Combating Fusarium Infection Using Bacillus-Based Antimicrobials


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. F. oxysporum and Vascular Wilt
3. F. graminearum, Killer of Cereals
4. Major Fusarium Disease Determinants: Mycotoxins
5. Management of Fusarium Wilt
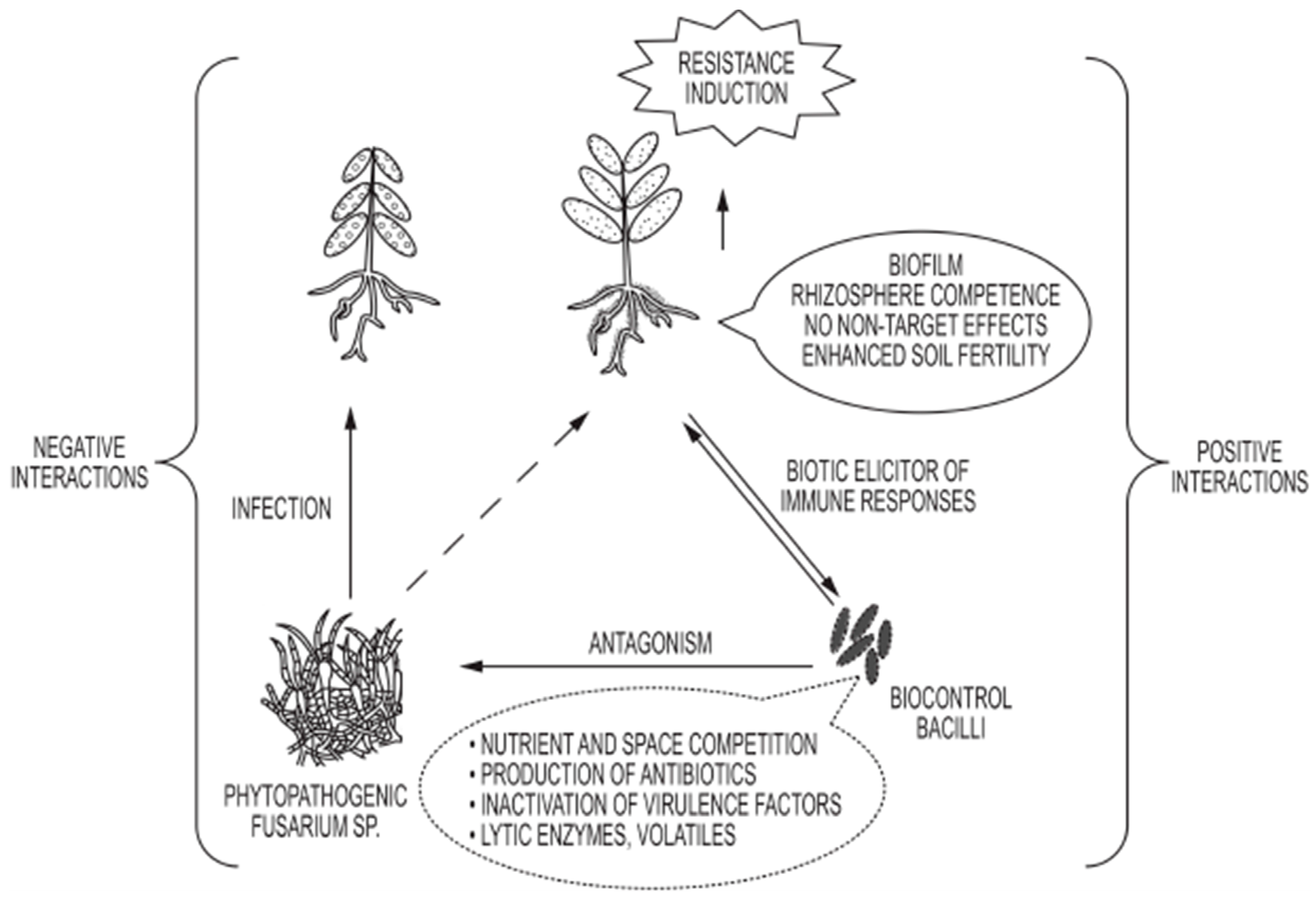
6. Biocontrol Attributes of Various Bacillus Species
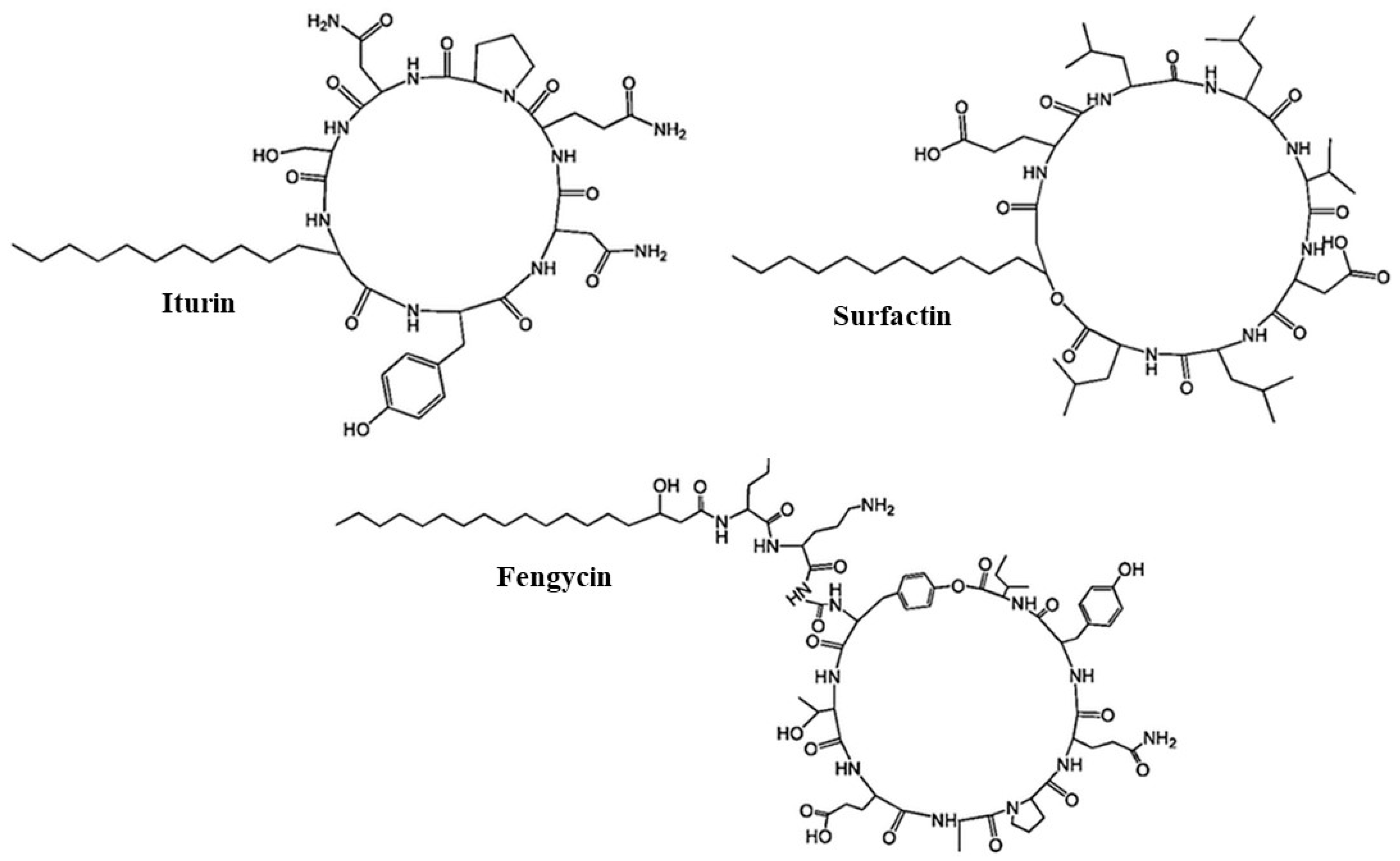
6.1. Bacillus Peptide Antibiotics
6.2. Non-Ribosomal Biosynthesized Peptides
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cao, Y.; Zhang, Z.; Ling, N.; Yuan, Y.; Zheng, X.; Shen, B.; Shen, Q. Bacillus subtilis SQR 9 can control Fusarium wilt in cucumber by colonizing plant roots. Biol. Fertil. Soils 2011, 47, 495–506. [Google Scholar] [CrossRef]
- Mazzola, M.; Freilich, S. Prospects for biological soilborne disease control: Application of indigenous versus synthetic microbiomes. Phytopathology 2017, 107, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Haas, D.; Geneviève, D. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Veliz, E.A.; Martinez-Hidalgo, P.; Hirsch, A.M. Chitinase producing bacteria and their role in biocontrol. AIMS Microbiol. 2017, 3, 689–705. [Google Scholar] [CrossRef]
- Seiber, J.N.; Coats, J.; Duke, S.O.; Gross, A.D. Biopesticides: State of the art and future opportunities. J. Agric. Food Chem. 2014, 62, 11613–11619. [Google Scholar] [CrossRef] [PubMed]
- Bardin, M.; Ajouz, S.; Comby, M.; Lopez-Ferber, M.; Graillot, B.; Siegwart, M.; Nicot, P.C. Is the efficacy of biological control against plant diseases likely to be more durable than that of chemical pesticides? Front. Plant Sci. 2015, 6, 566. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.H.; Matthee, F.N.; Thomas, A.C. Biological control of Eutypa lata on grapevine by an antagonistic strain of Bacillus subtilis. Phytopathology 1991, 81, 283–287. [Google Scholar] [CrossRef]
- Law, J.W.-F.; Ser, H.-L.; Khan, T.M.; Chuah, L.-H.; Pusparajah, P.; Chan, K.-G.; Goh, B.-H.; Lee, L.-H. The potential of Streptomyces as biocontrol agents against the rice blast fungus, Magnaporthe oryzae (Pyricularia oryzae). Front. Microbiol. 2017, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, W.L.; Munakata, N.; Horneck, G.; Melosh, H.J.; Setlow, P. Resistance of Bacillus endospores to extreme terrestrial and extraterrestrial environments. Microbiol. Mol. Biol. Rev. 2000, 64, 548–572. [Google Scholar] [CrossRef] [PubMed]
- Angus, A.A.; Agapakis, C.M.; Fong, S.; Yerrapragada, S.; Estrada-de los Santos, P.; Yang, P.; Hirsch, A.M. Plant-associated symbiotic Burkholderia species lack hallmark strategies required in mammalian pathogenesis. PLoS ONE 2014, 9, e83779. [Google Scholar] [CrossRef] [PubMed]
- Donato, V.; Ayala, F.R.; Cogliati, S.; Bauman, C.; Costa, J.B.; Lenini, C.; Grau, R. Bacillus subtilis biofilm extends Caenorhabditis elegans longevity through downregulation of the insulin-like signalling pathway. Nat. Commun. 2017, 8, 14332. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Arrabit, M.; Hirsch, A.M. Interactions of Plant Beneficial Bacillus spp. and Caenorhabditis elegans—A Tool to Evaluate Pathogenic Potential of Bioinoculants. 2017; manuscript in preparation. [Google Scholar]
- Allard-Massicotte, R.; Tessier, L.; Lecuyer, F.; Lakshmanan, V.; Lucier, J.F.; Garneau, D.; Claudwell, L.; Vlamakis, H.; Bais, H.P.; Beauregard, P.B. Bacillus subtilis early colonization of Arabidopsis thaliana roots involve multiple chemotaxis receptors. mBio 2016, 7, e01664-16. [Google Scholar] [CrossRef] [PubMed]
- Beneduzi, A.; Ambrosini, A.; Passaglia, L.M.P. Plant growth-promoting rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genet. Mol. Biol. 2012, 35, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Saraf, M.; Pandya, U.; Thakkar, A. Role of allelochemicals in plant growth promoting rhizobacteria for biocontrol of phytopathogens. Microbiol. Res. 2014, 169, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Shlevin, E.; Mahrer, Y.; Katan, J. Effect of moisture on thermal inactivation of soilborne pathogens under structural solarization. Phytopathology 2004, 94, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Duan, C.; Qin, Z.; Yang, Z.; Li, W.; Sun, S.; Zhu, Z.; Wang, X. Identification of pathogenic Fusarium spp. causing maize ear rot and potential mycotoxin production in China. Toxins 2016, 8, 186. [Google Scholar] [CrossRef] [PubMed]
- McGovern, R.J. Management of tomato diseases caused by Fusarium oxysporum. Crop Protect. 2015, 73, 78–92. [Google Scholar] [CrossRef]
- Gu, Q.; Yang, Y.; Yuan, Q.; Shi, G.; Wu, L.; Lou, Z.; Huo, R.; Wu, H.; Borriss, R.; Gao, X. Bacillomycin D produced by Bacillus amyloliquefaciens is involved in the antagonistic interaction with the plant-pathogenic fungus Fusarium graminearum. Appl. Environ. Microbiol. 2017, 83, e01075-17. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, G.M.; Armstrong, J.K. Formae speciales and races of Fusarium oxysporum causing wilt diseases. In Fusarium: Diseases, Biology and Taxonomy; Nelson, P.E., Toussoun, T.A., Eds.; Pennsylvania State University Press: University Park, PA, USA, 1981; pp. 391–399. [Google Scholar]
- Fourie, G.; Steenkamp, E.T.; Ploetz, R.C.; Gordon, T.R.; Viljoen, A. Current status of the taxonomic position of Fusarium oxysporum formae specialis cubense within the Fusarium oxysporum complex. Infect. Genet. Evol. 2011, 11, 533–542. [Google Scholar] [CrossRef] [PubMed]
- MacHardy, W.E.; Beckman, C.H. Vascular wilt Fusaria: Infection and pathogenesis. In Fusarium: Disease, Biology, and Taxonomy; Nelson, P.E., Toussoun, T.A., Eds.; Pennsylvania State University Press: University Park, PA, USA, 1981; pp. 365–366. [Google Scholar]
- De Sain, M.; Rep, M. The role of pathogen-secreted proteins in fungal vascular wilt diseases. Int. J. Mol. Sci. 2015, 16, 23970–23993. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, A.; Rep, M.; Wang, B.; Ashton, A.; Dodds, P.; Ellis, J. Variation in potential effector genes distinguishing Australian and non-Australian isolates of the cotton wilt pathogen Fusarium oxysporum f. sp. vasinfectum. Plant Pathol. 2011, 60, 232–243. [Google Scholar] [CrossRef]
- Eskalen, A.; Gonzalez, A.; Wang, D.H.; Twizeyimana, M.; Mayorquin, M.; Lynch, S.C. First report of Fusarium sp. and its vector tea shot hole borer (Euwallacea fornicatus) causing Fusarium dieback on avocado in California. Plant Dis. 2012, 96, 1070. [Google Scholar] [CrossRef]
- Eskalen, A.; Stouthamer, R.; Lynch, S.C.; Rugman-Jones, P.F.; Twizeyimana, M. Host range of Fusarium dieback and its ambrosia beetle (Coleoptera: Scolytinae) vector in southern California. Plant Dis. 2013, 97, 938–951. [Google Scholar] [CrossRef]
- Mendel, Z.; Protosav, A.; Sharon, M.; Zveibil, A.; Ben Yahuda, S.; O’Donnell, K.; Rabaglia, R.; Wysoki, M.; Freeman, S. An Asian ambrosia beetle Euwallacea fornicatus and its novel symbiotic fungus Fusarium sp. pose a serious threat to Israeli avocado industry. Phytoparasitica 2012. [Google Scholar] [CrossRef]
- Yadeta, K.A.; Thomma, B.P. The xylem as battleground for plant hosts and vascular wilt pathogens. Front. Plant Sci. 2013, 4, 97. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186–194. [Google Scholar] [CrossRef] [PubMed]
- López-Berges, M.S.; Hera, C.; Sulyok, M.; Schäfer, K.; Capilla, J.; Guarro, J.; Di Pietro, A. The velvet complex governs mycotoxin production and virulence of Fusarium oxysporum on plant and mammalian hosts. Mol. Microbiol. 2013, 87, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Kazan, K.; Gardiner, D.M. Transcriptomics of cereal-Fusarium graminearum interactions: What we have learned so far. Mol. Plant Pathol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Osborne, L.E.; Stein, J.M. Epidemiology of Fusarium head blight on small grain cereals. Int. J. Food Microbiol. 2007, 119, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Del Ponte, E.M.; Valent, B.; Bergstrom, G.C. A special issue on Fusarium head blight and wheat blast. Trop. Plant Pathol. 2017, 42, 143–145. [Google Scholar] [CrossRef]
- Pestka, J.J.; Smolinski, A.T. Deoxynivalenol: Toxicology and potential effects on humans. J. Toxicol. Environ. Health Part B Crit. Rev. 2005, 8, 39–69. [Google Scholar] [CrossRef] [PubMed]
- Dweba, C.C.; Figlan, S.; Shimelis, H.A.; Motaung, T.E.; Sydenham, S.; Mwadzingeni, L.; Tsilo, T.J. Fusarium head blight of wheat: Pathogenesis and control strategies. Crop Prot. 2017, 91, 114–122. [Google Scholar] [CrossRef]
- Nesic, K.; Ivanovic, S.; Nesic, V. Fusarial toxins: Secondary metabolites of Fusarium fungi. Rev. Environ. Contam. Toxicol. 2014, 228, 101–120. [Google Scholar] [PubMed]
- Antonissen, G.; Martel, A.; Pasmans, F.; Ducatelle, R.; Verbrugghe, E.; Vandenbroucke, V.; Li, S.; Haesebrouck, F.; Van Immerseel, F.; Croubels, S. The impact of Fusarium mycotoxins on human and animal host susceptibility to infectious diseases. Toxins 2014, 6, 430–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battilani, P.; Pietri, A.; Barbano, C.; Scandolara, A.; Bertuzzi, T.; Marocco, A. Logistic regression modeling of cropping systems to predict fumonisin contamination in maize. J. Agric. Food Chem. 2008, 56, 10433–10438. [Google Scholar] [CrossRef] [PubMed]
- Bani, M.; Rispail, N.; Evidente, A.; Rubiales, D.; Cimmino, A. Identification of the main toxins isolated from Fusarium oxysporum f. sp. pisi Race 2 and their relation with isolates’ pathogenicity. J. Agric. Food Chem. 2014, 62, 2574–2580. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zuo, C.; Deng, G.; Kuang, R.; Yang, Q.; Hu, C.; Sheng, O.; Zhang, S.; Ma, L.; Wei, Y.; et al. Contamination of bananas with beauvericin and fusaric acid produced by Fusarium oxysporum f. sp. cubense. PLoS ONE 2013, 8, e70226. [Google Scholar] [CrossRef] [PubMed]
- Hernandes, L.; Marangon, A.V.; Salci, T.; Svidzinski, T.I.E. Toxic thermoresistant metabolites of Fusarium oxysporum are capable of inducing histopathological alterations in Wistar rats. J. Venom. Anim. Toxins Incl. Trop. Dis. 2012, 18, 144–149. [Google Scholar] [CrossRef]
- De Melo, I.S.; Picccinin, E. Toxic metabolites from culture filtrate of Fusarium oxysporum and its effects on cucumber cells and plantlets. Rev. Microbiol. 1999, 30, 104–106. [Google Scholar] [CrossRef]
- Dong, F.; Qiu, J.; Xu, J.; Yu, M.; Wang, S.; Sun, Y.; Zhang, G.; Shi, J. Effect of environmental factors on Fusarium population and associated trichothecenes in wheat grain grown in Jiangsu province, China. Int. J. Food Microbiol. 2016, 230, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Ok, H.E.; Choi, S.-W.; Chung, S.H.; Kang, Y.-W.; Kim, D.-S.; Chun, H.S. Natural occurrence of type-B trichothecene mycotoxins in Korean cereal-based products. Food Addit. Contam. Part B Surveill. 2011, 4, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Tan, Y.; Liu, N.; Liao, Y.; Sun, C.; Wang, S.; Wu, A. Functional agents to biologically control deoxynivalenol contamination in cereal grains. Front. Microbiol. 2016, 7, 395. [Google Scholar] [CrossRef] [PubMed]
- Maier, F.J.; Miedaner, T.; Salomon, S.; Hadeler, B.; Felk, A.; Lemmens, M.; Schäfer, W. The involvement of trichothecenes in fusarioses of wheat, barley, and maize evaluated by gene disruption of the trichothecene synthase gene in three field isolates of different chemotype and aggressiveness. Mol. Plant Pathol. 2006, 7, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.; Kim, Y.H. Potential reasons for prevalence of Fusarium wilt in Oriental Melon in Korea. Plant Pathol. J. 2017, 33, 249–263. [Google Scholar] [PubMed]
- Silva, J.C.; Bettiol, W. Potential of non-pathogenic Fusarium oxysporum isolates for control of Fusarium wilt of tomato. Fitopatologia Brasileira 2005, 30, 409–412. [Google Scholar] [CrossRef]
- Compant, S.; Duffy, B.; Nowak, J.; Clément, C.; Barka, E.A. Use of plant growth promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action, and future prospects. Appl. Environ. Microbiol. 2005, 7, 4951–4959. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.-T.; Fan, H.-F.; Gao, Y.-Q.; Li, H.; Zhang, A.-L.; Gao, J.-M. Natural products as sources of new fungicides (I): Synthesis and antifungal activity of acetophenone derivatives against phytopathogenic fungi. Chem. Biol. Drug Res. 2013, 81, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Uesugi, Y. Fungicide Classes: Chemistry, Uses and Mode of Action. In Fungicidal Activity: Chemical and Biological Approaches to Plant Protection; Hutson, D., Miyamoto, J., Eds.; John Wiley & Sons, Ltd.: New York, NY, USA, 1998; pp. 23–56. [Google Scholar]
- Everts, K.L.; Himmelstein, J.C. Fusarium wilt of watermelon: Towards sustainable management of a re-emerging plant disease. Crop Prot. 2015, 73, 93–99. [Google Scholar] [CrossRef]
- Minuto, A.; Gullino, M.L.; Lamberti, F.; D’Addabbo, T.; Tescari, E.; Garibaldi, A. Application of an emulsifiable mixture of 1,3-dichloropropene and chloropicrin against root knot nematodes and soilborne fungi for greenhouse tomatoes in Italy. Crop Prot. 2006, 25, 1244–1252. [Google Scholar] [CrossRef]
- Gilreath, J.P.; Santos, B.M. Efficacy of 1,3-dichloropropene plus chloropicrin incombination with herbicides on purple nutsedge (Cyperus rotundus) control in tomato. Weed Technol. 2004, 19, 137–140. [Google Scholar] [CrossRef]
- McGovern, R.J.; Vavrina, C.S.; Noling, J.S.; Datnoff, L.E.; Yonce, H.D. Evaluation of application methods of metam sodium for management of Fusarium Crown and Root Rot in tomato in southwest Florida. Plant Dis. 1998, 82, 919–923. [Google Scholar] [CrossRef]
- Santos, B.M.; Gilreath, J.P.; Motis, T.N.; Noling, J.W.; Jones, J.P.; Norton, J.A. Comparing methyl bromide alternatives for soilborne disease, nematode and weed management in fresh market tomato. Crop Prot. 2006, 25, 690–695. [Google Scholar] [CrossRef]
- Drinkwater, L.E.; Letourneau, D.K.; Workneh, F.; van Bruggen, A.H.C.; Shennan, C. Fundamental differences between conventional and organic tomato agroecosystems in California. Ecol. Appl. 1995, 5, 1098–1112. [Google Scholar] [CrossRef]
- Heydari, A.; Pessarakli, M. A review on biological control of fungal plant pathogens using microbial antagonists. J. Biol. Sci. 2010, 10, 273–290. [Google Scholar] [CrossRef]
- Arguelles-Arias, A.; Ongena, M.; Halimi, B.; Lara, Y.; Brans, A.; Joris, B.; Fickers, P. Bacillus amyloliquefaciens GA1 as a source of potent antibiotics and other secondary metabolites for biocontrol of plant pathogens. Microb. Cell Fact. 2009, 8, 63. [Google Scholar] [CrossRef] [PubMed]
- Fravel, D.; Olivain, C.; Alabouvette, C. Fusarium oxysporum and its biocontrol. New Phytol. 2003, 157, 493–502. [Google Scholar] [CrossRef]
- Bakkera, M.G.; Gloverb, J.D.; Maib, J.G.; Kinkela, L.L. Plant community effects on the diversity and pathogen suppressive activity of soil streptomycetes. Appl. Soil Ecol. 2010, 46, 35–42. [Google Scholar] [CrossRef]
- Pierson, E.A.; Weller, D.M. Use of mixtures of fluorescent pseudomonads to suppress take-all and improve the growth of wheat. Phytopathology 1994, 84, 940–947. [Google Scholar] [CrossRef]
- Lutz, M.P.; Wenger, S.; Maurhofer, M.; Défago, G.; Duffy, B. Signaling between bacterial and fungal biocontrol agents in a strain mixture. FEMS Microbiol. Ecol. 2004, 48, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Dunlap, C.A.; Schisler, D.A.; Bowman, M.J.; Rooney, A.P. Genomic analysis of Bacillus subtilis OH 131.1 and co-culturing with Cryptococcus flavescens for control of Fusarium head blight. Plant Gene 2015, 2, 1–9. [Google Scholar] [CrossRef]
- Zalila-Kolsi, I.; Mahmoud, A.B.; Ali, H.; Sellami, S.; Nasfi, Z.; Tounsi, S.; Jamoussi, K. Antagonist effects of Bacillus spp. strains against Fusarium graminearum for protection of durum wheat (Triticum turgidum L. sub sp. durum). Microbiol. Res. 2016, 192, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Mishra, A.; Nautiyal, C.S. Paenibacillus lentimorbus B-30488r controls early blight disease in tomato by inducing host resistance associated gene expression and inhibiting Alternaria solani. Biol. Control. 2012, 62, 65–74. [Google Scholar] [CrossRef]
- Whipps, J.M. Microbial interactions and biocontrol in the rhizosphere. J. Exp. Bot. 2001, 511, 487–511. [Google Scholar] [CrossRef]
- Cawoy, H.; Debois, D.; Franzil, L.; De Pauw, E.; Thonart, P.; Ongena, M. Lipopeptides as main ingredients for inhibition of fungal phytopathogens by Bacillus subtilis/amyloliquefaciens. Microb. Biotechnol. 2015, 8, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, R.; Hashem, A.; Abd Allah, E.F. Bacillus: A biological tool for crop improvement through bio-molecular changes in adverse environments. Front. Physiol. 2017, 8, 667. [Google Scholar] [CrossRef] [PubMed]
- Heidarzadeh, N.; Baghaee-Ravari, S. Application of Bacillus pumilus as a potential biocontrol agent of Fusarium wilt of tomato. Arch. Phytopathol. Plant Protect. 2015, 48, 13–16. [Google Scholar] [CrossRef]
- Swiontek, B.M.; Jankiewicz, U.; Burkowska, A.; Walczak, M. Chitinolytic microorganisms and their possible application in environmental protection. Curr. Microbiol. 2014, 68, 71–81. [Google Scholar] [CrossRef] [PubMed]
- DasGupta, S.M.; Khan, N.; Nautiyal, C.S. Biologic control ability of plant growth-promoting Paenibacillus lentimorbus NRRL B-30488 isolated from milk. Curr. Microbiol. 2006, 53, 502–505. [Google Scholar] [CrossRef] [PubMed]
- Swain, R.C.; Ray, R.C.; Nautiyal, C.S. Biocontrol efficacy of Bacillus subtilis strains isolated from cow dung against postharvest yam (Dioscorea rotundata L.) pathogens. Curr. Microbiol. 2008, 57, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Selvaraj, J.N.; Xing, F.; Zhou, L.; Wang, Y.; Song, H.; Tan, X.; Sun, L.; Sangare, L.; Folly, Y.M.E.; et al. Antagonistic action of Bacillus subtilis strain SG6 on Fusarium graminearum. PLoS ONE 2014, 9, e92486. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, E.Z. Chitinase production by Bacillus thuringiensis and Bacillus licheniformis: Their potential in antifungal biocontrol. J. Microbiol. 2012, 50, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Raza, W.; Shen, Q.; Huang, Q. Antifungal activity of Bacillus amyloliquefaciens NJN-6 volatile compounds against Fusarium oxysporum f. sp. cubense. Appl. Environ. Microbiol. 2012, 78, 5942–5944. [Google Scholar] [CrossRef] [PubMed]
- Akram, W.; Anjum, T.; Ali, B.; Ahmad, A. Screening of native Bacillus strains to induce systemic resistance in tomato plants against Fusarium wilt in split root system and its field applications. Int. J. Agric. Biol. 2013, 15, 1289–1294. [Google Scholar]
- Akram, W.; Anjum, T.; Ali, B. Phenylacetic acid is ISR determinant produced by Bacillus fortis IAGS162, which involves extensive re-modulation in metabolomics of tomato to protect against Fusarium wilt. Front. Plant Sci. 2016, 7, 498. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, A.; Ortiz, I.; Maymon, M.; Herbold, C.; Fujishige, N.; Vijanderan, J.; Villella, W.; Hanamoto, K.; Diener, A.; Sanders, E.; et al. Bacillus simplex—A little known PGPB with anti-fungal activity-alters pea legume root architecture and nodule morphology when coinoculated with Rhizobium leguminosarum bv. viciae. Agronomy 2013, 3, 595–620. [Google Scholar] [CrossRef]
- Michiko, M.N.; Zuber, P. Molecular biology of antibiotic production in Bacillus. Crit. Rev. Biotechnol. 1990, 10, 223–240. [Google Scholar]
- Abriouel, H.; Franz, C.; El Bakali, N.; Gálvez, A. Diversity and applications of Bacillus bacteriocins. FEMS Microbiol. Rev. 2011, 35, 201–232. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, J.; Caetano, T.; Mendo, S. Class I and Class II lanthipeptides produced by Bacillus spp. J. Nat. Prod. 2015, 78, 2850–2866. [Google Scholar] [CrossRef] [PubMed]
- Inès, M.; Dhouha, G. Lipopeptide surfactants: Production, recovery and pore forming capacity. Peptides 2015, 71, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Stein, T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions. Mol. Microbiol. 2005, 56, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Hamley, I.W. Lipopeptides: From self-assembly to bioactivity. Chem. Commun. 2015, 51, 8574–8583. [Google Scholar] [CrossRef] [PubMed]
- Geissler, M.; Oellig, C.; Moss, K.; Schwack, W.; Henkel, M.; Hausmann, R. High-performance thin-layer chromatography (HPTLC) for the simultaneous quantification of the cyclic lipopeptides surfactin, iturin A and fengycin in culture samples of Bacillus species. J. Chromatogr. B 2017, 1044–1045, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Sandrin, C.; Peypoux, F.; Michel, G. Coproduction of surfactin and iturin A, lipopeptides with surfactant and antifungal properties, by Bacillus subtilis. Biotechnol. Appl. Biochem. 1990, 12, 370–375. [Google Scholar] [PubMed]
- Maget-Dana, R.; Thimon, L.; Peypoux, F.; Ptak, M. Surfactin/iturin A interactions may explain the synergistic effect of surfactin on the biological properties of iturin A. Biochimie 1992, 74, 1047–1051. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Bruijn, I.D.; Nybroe, O.; Ongena, M. Natural functions of lipopeptides from Bacillus and Pseudomonas: More than surfactants and antibiotics. FEMS Microbiol. Rev. 2010, 34, 1037–1062. [Google Scholar] [CrossRef] [PubMed]
- Vitullo, D.; Di Pietro, A.; Romano, A.; Lanzotti, V.; Lima, G. Role of new bacterial surfactins in the antifungal interaction between Bacillus amyloliquefaciens and Fusarium oxysporum. Plant Pathol. 2012, 61, 689–699. [Google Scholar] [CrossRef]
- Gong, A.D.; Li, H.P.; Yuan, Q.S.; Song, X.S.; Yao, W.; He, W.J.; Zhang, J.B.; Liao, Y.C. Antagonistic Mechanism of iturin A and plipastatin A from Bacillus amyloliquefaciens S76-3 from wheat spikes against Fusarium graminearum. PLoS ONE 2015, 10, e0116871. [Google Scholar] [CrossRef] [PubMed]
- Blacutt, A.A.; Mitchell, T.R.; Bacon, C.W.; Gold, S.E. Bacillus mojavensis RRC101 lipopeptides provoke physiological and metabolic changes during antagonism against Fusarium verticillioides. MPMI 2016, 29, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Li, Q.; Xu, Z.; Zhang, N.; Shen, Q.; Zhang, R. Responses of beneficial Bacillus amyloliquefaciens SQR9 to different soil borne fungal pathogens through the alteration of antifungal compounds production. Front. Microbiol. 2014, 5, 636. [Google Scholar] [CrossRef] [PubMed]
- Zihalirwa Kulimushi, P.; Argüelles Arias, A.; Franzil, L.; Steels, S.; Ongena, M. Stimulation of fengycin-type antifungal lipopeptides in Bacillus amyloliquefaciens in the presence of the maize fungal pathogen Rhizomucor variabilis. Front. Microbiol. 2017, 8, 850. [Google Scholar] [CrossRef] [PubMed]
- González-Sánchez, M.A.; Pérez-Jimenez, R.M.; Pliego, C.; Ramos, C.; de Vicente, A.; Cazorla, F.M. Biocontrol bacteria selected by a direct plant protection strategy against avocado white root rot show antagonism as a prevalent trait. J. Appl. Microbiol. 2010, 109, 65–78. [Google Scholar] [PubMed]
- Joshi, R.; McSpadden-Gardener, B.B. Identification and characterization of novel genetic markers associated with biological control activities in Bacillus subtilis. Phytopathology 2006, 96, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, D.K.; Johri, B.N. Interactions of Bacillus spp. and plants—With special reference to induced systemic resistance (ISR). Microbiol. Res. 2009, 164, 493–513. [Google Scholar] [CrossRef] [PubMed]
- Guanhua, F.; Junfei, M.; Lihong, W.; Xin, Y.; Jeruei, L.; Zhao, X. Effect of degradation of zearalenone-contaminated feed by Bacillus licheniformis CK1 on post weaning female piglets. Toxins 2016, 8, 300. [Google Scholar]



© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, N.; Maymon, M.; Hirsch, A.M. Combating Fusarium Infection Using Bacillus-Based Antimicrobials. Microorganisms 2017, 5, 75. https://doi.org/10.3390/microorganisms5040075
Khan N, Maymon M, Hirsch AM. Combating Fusarium Infection Using Bacillus-Based Antimicrobials. Microorganisms. 2017; 5(4):75. https://doi.org/10.3390/microorganisms5040075
Chicago/Turabian StyleKhan, Noor, Maskit Maymon, and Ann M. Hirsch. 2017. "Combating Fusarium Infection Using Bacillus-Based Antimicrobials" Microorganisms 5, no. 4: 75. https://doi.org/10.3390/microorganisms5040075
APA StyleKhan, N., Maymon, M., & Hirsch, A. M. (2017). Combating Fusarium Infection Using Bacillus-Based Antimicrobials. Microorganisms, 5(4), 75. https://doi.org/10.3390/microorganisms5040075