Nutrient Excess Triggers the Expression of the Penicillium chrysogenum Antifungal Protein PAFB



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Media and Cultivation Conditions
2.2. Cloning Vector Construction and Fungal Transformation for Promoter/Terminator Exchange
2.3. Southern Blot Analysis
2.4. Northern Blot Analysis
2.5. Protein Expression, Purification and Verification
2.6. SDS-PAGE and Western Blotting
2.7. Agar Diffusion Test
2.8. Microscopy and Image Processing
2.9. Statistical Analysis
3. Results
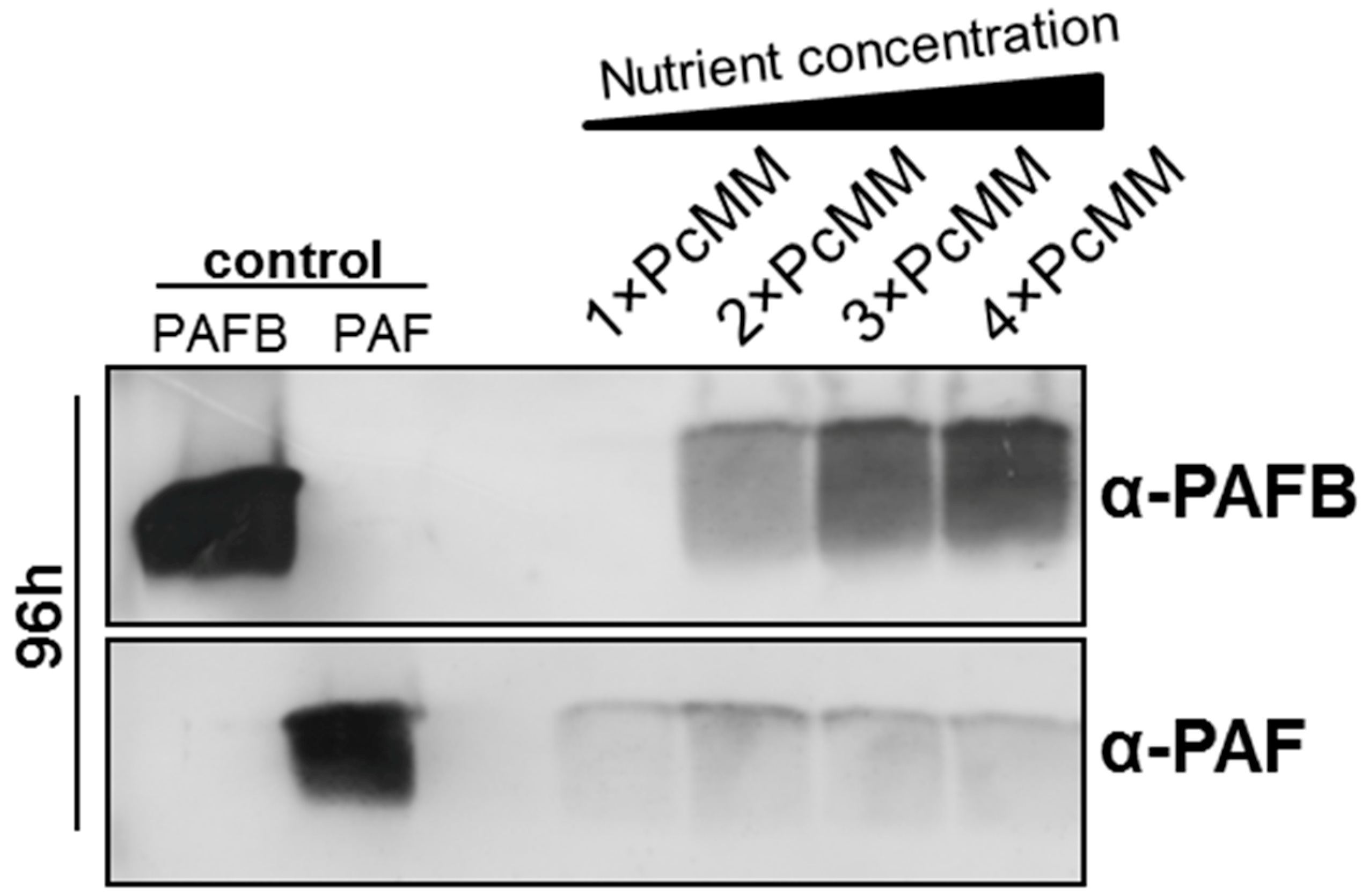
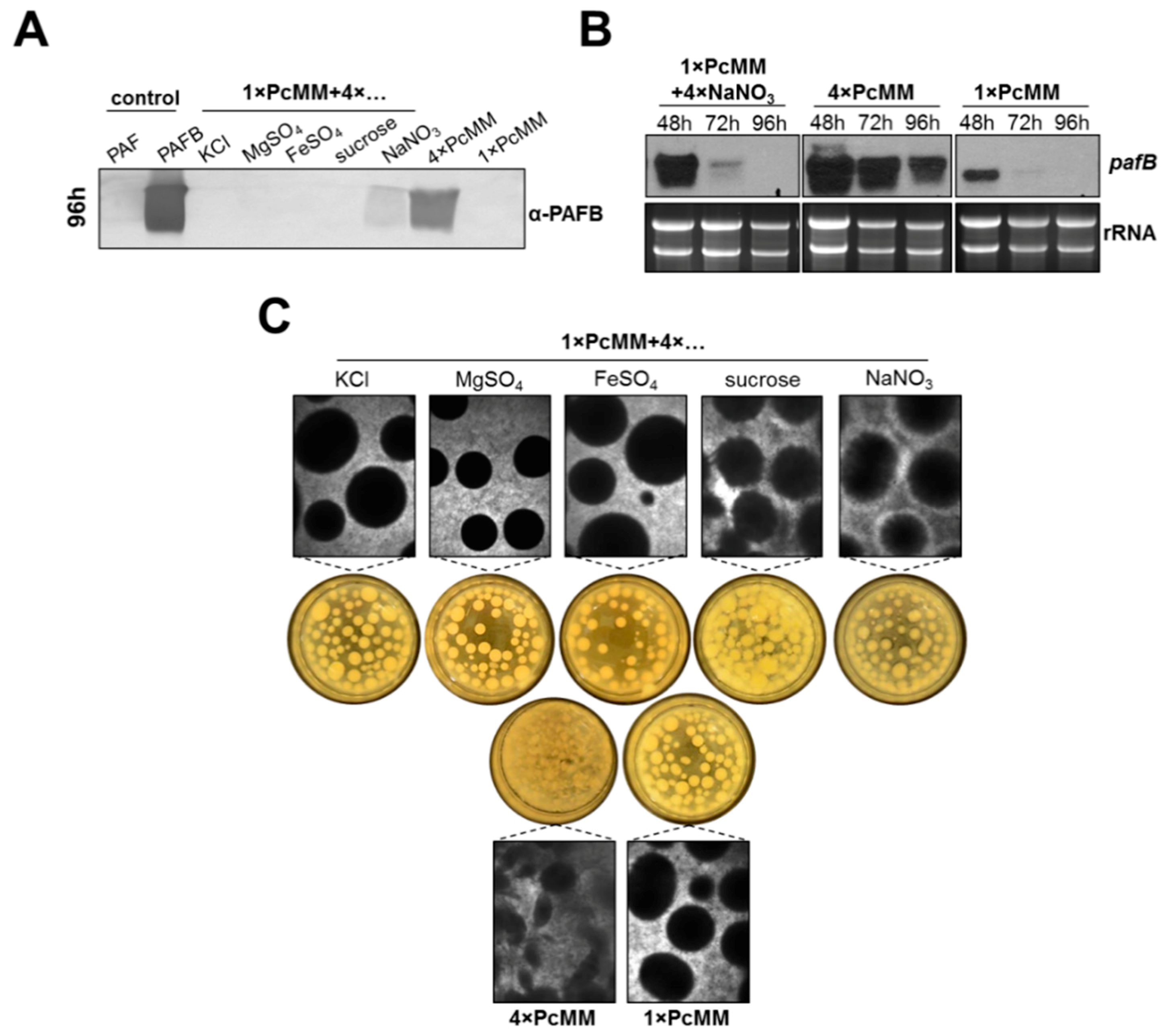
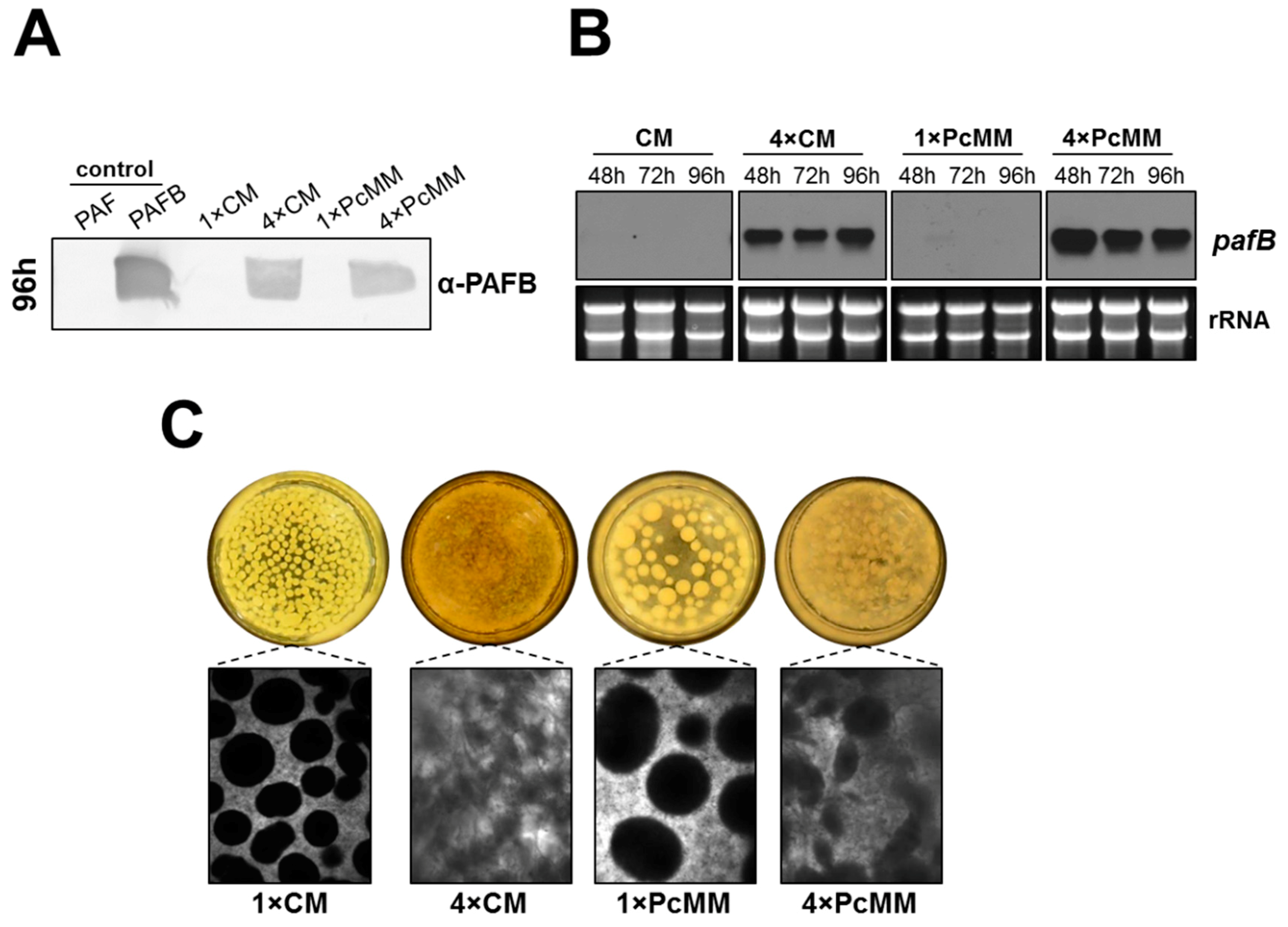
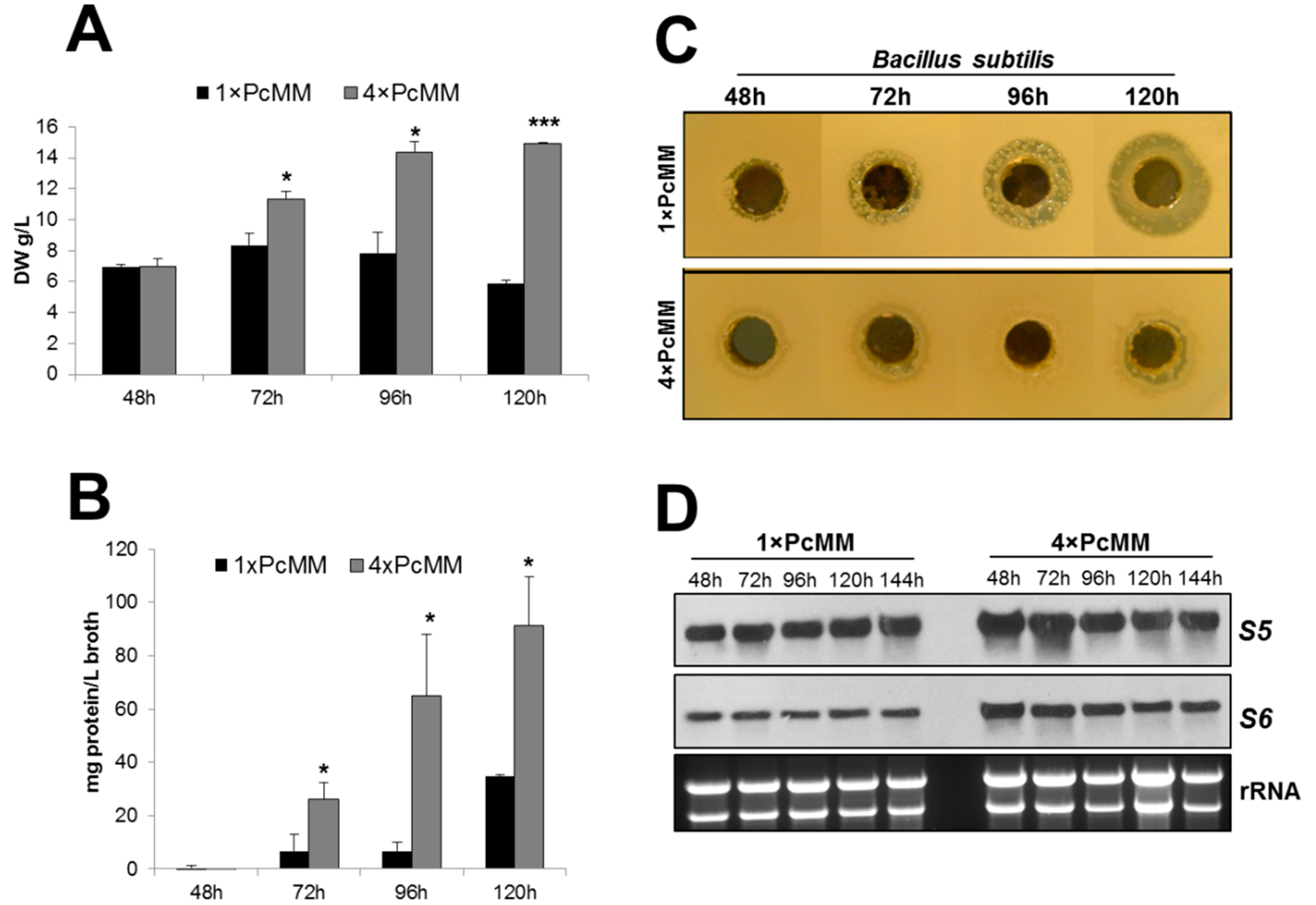
3.1. The Expression of pafB Is Induced by Nutrient Excess
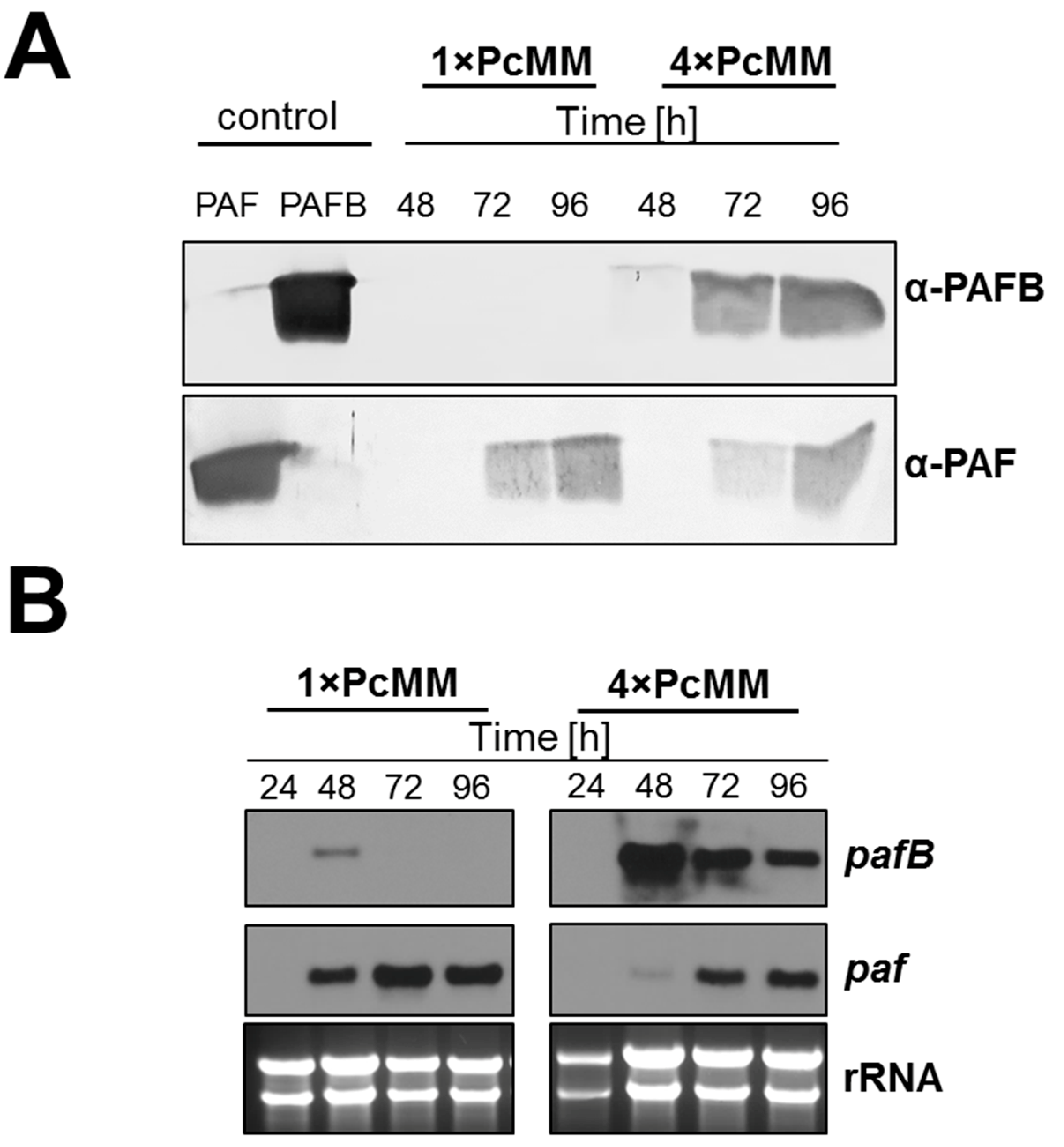
3.2. PAFB Is Expressed during Exponential Growth
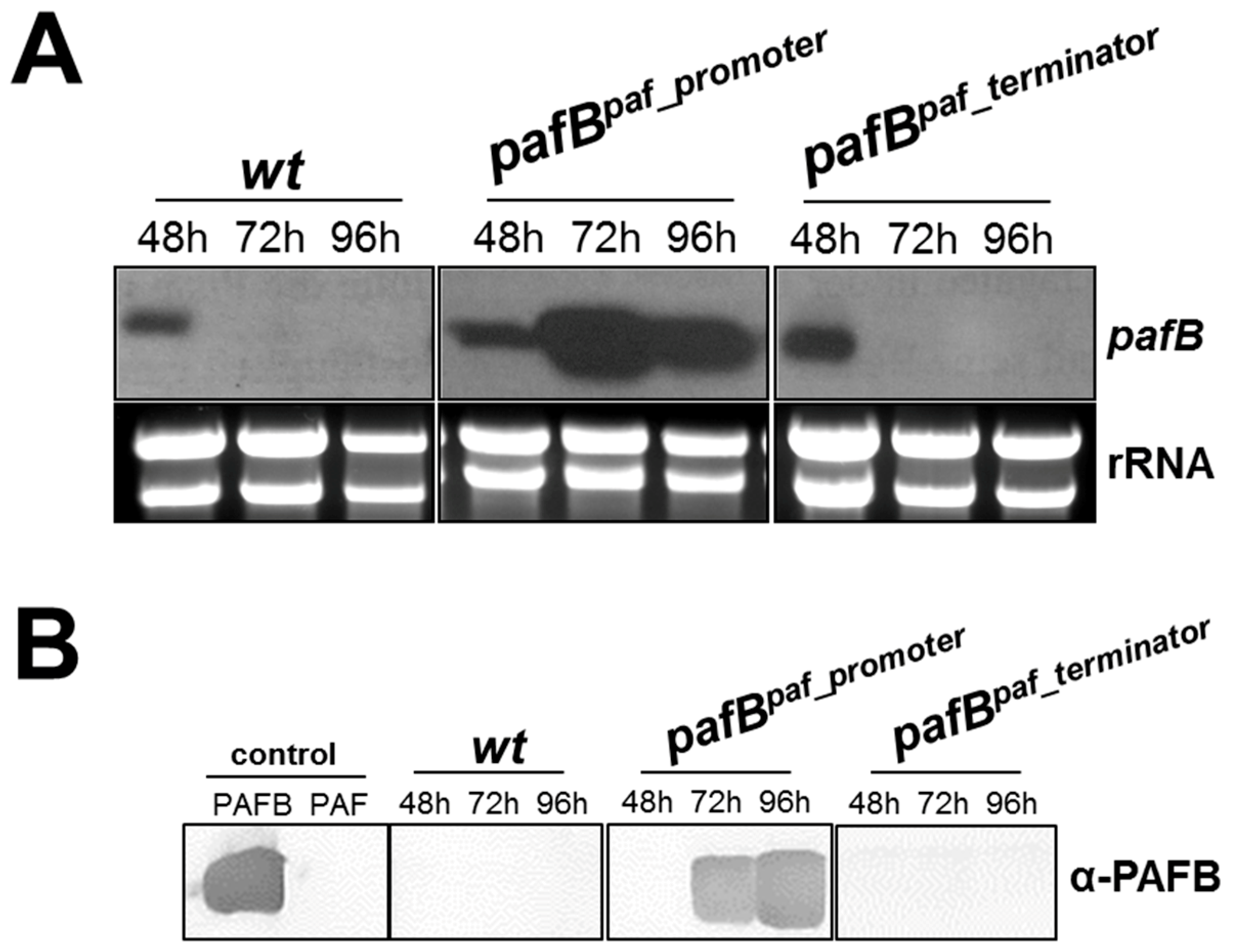
3.3. Increased and Sustained pafB Gene Expression Is Promoter-Regulated
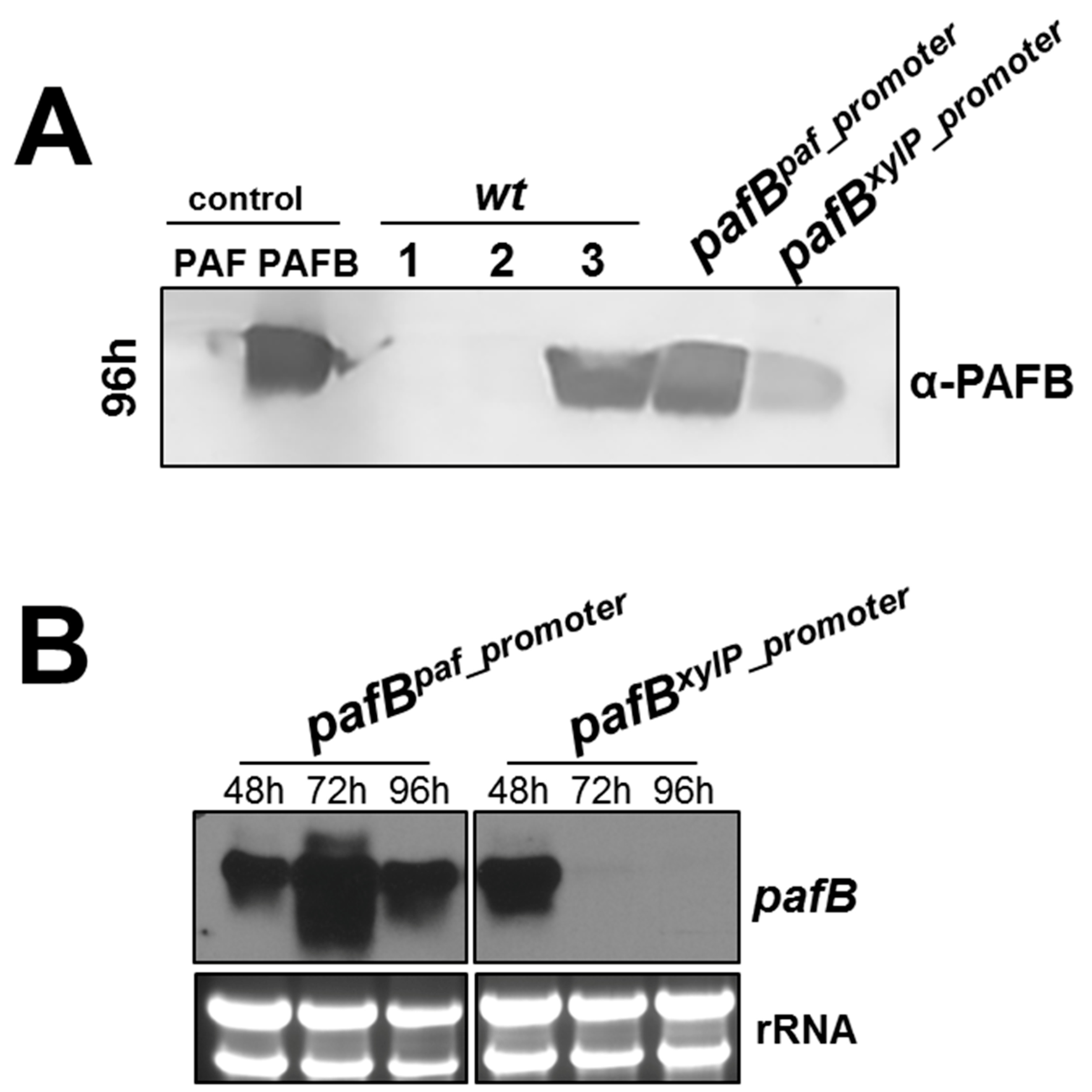
3.4. The pafB-Promoter Is Strong and Inducible
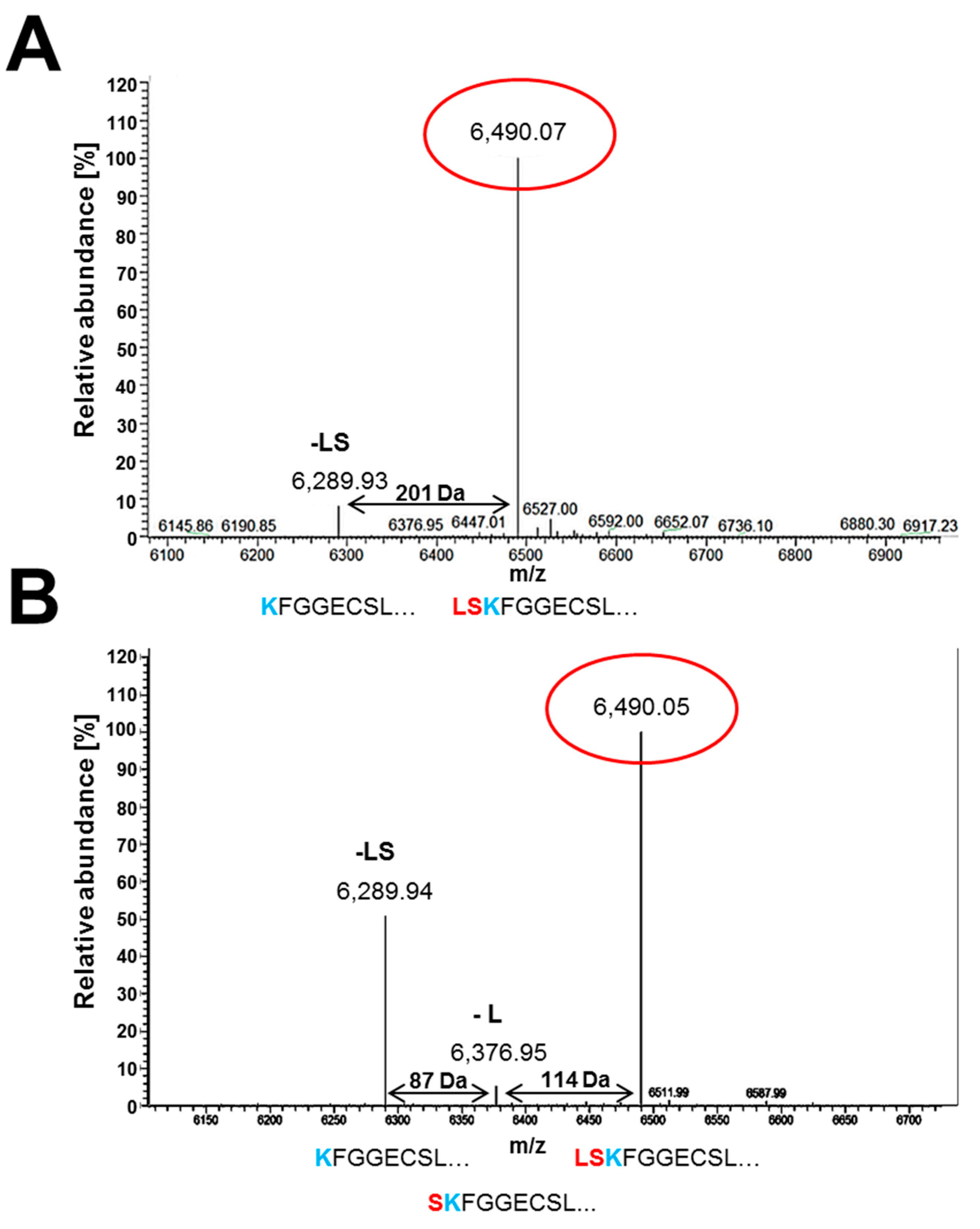
3.5. Secreted PAFB Exhibits Different N-Termini
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sonderegger, C.; Váradi, G.; Galgóczy, L.; Kocsubé, S.; Posch, W.; Borics, A.; Dubrac, S.; Tóth, G.K.; Wilflingseder, D.; Marx, F. The evolutionary conserved gamma-core motif influences the anti-Candida activity of the Penicillium chrysogenum antifungal protein PAF. Front. Microbiol. 2018, 9, 1655. [Google Scholar] [CrossRef] [PubMed]
- Hajdu, D.; Huber, A.; Czajlik, A.; Tóth, L.; Kele, Z.; Kocsubé, S.; Fizil, Á.; Marx, F.; Galgóczy, L.; Batta, G. Solution structure and novel insights into phylogeny and mode of action of the Neosartorya (Aspergillus) fischeri antifungal protein (NFAP). Int. J. Biol. Macromol. 2019, 129, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Galgóczy, L.; Borics, A.; Virágh, M.; Ficze, H.; Váradi, G.; Kele, Z.; Marx, F. Structural determinants of Neosartorya fischeri antifungal protein (NFAP) for folding, stability and antifungal activity. Sci. Rep. 2017, 7, 1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batta, G.; Barna, T.; Gáspári, Z.; Sándor, S.; Kövér, K.E.; Binder, U.; Sarg, B.; Kaiserer, L.; Chhillar, A.K.; Eigentler, A.; et al. Functional aspects of the solution structure and dynamics of PAF--a highly-stable antifungal protein from Penicillium chrysogenum. FEBS J. 2009, 276, 2875–2890. [Google Scholar] [CrossRef] [Green Version]
- Fizil, Á.; Gáspári, Z.; Barna, T.; Marx, F.; Batta, G. “Invisible” conformers of an antifungal disulfide protein revealed by constrained cold and heat unfolding, CEST-NMR experiments, and molecular dynamics calculations. Chemistry 2015, 21, 5136–5144. [Google Scholar] [CrossRef] [Green Version]
- Marx, F.; Haas, H.; Reindl, M.; Stöffler, G.; Lottspeich, F.; Redl, B. Cloning, structural organization and regulation of expression of the Penicillium chrysogenum paf gene encoding an abundantly secreted protein with antifungal activity. Gene 1995, 167, 167–171. [Google Scholar] [CrossRef]
- Huber, A.; Hajdu, D.; Bratschun-Khan, D.; Gáspári, Z.; Varbanov, M.; Philippot, S.; Fizil, Á.; Czajlik, A.; Kele, Z.; Sonderegger, C.; et al. New antimicrobial potential and structural properties of PAFB: A cationic, cysteine-rich protein from Penicillium chrysogenum Q176. Sci. Rep. 2018, 8, 1751. [Google Scholar] [CrossRef]
- Kovács, L.; Virágh, M.; Takó, M.; Papp, T.; Vágvölgyi, C.; Galgóczy, L. Isolation and characterization of Neosartorya fischeri antifungal protein (NFAP). Peptides 2011, 32, 1724–1731. [Google Scholar] [CrossRef]
- Tóth, L.; Kele, Z.; Borics, A.; Nagy, L.G.; Váradi, G.; Virágh, M.; Takó, M.; Vágvölgyi, C.; Galgóczy, L. NFAP2, a novel cysteine-rich anti-yeast protein from Neosartorya fischeri NRRL 181: Isolation and characterization. AMB Express 2016, 6, 75. [Google Scholar] [CrossRef] [Green Version]
- Garrigues, S.; Gandía, M.; Popa, C.; Borics, A.; Marx, F.; Coca, M.; Marcos, J.F.; Manzanares, P. Efficient production and characterization of the novel and highly active antifungal protein AfpB from Penicillium digitatum. Sci. Rep. 2017, 7, 14663. [Google Scholar] [CrossRef] [Green Version]
- Garrigues, S.; Gandía, M.; Castillo, L.; Coca, M.; Marx, F.; Marcos, J.F.; Manzanares, P. Three antifungal proteins from Penicillium expansum: Different patterns of production and antifungal activity. Front. Microbiol. 2018, 9, 2370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacadena, J.; Martínez del Pozo, A.; Gasset, M.; Patiño, B.; Campos-Olivas, R.; Vázquez, C.; Martínez-Ruiz, A.; Mancheño, J.M.; Oñaderra, M.; Gavilanes, J.G. Characterization of the antifungal protein secreted by the mould Aspergillus giganteus. Arch. Biochem. Biophys. 1995, 324, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Wnendt, S.; Ulbrich, N.; Stahl, U. Molecular-cloning, sequence-analysis and expression of the gene encoding an antifungal-protein from Aspergillus giganteus. Curr. Genet. 1994, 25, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.G.; Shin, S.Y.; Maeng, C.Y.; Jin, Z.Z.; Kim, K.L.; Hahm, K.S. Isolation and characterization of a novel antifungal peptide from Aspergillus niger. Biochem. Biophys. Res. Commun. 1999, 263, 646–651. [Google Scholar] [CrossRef]
- Marx, F.; Binder, U.; Leiter, E.; Pócsi, I. The Penicillium chrysogenum antifungal protein PAF, a promising tool for the development of new antifungal therapies and fungal cell biology studies. Cell. Mol. Life Sci. 2008, 65, 445–454. [Google Scholar] [CrossRef]
- Kovács, R.; Holzknecht, J.; Hargital, Z.; Papp, C.; Farkas, A.; Borics, A.; Tóth, L.; Váradi, G.; Tóth, G.K.; Kovács, I.; et al. In Vivo applicability of Neosartorya fischeri antifungal Protein 2 (NFAP2) in treatment of vulvovaginal candidiasis. Antimicrob. Agents Chemother. 2019, 63, e01777-18. [Google Scholar] [CrossRef] [Green Version]
- Szappanos, H.; Szigeti, G.P.; Pál, B.; Rusznák, Z.; Szucs, G.; Rajnavölgyi, E.; Balla, J.; Balla, G.; Nagy, E.; Leiter, E.; et al. The antifungal protein AFP secreted by Aspergillus giganteus does not cause detrimental effects on certain mammalian cells. Peptides 2006, 27, 1717–1725. [Google Scholar] [CrossRef]
- Szappanos, H.; Szigeti, G.P.; Pál, B.; Rusznák, Z.; Szucs, G.; Rajnavölgyi, E.; Balla, J.; Balla, G.; Nagy, E.; Leiter, E.; et al. The Penicillium chrysogenum-derived antifungal peptide shows no toxic effects on mammalian cells in the intended therapeutic concentration. Naunyn-Schmiedebergs Arch. Pharmacol. 2005, 371, 122–132. [Google Scholar] [CrossRef]
- Palicz, Z.; Jenes, A.; Gáll, T.; Miszti-Blasius, K.; Kollár, S.; Kovács, I.; Emri, M.; Márián, T.; Leiter, E.; Pócsi, I.; et al. In Vivo application of a small molecular weight antifungal protein of Penicillium chrysogenum (PAF). Toxicol. Appl. Pharmacol. 2013, 269, 8–16. [Google Scholar] [CrossRef] [Green Version]
- Sonderegger, C.; Galgóczy, L.; Garrigues, S.; Fizil, Á.; Borics, A.; Manzanares, P.; Hegedüs, N.; Huber, A.; Marcos, J.F.; Batta, G.; et al. A Penicillium chrysogenum-based expression system for the production of small, cysteine-rich antifungal proteins for structural and functional analyses. Microb. Cell Fact. 2016, 15, 192. [Google Scholar] [CrossRef] [Green Version]
- Meyer, V.; Wedde, M.; Stahl, U. Transcriptional regulation of the antifungal protein in Aspergillus giganteus. Mol. Genet. Genomics 2002, 266, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Paege, N.; Jung, S.; Schäpe, P.; Müller-Hagen, D.; Ouedraogo, J.P.; Heiderich, C.; Jedamzick, J.; Nitsche, B.M.; van den Hondel, C.A.; Ram, A.F.; et al. A transcriptome meta-analysis proposes novel biological roles for the antifungal protein AnAFP in Aspergillus niger. PLoS ONE 2016, 11, e0165755. [Google Scholar] [CrossRef] [PubMed]
- Hegedüs, N.; Sigl, C.; Zadra, I.; Pócsi, I.; Marx, F. The paf gene product modulates asexual development in Penicillium chrysogenum. J. Basic Microbiol. 2011, 51, 253–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovács, B.; Hegedüs, N.; Bálint, M.; Szabó, Z.; Emri, T.; Kiss, G.; Antal, M.; Pócsi, I.; Leiter, E. Penicillium antifungal protein (PAF) is involved in the apoptotic and autophagic processes of the producer Penicillium chrysogenum. Acta Microbiol. Immunol. Hung. 2014, 61, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Meyer, V.; Jung, S. Antifungal peptides of the AFP family revisited: Are these cannibal toxins? Microorganisms 2018, 6, 50. [Google Scholar] [CrossRef] [Green Version]
- Zadra, I.; Abt, B.; Parson, W.; Haas, H. XylP promoter-based expression system and its use for antisense downregulation of the Penicillium chrysogenum nitrogen regulator NRE. Appl. Environ. Microbiol. 2000, 66, 4810–4816. [Google Scholar] [CrossRef] [Green Version]
- Kück, U.; Hoff, B. Application of the nourseothricin acetyltransferase gene (nat1) as dominant marker for the transformation of filamentous fungi. Fungal Genet. Rep. 2006, 53, 3. [Google Scholar] [CrossRef] [Green Version]
- Sigl, C.; Handler, M.; Sprenger, G.; Kürnsteiner, H.; Zadra, I. A novel homologous dominant selection marker for genetic transformation of Penicillium chrysogenum: Overexpression of squalene epoxidase-encoding ergA. J. Biotechnol. 2010, 150, 307–311. [Google Scholar] [CrossRef]
- Cantoral, J.M.; Díez, B.; Barredo, J.L.; Alvarez, E.; Martín, J.F. High-frequency transformation of Penicillium chrysogenum. Nat. Biotechnol. 1987, 5, 494–497. [Google Scholar] [CrossRef]
- Barreiro, C.; García-Estrada, C. Proteomics and Penicillium chrysogenum: Unveiling the secrets behind penicillin production. J. Proteom. 2019, 198, 119–131. [Google Scholar] [CrossRef]
- Pandey, S.; Ahmad, T.; Aryal, S.; Rana, B.; Sapkota, B. Penicillin production and history: An overview. IJOMAS 2014, 1, 103–108. [Google Scholar]
- Abraham, E.P.; Chain, E. An enzyme from bacteria able to destroy penicillin. Rev. Infect. Dis. 1988, 10, 677–678. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Troya, S.; Pérez-Pérez, M.E.; Florencio, F.J.; Crespo, J.L. The role of TOR in autophagy regulation from yeast to plants and mammals. Autophagy 2008, 4, 851–865. [Google Scholar] [CrossRef]
- Marx, F. Small, basic antifungal proteins secreted from filamentous ascomycetes: A comparative study regarding expression, structure, function and potential application. Appl. Microbiol. Biotechnol. 2004, 65, 133–142. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, J. The filamentous fungal pellet and forces driving its formation. Crit. Rev. Biotechnol. 2016, 36, 1066–1077. [Google Scholar] [CrossRef]
- Gómez, R.; Schnabel, I.; Garrido, J. Pellet growth and citric-acid yield of Aspergillus niger 110. Enzym. Microb. Technol. 1988, 10, 188–191. [Google Scholar] [CrossRef]
- Driouch, H.; Sommer, B.; Wittmann, C. Morphology engineering of Aspergillus niger for improved enzyme production. Biotechnol. Bioeng. 2010, 105, 1058–1068. [Google Scholar] [CrossRef]
- Garrigues, S.; Gandía, M.; Marcos, J.F. Occurrence and function of fungal antifungal proteins: A case study of the citrus postharvest pathogen Penicillium digitatum. Appl. Microbiol. Biotechnol. 2016, 100, 2243–2256. [Google Scholar] [CrossRef]
- Jami, M.S.; Barreiro, C.; García-Estrada, C.; Martín, J.F. Proteome analysis of the penicillin producer Penicillium chrysogenum: Characterization of protein changes during the industrial strain improvement. Mol. Cell Proteom. 2010, 9, 1182–1198. [Google Scholar] [CrossRef] [Green Version]
- Marx, F.; Salvenmoser, W.; Kaiserer, L.; Graessle, S.; Weiler-Görz, R.; Zadra, I.; Oberparleiter, C. Proper folding of the antifungal protein PAF is required for optimal activity. Res. Microbiol. 2005, 156, 35–46. [Google Scholar] [CrossRef]
- Ire, F.S.; Okolo, B.N.; Moneke, A.N.; Odibo, F.J.C. Influence of cultivation conditions on the production of a protease from Aspergillus carbonarius using submerged fermentation. Afr. J. Food Sci. 2011, 5, 353–365. [Google Scholar]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huber, A.; Lerchster, H.; Marx, F. Nutrient Excess Triggers the Expression of the Penicillium chrysogenum Antifungal Protein PAFB. Microorganisms 2019, 7, 654. https://doi.org/10.3390/microorganisms7120654
Huber A, Lerchster H, Marx F. Nutrient Excess Triggers the Expression of the Penicillium chrysogenum Antifungal Protein PAFB. Microorganisms. 2019; 7(12):654. https://doi.org/10.3390/microorganisms7120654
Chicago/Turabian StyleHuber, Anna, Hannah Lerchster, and Florentine Marx. 2019. "Nutrient Excess Triggers the Expression of the Penicillium chrysogenum Antifungal Protein PAFB" Microorganisms 7, no. 12: 654. https://doi.org/10.3390/microorganisms7120654
APA StyleHuber, A., Lerchster, H., & Marx, F. (2019). Nutrient Excess Triggers the Expression of the Penicillium chrysogenum Antifungal Protein PAFB. Microorganisms, 7(12), 654. https://doi.org/10.3390/microorganisms7120654