Longitudinal Analysis of the Microbiota Composition and Enterotypes of Pigs from Post-Weaning to Finishing

Abstract
:1. Introduction
2. Materials and Methods
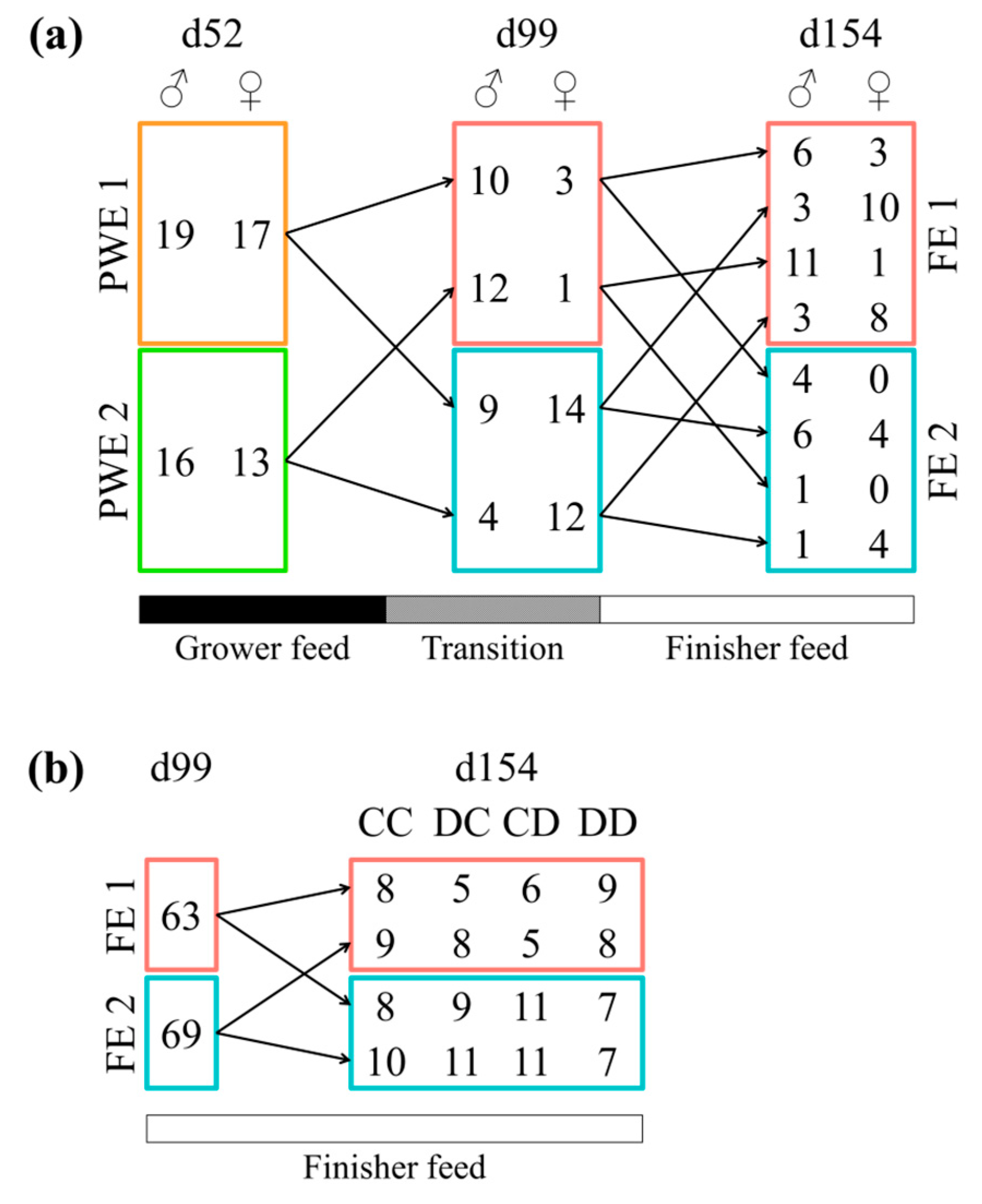
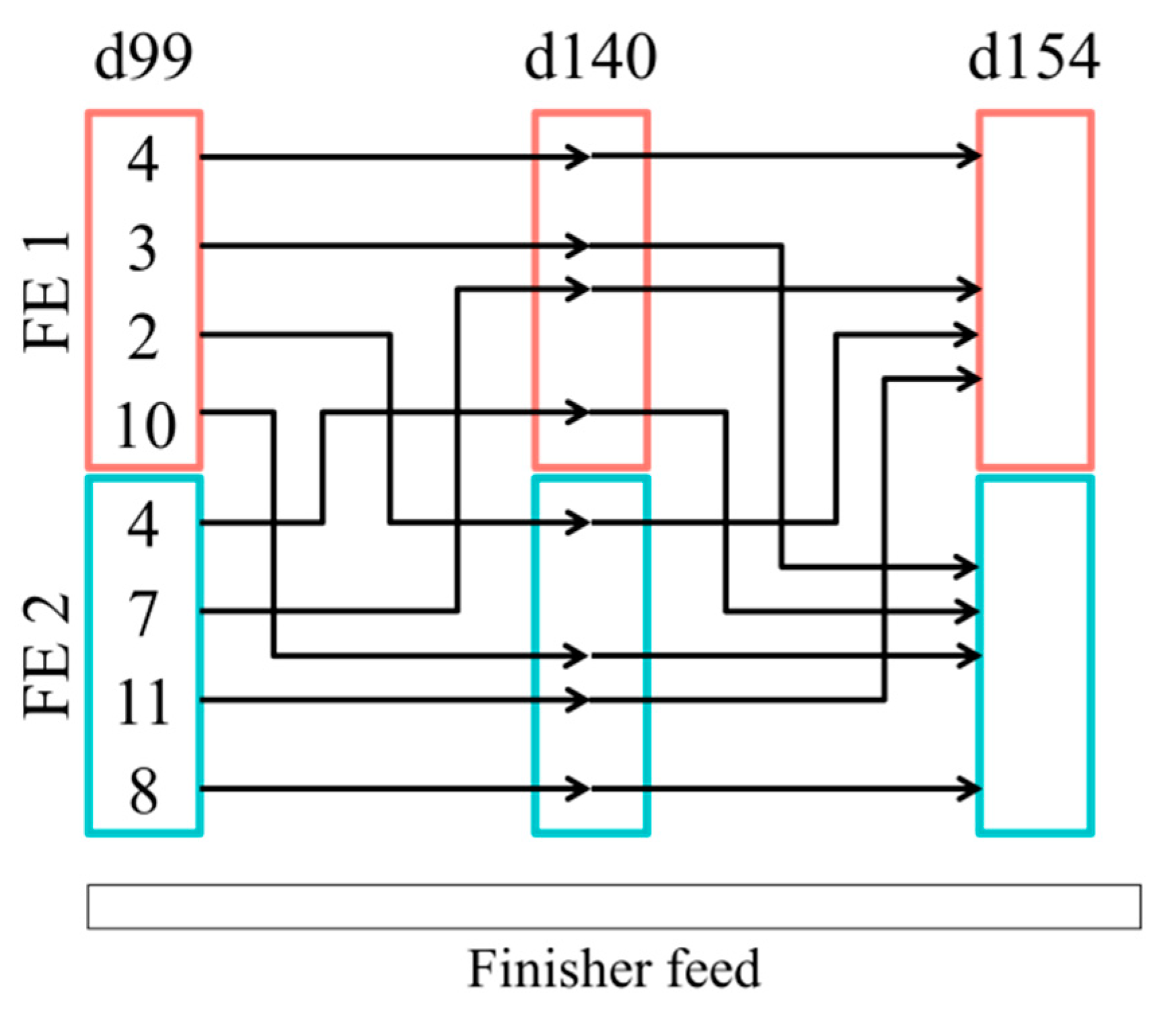
2.1. Animals and Experimental Design
2.2. Measurements
2.3. DNA Sequencing
2.4. Calculations and Statistical Analysis
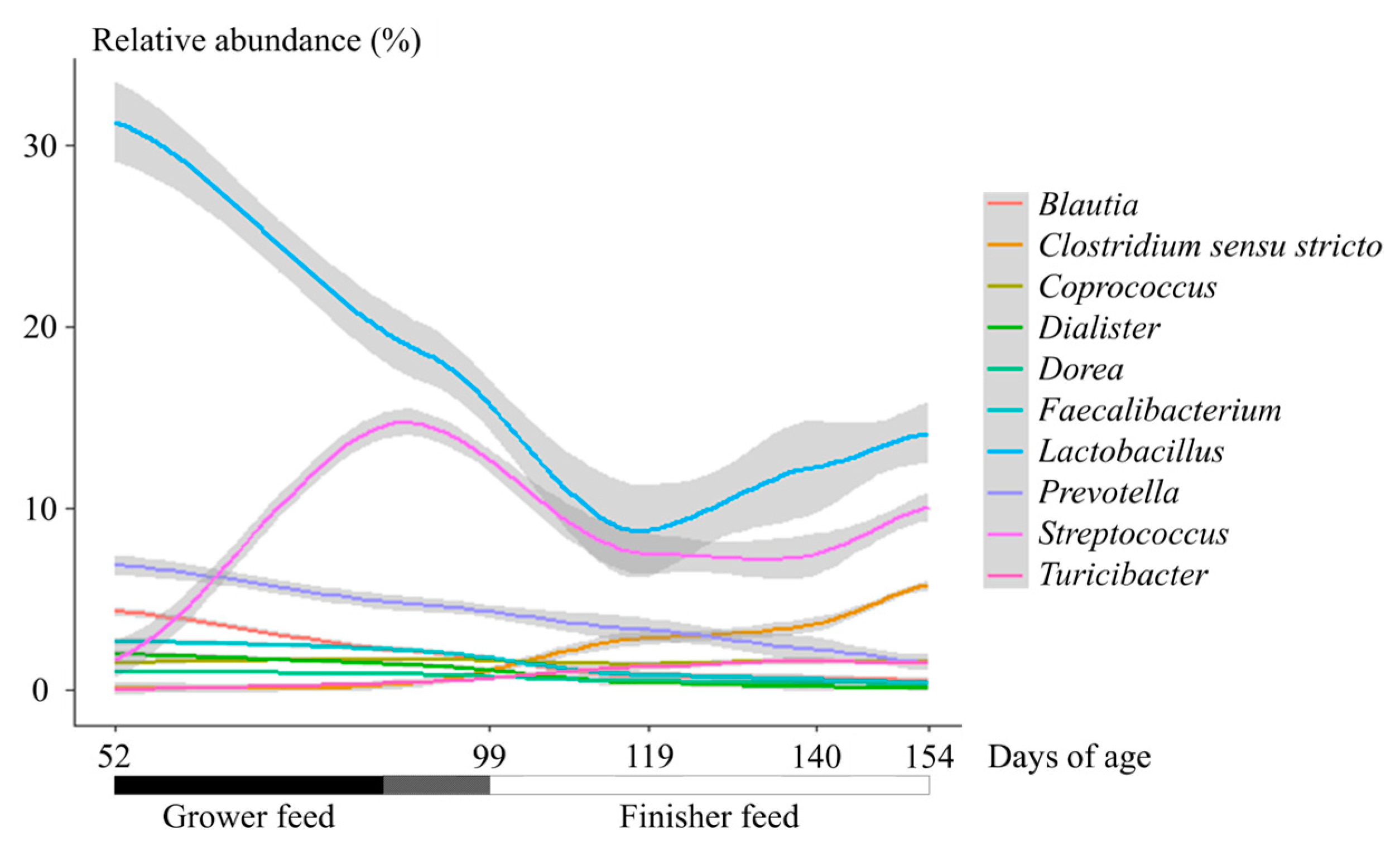
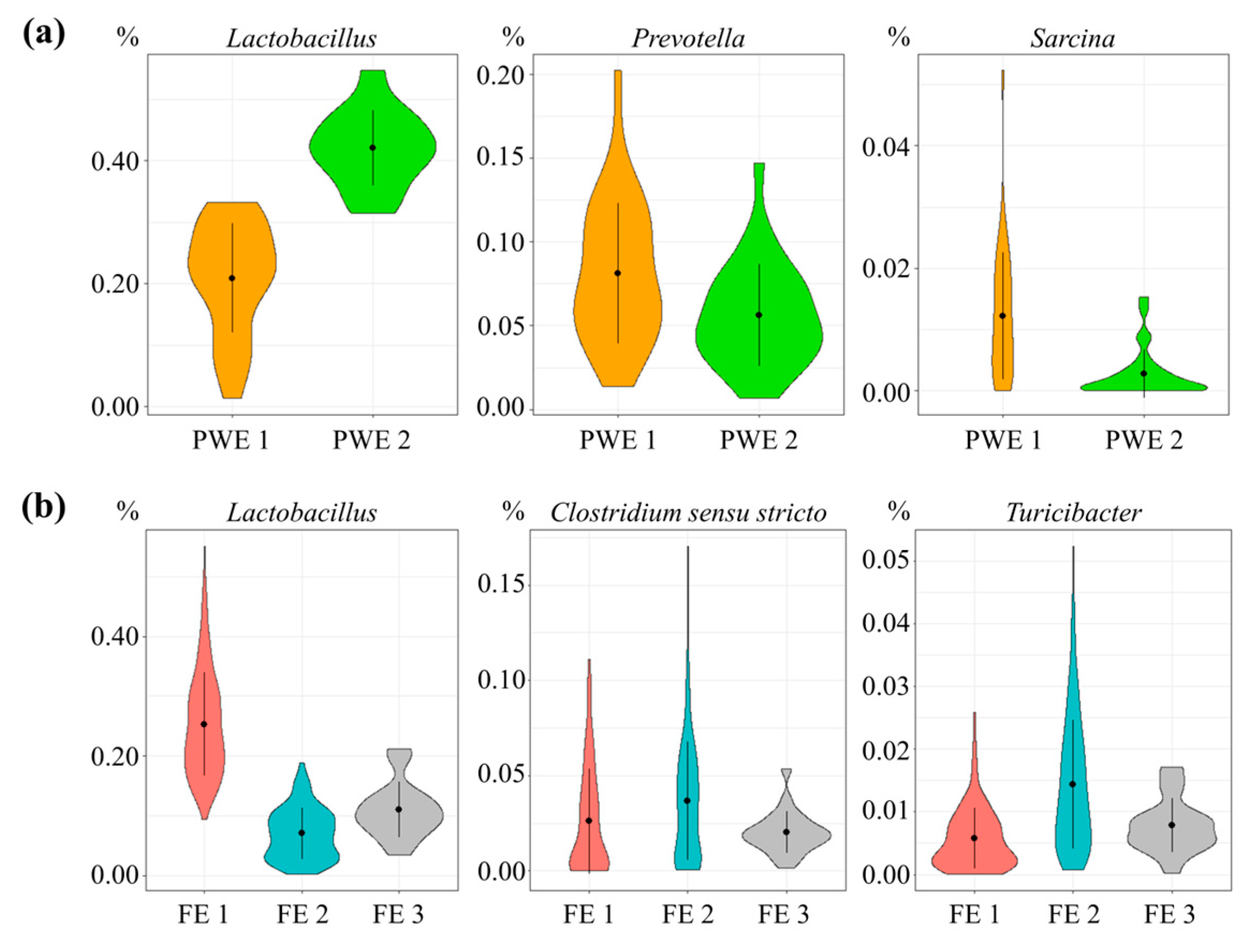
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Xiao, L.; Estellé, J.; Kiilerich, P.; Ramayo-Caldas, Y.; Xia, Z.; Feng, Q.; Liang, S.; Pedersen, A. Øyan; Kjeldsen, N.J.; Liu, C.; et al. A reference gene catalogue of the pig gut microbiome. Nat. Microbiol. 2016, 1, 16161. [Google Scholar] [CrossRef] [PubMed]
- Anguita, M.; Canibe, N.; Pérez, J.F.; Jensen, B.B. Influence of the amount of dietary fiber on the available energy from hindgut fermentation in growing pigs: Use of cannulated pigs and in vitro fermentation. J. Anim. Sci. 2006, 84, 2766–2778. [Google Scholar] [CrossRef] [PubMed]
- Lamendella, R.; Domingo, J.W.S.; Ghosh, S.; Martinson, J.; Oerther, D.B. Comparative fecal metagenomics unveils unique functional capacity of the swine gut. BMC Microbiol. 2011, 11, 103. [Google Scholar] [CrossRef] [PubMed]
- Haenen, D.; Zhang, J.; Da Silva, C.S.; Bosch, G.; Van Der Meer, I.M.; Van Arkel, J.; Borne, J.J.G.C.V.D.; Gutiérrez, O.P.; Smidt, H.; Kemp, B.; et al. A Diet High in Resistant Starch Modulates Microbiota Composition, SCFA Concentrations, and Gene Expression in Pig Intestine. J. Nutr. 2013, 143, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhou, L.; Fang, L.; Su, Y.; Zhu, W. Responses in colonic microbial community and gene expression of pigs to a long-term high resistant starch diet. Front. Microbiol. 2015, 6, 877. [Google Scholar] [CrossRef] [PubMed]
- Le Sciellour, M.; Labussière, E.; Zemb, O.; Renaudeau, D. Effect of dietary fiber content on nutrient digestibility and fecal microbiota composition in growing-finishing pigs. PLoS ONE 2018, 13, e0206159. [Google Scholar] [CrossRef] [PubMed]
- Verschuren, L.M.G.; Calus, M.P.L.; Jansman, A.J.M.; Bergsma, R.; Knol, E.F.; Gilbert, H.; Zemb, O.; Jansman, A.J.M. Fecal microbial composition associated with variation in feed efficiency in pigs depends on diet and sex. J. Anim. Sci. 2018, 96, 1405–1418. [Google Scholar] [CrossRef]
- Niu, Q.; Li, P.; Hao, S.; Kim, S.W.; Du, T.; Hua, J.; Huang, R. Characteristics of Gut Microbiota in Sows and Their Relationship with Apparent Nutrient Digestibility. Int. J. Mol. Sci. 2019, 20, 870. [Google Scholar] [CrossRef]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.-M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef]
- Costea, P.I.; Hildebrand, F.; Arumugam, M.; Bäckhed, F.; Blaser, M.J.; Bushman, F.D.; Fraser, C.M.; Hattori, M.; Huttenhower, C. Enterotypes in the landscape of gut microbial community composition. Nat. Microbiol. 2018, 3, 8–16. [Google Scholar] [CrossRef]
- Mach, N.; Berri, M.; Estellé, J.; Levenez, F.; Lemonnier, G.; Denis, C.; Leplat, J.-J.; Chevaleyre, C.; Billon, Y.; Dore, J.; et al. Early-life establishment of the swine gut microbiome and impact on host phenotypes. Environ. Microbiol. Rep. 2015, 7, 554–569. [Google Scholar] [CrossRef] [PubMed]
- Ramayo-Caldas, Y.; Mach, N.; Lepage, P.; Levenez, F.; Denis, C.; Lemonnier, G.; Leplat, J.-J.; Billon, Y.; Berri, M.; Doré, J.; et al. Phylogenetic network analysis applied to pig gut microbiota identifies an ecosystem structure linked with growth traits. ISME J. 2016, 10, 2973–2977. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Tiezzi, F.; Schillebeeckx, C.; McNulty, N.P.; Schwab, C.; Shull, C.; Maltecca, C. Host contributes to longitudinal diversity of fecal microbiota in swine selected for lean growth. Microbiome 2018, 6, 4. [Google Scholar] [CrossRef]
- Frese, S.A.; Parker, K.; Calvert, C.C.; Mills, D.A. Diet shapes the gut microbiome of pigs during nursing and weaning. Microbiome 2015, 3, 28. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 2 February 2016).
- McMurdie, P.J.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Shields-Cutler, R.R.; Al-Ghalith, G.A.; Yassour, M.; Knights, D. SplinectomeR Enables Group Comparisons in Longitudinal Microbiome Studies. Front. Microbiol. 2018, 9, 785. [Google Scholar] [CrossRef]
- Holman, D.B.; Chénier, M.R. Temporal changes and the effect of subtherapeutic concentrations of antibiotics in the gut microbiota of swine. FEMS Microbiol. Ecol. 2014, 90, 599–608. [Google Scholar] [CrossRef]
- Niu, Q.; Li, P.; Hao, S.; Zhang, Y.; Kim, S.W.; Li, H.; Ma, X.; Gao, S.; He, L.; Wu, W.; et al. Dynamic Distribution of the Gut Microbiota and the Relationship with Apparent Crude Fiber Digestibility and Growth Stages in Pigs. Sci. Rep. 2015, 5, 9938. [Google Scholar] [CrossRef]
- Zhao, W.; Wang, Y.; Liu, S.; Huang, J.; Zhai, Z.; He, C.; Ding, J.; Wang, J.; Wang, H.; Fan, W.; et al. The Dynamic Distribution of Porcine Microbiota across Different Ages and Gastrointestinal Tract Segments. PLoS ONE 2015, 10, e0117441. [Google Scholar] [CrossRef] [PubMed]
- Han, G.G.; Lee, J.-Y.; Jin, G.-D.; Park, J.; Choi, Y.H.; Kang, S.-K.; Chae, B.J.; Kim, E.B.; Choi, Y.-J. Tracing of the fecal microbiota of commercial pigs at five growth stages from birth to shipment. Sci. Rep. 2018, 8, 6012. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.B.; Borewicz, K.; White, B.A.; Singer, R.S.; Sreevatsan, S.; Tu, Z.J.; Isaacson, R.E. Longitudinal investigation of the age-related bacterial diversity in the feces of commercial pigs. Vet. Microbiol. 2011, 153, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Le Sciellour, M.; Zemb, O.; Hochu, I.; Riquet, J.; Gilbert, H.; Giorgi, M.; Billon, Y.; Gourdine, J.-L.; Renaudeau, D. Effect of chronic and acute heat challenges on fecal microbiota composition, production, and thermoregulation traits in growing pigs 1. J. Anim. Sci. 2019, 97, 3845–3858. [Google Scholar] [CrossRef] [PubMed]
- Knights, D.; Ward, T.L.; McKinlay, C.E.; Miller, H.; Gonzalez, A.; McDonald, D.; Knight, R. Rethinking “Enterotypes”. Cell Host Microbe 2014, 16, 433–437. [Google Scholar] [CrossRef]
- Serviento, A.M.; Brossard, L.; Renaudeau, D. An acute challenge with a deoxynivalenol-contaminated diet has short- and long-term effects on performance and feeding behavior in finishing pigs 1. J. Anim. Sci. 2018, 96, 5209–5221. [Google Scholar] [CrossRef]
- Le Sciellour, M.; Zemb, O.; Renaudeau, D. Acute Challenges with a DON-Contaminated Diet Induce Transient Changes on Microbiota Composition in Finishing Pigs; American Society of Animal Science: Austin, TX, USA, 2019. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days of Age | |||||
|---|---|---|---|---|---|
| Item | 52 | 99 | 119 | 140 | 154 |
| Number of samples 1 | 32 | 64 | 60 | 61 | 65 |
| Diversity indexes | |||||
| Nb of OTU | 2 709 ± 323 a | 3 666 ± 363 b | 3 600 ± 334 b | 3 719 ± 288 b | 3 726 ± 298 b |
| Shannon index | 6.67 ± 0.17 a | 7.14 ± 0.16 b | 7.15 ± 0.13 b | 7.19 ± 0.11 b | 7.17 ± 0.11 b |
| Chao1 | 3 707 ± 506 a | 5 229 ± 876 bc | 4 966 ± 731 c | 5 119 ± 583 bc | 5 276 ± 670 b |
| ACE | 3 866 ± 563 a | 5 559 ± 980 b | 5 227 ± 814 c | 5 403 ± 649 bc | 5 602 ± 772 b |
| Simpson | 0.994 ± 0.003 a | 0.997 ± 0.001 b | 0.996 ± 0.001 c | 0.997 ± 0.001 bc | 0.997 ± 0.001 b |
| InvSimpson | 209 ± 64 a | 310 ± 68 b | 284 ± 78 c | 309 ± 75 bc | 324 ± 86 b |
| Fisher | 949 ± 165 a | 1 512 ± 242 b | 1 467 ± 217 b | 1 543 ± 191 b | 1 548 ± 200 b |
| Phyla relative abundance (%) | |||||
| Firmicutes | 72.09 ± 7.61 a | 80.31 ± 5.52 b | 78.15 ± 5.05 b | 78.44 ± 5.89 b | 78.30 ± 6.77 b |
| Bacteroidetes | 22.56 ± 7.82 a | 15.02 ± 5.26 bc | 16.18 ± 5.14 b | 15.17 ± 5.66 bc | 13.26 ± 3.90 c |
| Spirochaetes | 0.34 ± 0.88 a | 0.18 ± 0.28 a | 0.47 ± 0.45 b | 0.70 ± 0.49 c | 1.33 ± 1.28 d |
| Actinobacteria | 0.66 ± 1.12 a | 0.71 ± 1.16 a | 0.26 ± 0.20 b | 0.20 ± 0.09 b | 0.21 ± 0.15 b |
| Proteobacteria | 0.58 ± 0.51 | 0.48 ± 0.34 | 0.44 ± 0.22 | 0.39 ± 0.24 | 0.53 ± 0.33 |
| Fibrobacteres | <0.1 | <0.1 | <0.1 | <0.1 | <0.1 |
| Tenericutes | <0.1 | <0.1 | <0.1 | <0.1 | <0.1 |
| Unclassified | 3.75 ± 2.15 a | 3.28 ± 1.57 a | 4.47 ± 1.89 b | 5.06 ± 1.84 b | 6.29 ± 2.88 c |
| Firmicutes/Bacteroidetes | 3.84 ± 2.10 a | 6.27 ± 3.05 bc | 5.52 ± 2.43 c | 5.98 ± 2.45 b c | 6.66 ± 2.86 b |
| Age at Fecal Sampling (day) 1 | Enterotype | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| FE1 | FE2 | FE3 | ||||||||
| Trait Days of Age | mean | SE | mean | SE | mean | SE | RSD 2 | Tested Factors | Statistics 3 | |
| 99 | No. of pigs | 86 | 97 | |||||||
| ADFI99–113 (kg/day) | 2.45 | 0.04 | 2.42 | 0.04 | - | - | 0.30 | E, R, I, S | I | |
| ADG99–113 (kg/day) | 0.86 | 0.02 | 0.87 | 0.02 | - | - | 0.12 | E, R, I, S | I | |
| FCR99–113 | 2.89 | 0.06 | 2.84 | 0.05 | - | - | 0.44 | E, R, I, S | I | |
| 119 | No. of pigs | 7 | 33 | 26 | ||||||
| ADFI113–119 (kg/day) | 2.08 | 0.12 | 1.97 | 0.06 | 2.12 | 0.07 | 0.31 | E, R, I, D | R, I, D | |
| ADG113–119 (kg/day) | 0.84 ab | 0.08 | 0.65 b | 0.04 | 0.89 a | 0.04 | 0.21 | E, R, I, D | E, D | |
| FCR113–119 | 2.96 | 1.25 | 4.21 | 0.65 | 3.04 | 0.68 | 3.18 | E, R, I, D | D | |
| ADFI119–134 (kg/day) | 2.50 | 0.11 | 2.59 | 0.06 | 2.58 | 0.06 | 0.27 | E, R, I, D | R, I | |
| ADG119–134 (kg/day) | 1.01 | 0.06 | 1.04 | 0.03 | 1.04 | 0.04 | 0.16 | E, R, I, D | I | |
| FCR119–134 | 2.51 | 0.46 | 2.43 | 0.24 | 2.89 | 0.25 | 1.18 | E, R, I, D | - | |
| 140 | No. of pigs | 28 | 44 | |||||||
| ADFI134–140 (kg/day) | 2.61 | 0.07 | 2.55 | 0.06 | - | - | 0.37 | E, R, I, D | I, D | |
| ADG134–140 (kg/day) | 0.91 | 0.04 | 0.91 | 0.04 | - | - | 0.22 | E, R, I, D | I, D | |
| FCR134–140 | 3.21 | 2.23 | 5.36 | 1.86 | - | - | 11.14 | E, R, I, D | - | |
| ADFI140–154 (kg/day) | 3.18 | 0.07 | 3.07 | 0.06 | - | - | 0.35 | E, R, I, D | I | |
| ADG140–154 (kg/day) | 1.12 | 0.04 | 1.10 | 0.03 | - | - | 0.02 | E, R, I, D | D | |
| FCR140–154 | 2.89 | 0.08 | 2.86 | 0.07 | - | - | 0.42 | E, R, I, D | - | |
| 154 | No. of pigs | 72 | 81 | |||||||
| ADFI140–154 (kg/day) | 3.34 | 0.05 | 3.35 | 0.05 | - | - | 0.38 | E, R, I, S | I, S | |
| ADG140–154 (kg/day) | 1.12 | 0.03 | 1.10 | 0.02 | - | - | 0.21 | E, R, I, S | R | |
| FCR140–154 | 3.06 | 0.07 | 3.11 | 0.06 | - | - | 0.48 | E, R, I, S | R, I, S | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le Sciellour, M.; Renaudeau, D.; Zemb, O. Longitudinal Analysis of the Microbiota Composition and Enterotypes of Pigs from Post-Weaning to Finishing. Microorganisms 2019, 7, 622. https://doi.org/10.3390/microorganisms7120622
Le Sciellour M, Renaudeau D, Zemb O. Longitudinal Analysis of the Microbiota Composition and Enterotypes of Pigs from Post-Weaning to Finishing. Microorganisms. 2019; 7(12):622. https://doi.org/10.3390/microorganisms7120622
Chicago/Turabian StyleLe Sciellour, Mathilde, David Renaudeau, and Olivier Zemb. 2019. "Longitudinal Analysis of the Microbiota Composition and Enterotypes of Pigs from Post-Weaning to Finishing" Microorganisms 7, no. 12: 622. https://doi.org/10.3390/microorganisms7120622
APA StyleLe Sciellour, M., Renaudeau, D., & Zemb, O. (2019). Longitudinal Analysis of the Microbiota Composition and Enterotypes of Pigs from Post-Weaning to Finishing. Microorganisms, 7(12), 622. https://doi.org/10.3390/microorganisms7120622