Chromatography. Turk. J. Chem. 2004, 28, 603–610. [Google Scholar]
- Iriti, M.; Picchi, V.; Rossoni, M.; Gomarasca, S.; Ludwig, N.; Gargano, M.; Faoro, F. Chitosan antitranspirant activity is due to abscisic acid-dependent stomatal closure. Environ. Exp. Bot. 2009, 66, 493–500. [Google Scholar] [CrossRef]
- Teulat, B.; Zoumarou-Wallis, N.; Rotter, B.; Ben Salem, M.; Bahri, H.; This, D. QTL for relative water content in field-grown barley and their stability across Mediterranean environments. Theor. Appl. Genet. 2003, 108, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Troll, W.; Lindsley, J. A photometric method for the determination of proline. J. Biol. Chem. 1955, 215, 655–660. [Google Scholar] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Salinas-Moreno, Y.; González-Henández, V.A. Potencial hídrico y ácido abscísico en hoja, jilote y espiga de maíz sometido a sequía. Agricultura Técnica en México 2003, 29, 135–143. [Google Scholar]
- Gagné-Bourque, F.; Mayer, B.F.; Charron1, J.B.; Vali, H.; Bertrand, A.; Jabaji, S. Accelerated Growth Rate and Increased Drought Stress Resilience of the Model Grass Brachypodium distachyon Colonized by Bacillus subtilis B26. PLoS ONE 2015, 10, e0130456. [Google Scholar] [CrossRef] [PubMed]
- Moussa, H.R.; Abdel-Aziz, S.M. Comparative response of drought tolerant and drought sensitive maize genotypes to water stress. Aust. J. Crop. Sci. 2008, 1, 31–36. [Google Scholar]
- Paleg, L.G.; Stewart, G.R.; Bradbeer, J.W. Proline and Glycine Betaine Influence Protein Solvation. Plant Physiol. 1983, 75, 974–978. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2005, 59, 206–216. [Google Scholar] [CrossRef]
- Bogges, S.F.; Aspinnal, D.; Paleg, L.G. Inhibition of Proline Oxidation by Water Stress. Plant Physiol. 1977, 59, 930–932. [Google Scholar] [CrossRef]
- Zhou, C.; Ma, Z.; Zhu, L.; Xiao, X.; Xie, Y.; Zhu, J.; Wang, J. Rhizobacterial Strain Bacillus megaterium BOFC15 Induces Cellular Polyamine Changes that Improve Plant Growth and Drought Resistance. Int. J. Mol. Sci. 2016, 17, 976. [Google Scholar] [CrossRef] [PubMed]
- Chernys, J.T.; Zeevaart, J.A.D. Characterization of the 9-cis-Epoxycarotenoid Dioxygenase Gene Family and the Regulation of Abscisic Acid Biosynthesis in Avocado. Plant Physiol. 2000, 124, 343–353. [Google Scholar] [CrossRef] [PubMed]
- El-Komy, H.M.; Hamdia, M.A.; El-Baki, G.K.A. Nitrate reductase in wheat plants grown under water stress and inoculated with Azospirillum spp. Biol. Plant. 2003, 46, 281–287. [Google Scholar] [CrossRef]
- Creus, C.M.; Graziano, M.; Casanovas, E.M.; Pereyra, M.A.; Simontacchi, M.; Puntarulo, S.; Barassi, C.A.; Lamattina, L. Nitric oxide is involved in the Azospirillum brasilense-induced lateral root formation in tomato. Planta 2005, 221, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Cassán, F.D.; Maiale, S.; Masciarelli, O.; Vidal, A.; Luna, V.; Ruiz, O. Cadaverine production by Azospirillum brasilense and its possible role in plant growth promotion and osmotic stress mitigation. Eur. J. Soil Biol. 2009, 45, 12–19. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | RWC (%) | |
|---|---|---|
| WW | D | |
| Control | 52.71 ± 2.5 c | 45.45 ± 1.17 a |
| A. brasilense | 58.81 ± 2.7 e | 47.86 ± 1.37 b |
| H. seropedicae | 53.93 ± 3.16 d | 48.1 ± 1.95 b |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Curá, J.A.; Franz, D.R.; Filosofía, J.E.; Balestrasse, K.B.; Burgueño, L.E. Inoculation with Azospirillum sp. and Herbaspirillum sp. Bacteria Increases the Tolerance of Maize to Drought Stress. Microorganisms 2017, 5, 41. https://doi.org/10.3390/microorganisms5030041
Curá JA, Franz DR, Filosofía JE, Balestrasse KB, Burgueño LE. Inoculation with Azospirillum sp. and Herbaspirillum sp. Bacteria Increases the Tolerance of Maize to Drought Stress. Microorganisms. 2017; 5(3):41. https://doi.org/10.3390/microorganisms5030041
Chicago/Turabian StyleCurá, José Alfredo, Diego Reinaldo Franz, Julián Ezequiel Filosofía, Karina Beatríz Balestrasse, and Lautaro Exequiel Burgueño. 2017. "Inoculation with Azospirillum sp. and Herbaspirillum sp. Bacteria Increases the Tolerance of Maize to Drought Stress" Microorganisms 5, no. 3: 41. https://doi.org/10.3390/microorganisms5030041
APA StyleCurá, J. A., Franz, D. R., Filosofía, J. E., Balestrasse, K. B., & Burgueño, L. E. (2017). Inoculation with Azospirillum sp. and Herbaspirillum sp. Bacteria Increases the Tolerance of Maize to Drought Stress. Microorganisms, 5(3), 41. https://doi.org/10.3390/microorganisms5030041