Investigation on Antibiotic-Resistance, Biofilm Formation and Virulence Factors in Multi Drug Resistant and Non Multi Drug Resistant Staphylococcus pseudintermedius




Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolation and Identification
2.2. DNA Extraction
2.3. Molecular Typing
2.4. Determination of Antibiotic-Resistance Profile
2.4.1. Kirby-Bauer Disk Diffusion Method
2.4.2. Amplification of Antibiotic-Resistance Genes (ARg)
2.5. Biofilm Analysis
2.5.1. Identification of Biofilm-Forming Strains
2.5.2. Amplification of Biofilm-Associated Genes and Agr-Typing
2.6. Virulence Factors Carriage
2.7. Statistical Analysis
3. Results
3.1. Molecular Identification of S. pseudintermedius, MLST, and SCCmec Typing
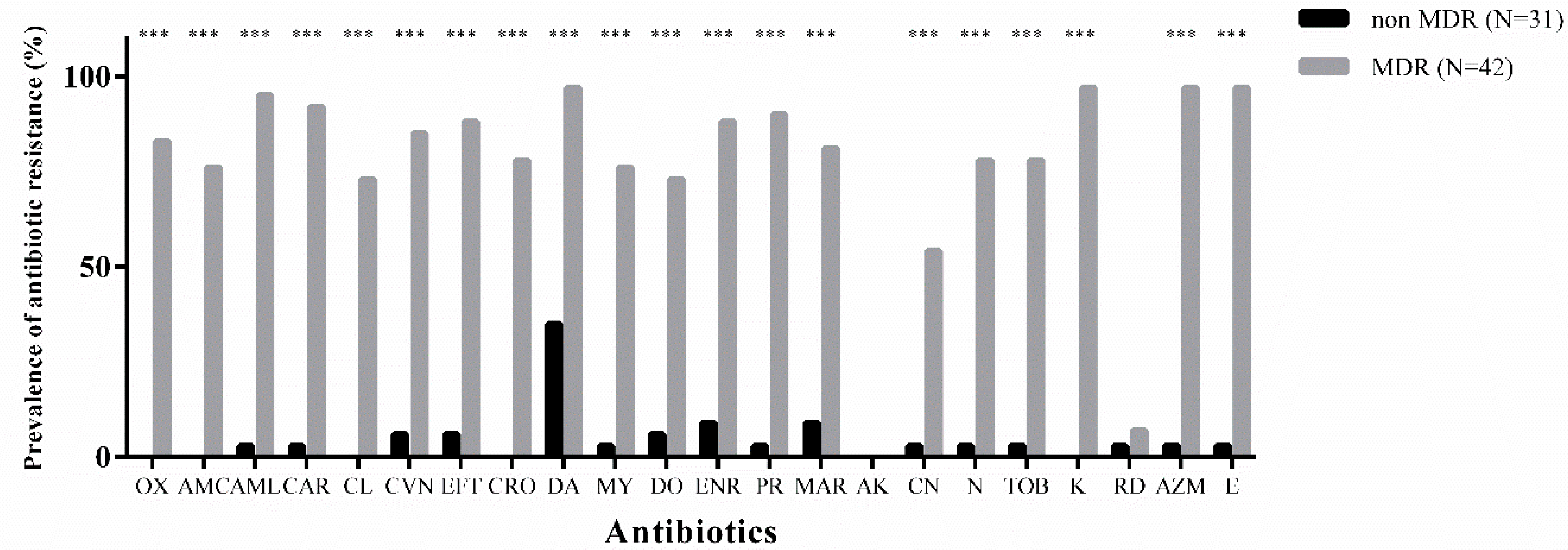
3.2. Overall Antibiotic-Resistance
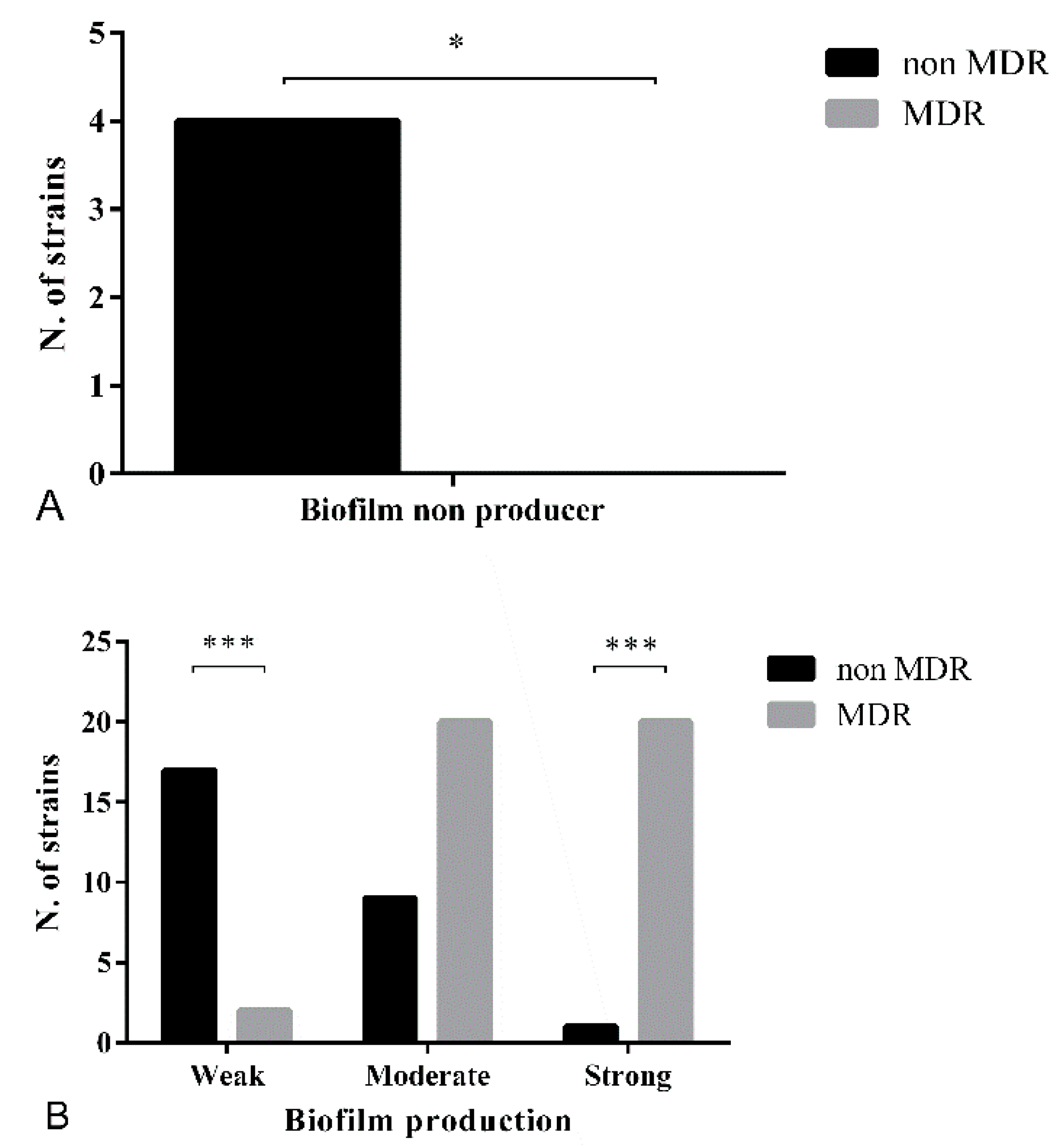
3.3. Biofilm Formation Assay
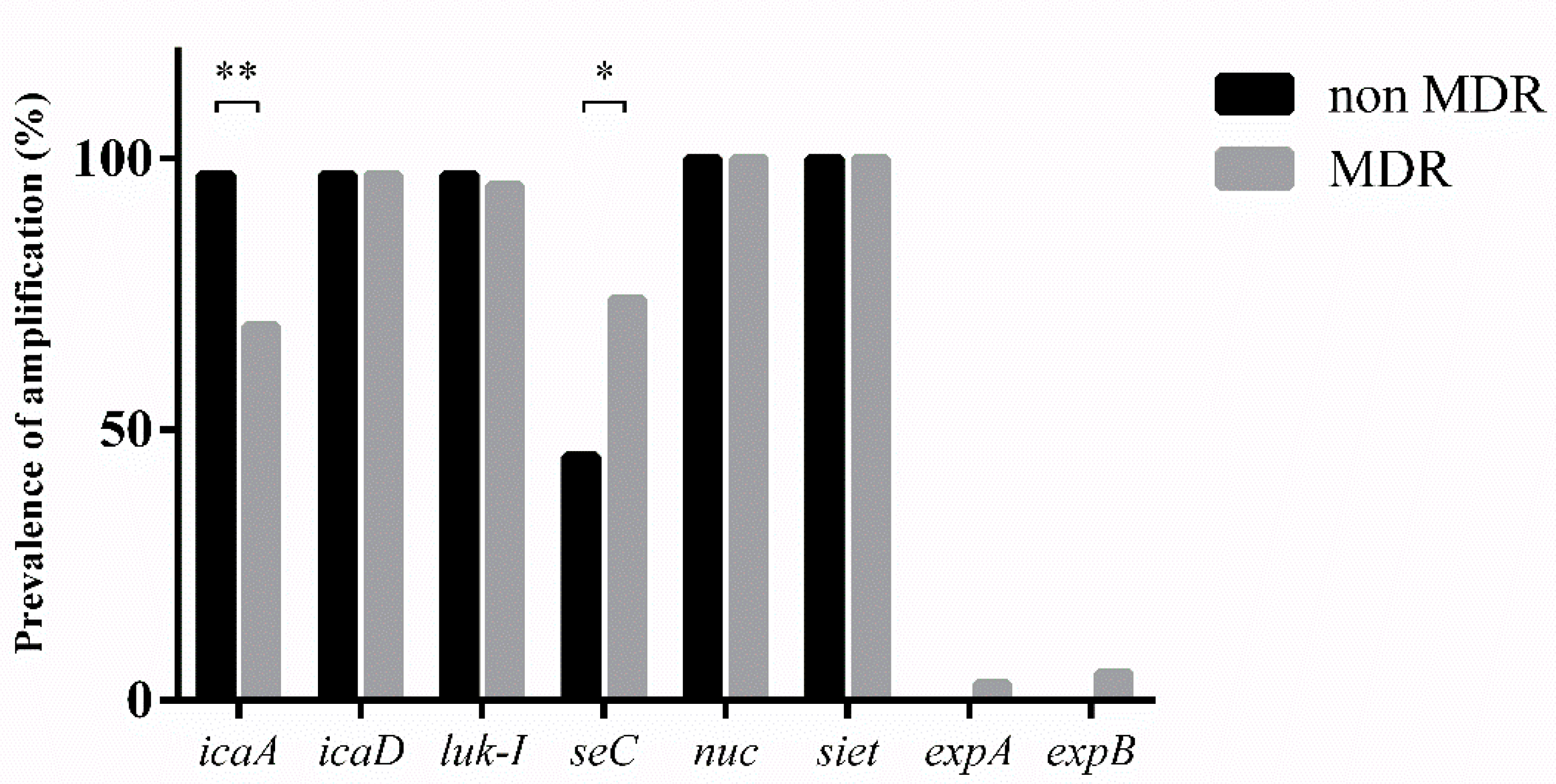
3.4. Virulence Factors
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Devriese, L.A.; Vancanneyt, M.; Baele, M.; Vaneechoutte, M.; De Graef, E.; Snauwaert, C.; Haesebrouck, F. Staphylococcus pseudintermedius sp. nov., a coagulase-positive species from animals. Int. J. Syst. Evol. Microbiol. 2005, 55, 1569–1573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Hoovels, L.; Vankeerberghen, A.; Boel, A.; Van Vaerenbergh, K.; De Beenhouwer, H. First case of Staphylococcus pseudintermedius infection in a human. J. Clin. Microbiol. 2006, 44, 4609–4612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bannoehr, J.; Franco, A.; Iurescia, M.; Battisti, A.; Fitzgerald, J.R. Molecular diagnostic identification of Staphylococcus pseudintermedius. J. Clin. Microbiol. 2009, 47, 469–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guardabassi, L.; Loeber, M.E.; Jacobson, A. Transmission of multiple antimicrobial-resistant Staphylococcus intermedius between dogs affected by deep pyoderma and their owners. Vet. Microbiol. 2004, 98, 23–27. [Google Scholar] [CrossRef]
- Bannoehr, J.; Guardabassi, L. Staphylococcus pseudintermedius in the dog: Taxonomy, diagnostics, ecology, epidemiology and pathogenicity. Vet. Dermatol. 2012, 23, 253-e52. [Google Scholar] [CrossRef]
- Bartlett, S.J.; Rosenkrantz, W.S.; Sanchez, S. Bacterial contamination of commercial ear cleaners following routine home use. Vet. Dermatol. 2011, 22, 546–553. [Google Scholar] [CrossRef]
- Little, S.V.; Bryan, L.K.; Hillhouse, A.E.; Cohen, N.D.; Lawhon, S.D. Characterization of agr Groups of Staphylococcus pseudintermedius Isolates from Dogs in Texas. mSphere 2019, 4, e00033-19. [Google Scholar] [CrossRef] [Green Version]
- Starlander, G.; Börjesson, S.; Grönlund-Andersson, U.; Tellgren-Roth, C.; Melhus, Å. Cluster of Infections Caused by Methicillin-Resistant Staphylococcus pseudintermedius in Humans in a Tertiary Hospital. J. Clin. Microbiol. 2014, 52, 3118–3120. [Google Scholar] [CrossRef] [Green Version]
- Somayaji, R.; Priyantha, M.A.R.; Rubin, J.E.; Church, D. Human infections due to Staphylococcus pseudintermedius, an emerging zoonosis of canine origin: Report of 24 cases. Diagn. Microbiol. Infect. Dis. 2016, 85, 471–476. [Google Scholar] [CrossRef]
- Börjesson, S.; Gómez-Sanz, E.; Ekström, K.; Torres, C.; Grönlund, U. Staphylococcus pseudintermedius can be misdiagnosed as Staphylococcus aureus in humans with dog bite wounds. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 839–844. [Google Scholar] [CrossRef]
- Stefanetti, V.; Bietta, A.; Pascucci, L.; Marenzoni, M.L.; Coletti, M.; Franciosini, M.P.; Passmonti, F.; Casagrandde, P.P. Investigation of the antibiotic resistance and biofilm formation of Staphylococcus pseudintermedius strains isolated from canine pyoderma. Vet. Ital. 2017, 53, 289–296. [Google Scholar]
- Arciola, C.R.; Campoccia, D.; Gamberini, S.; Donati, M.E.; Pirini, V.; Visai, L.; Speziale, P.; Montanaro, L. Antibiotic resistance in exopolysaccharide-forming Staphylococcus epidermidis clinical isolates from orthopaedic implant infections. Biomaterials 2005, 26, 6530–6535. [Google Scholar] [CrossRef]
- Resch, A.; Rosenstein, R.; Nerz, C.; Götz, F. Differential gene expression profiling of Staphylococcus aureus cultivated under biofilm and planktonic conditions. Appl. Environ. Microbiol. 2005, 71, 2663–2676. [Google Scholar] [CrossRef] [Green Version]
- Proietti, P.C.; Stefanetti, V.; Hyatt, D.R.; Marenzoni, M.L.; Capomaccio, S.; Coletti, M.; Bietta, A.; Franciosini, M.P.; Passmonti, F. Phenotypic and genotypic characterization of canine pyoderma isolates of Staphylococcus pseudintermedius for biofilm formation. J. Vet. Med. Sci. 2015, 77, 945–951. [Google Scholar]
- Singh, A.; Walker, M.; Rousseau, J.; Weese, J.S. Characterization of the biofilm forming ability of Staphylococcus pseudintermedius from dogs. BMC Vet. Res. 2013, 9, 93. [Google Scholar] [CrossRef] [Green Version]
- O’Gara, J.P. Ica and beyond: Biofilm mechanisms and regulation in Staphylococcus epidermidis and Staphylococcus aureus. FEMS Microbiol. Lett. 2007, 270, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Panda, S.; Singh, D.V. Biofilm formation by ica-negative ocular isolates of staphylococcus haemolyticus. Front. Microbiol. 2018, 9, 1–9. [Google Scholar] [CrossRef]
- Hennig, S.; Wai, S.N.; Ziebuhr, W. Spontaneous switch to PIA-independent biofilm formation in an ica-positive Staphylococcus epidermidis isolate. Int. J. Med. Microbiol. 2007, 297, 117–122. [Google Scholar] [CrossRef]
- Sasaki, T.; Tsubakishita, S.; Tanaka, Y.; Sakusabe, A.; Ohtsuka, M.; Hirotaki, S.; Kawakami, T.; Fukata, T.; Hiramatsu, K. Multiplex-PCR method for species identification of coagulase-positive staphylococci. J. Clin. Microbiol. 2010, 48, 765–769. [Google Scholar] [CrossRef] [Green Version]
- Adwan, K. Fast DNA isolation and PCR protocols for detection of methicillin-resistant staphylococci. Folia Microbiol. 2014, 59, 5–8. [Google Scholar] [CrossRef]
- Solyman, S.M.; Black, C.C.; Duim, B.; Perreten, V.; Duijkeren, E.V.; Wagenaar, J.A.; Eberlein, L.C.; Sadeghi, L.N.; Videla, R. Multilocus Sequence Typing for Characterization of Staphylococcus pseudintermedius. J. Clin. Microbiol. 2013, 51, 306–310. [Google Scholar] [CrossRef] [Green Version]
- Bannoehr, J.; Zakour, N.L.B.; Waller, A.S.; Guardabassi, L.; Thoday, K.L.; Van Den Broek, A.H.; Fitzgerald, J.R. Population genetic structure of the Staphylococcus intermedius group: Insights into agr diversification and the emergence of methicillin-resistant strains. J. Bacteriol. 2007, 189, 8685–8692. [Google Scholar] [CrossRef] [Green Version]
- Kadlec, K.; Schwarz, S.; Perreten, V.; Grönlund Andersson, U.; Finn, M.; Greko, C.; Moodley, A.; Kania, S.A.; Frank, L.A.; Bemis, D.A. Molecular analysis of methicillin-resistant Staphylococcus pseudintermedius of feline origin from different European countries and North America. J. Antimicrob Chemother. 2010, 65, 1826–1828. [Google Scholar] [CrossRef]
- Perreten, V.; Kadlec, K.; Schwarz, S.; Andersson, U.G.; Finn, M.; Greko, C.; Moodley, A.; Kania, S.A.; Frank, L.A.; Bemis, D.A. Clonal spread of methicillin-resistant Staphylococcus pseudintermedius in Europe and North America: An international multicentre study. J. Antimicrob. Chemother. 2010, 65, 1145–1154. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.H.; Chae, M.J.; Yoon, J.W.; Kim, S.G.; Lee, S.Y.; Yoo, J.H.; Park, H.M. Antibiotic resistance and molecular characterization of ophthalmic Staphylococcus pseudintermedius isolates from dogs. J. Vet. Sci. 2014, 15, 409–415. [Google Scholar] [CrossRef] [Green Version]
- Strommenger, B.; Kettlitz, C.; Werner, G. Multiplex PCR assay for simultaneous detection of nine clinically relevant antibiotic resistance genes in Staphylococcus aureus. J. Clin. Microbiol. 2003, 41, 4089–4094. [Google Scholar] [CrossRef] [Green Version]
- Shopsin, B.; Mathema, B.; Alcabes, P.; Said-Salim, B.; Lina, G.; Matsuka, A. Prevalence of agr specificity groups among Staphylococcus aureus strains colonizing children and their guardians. J. Clin. Microbiol. 2003, 41, 456–459. [Google Scholar] [CrossRef] [Green Version]
- Futagawa-Saito, K.; Sugiyama, T.; Karube, S.; Sakurai, N.; Ba-Thein, W.; Fukuyasu, T. Prevalence and characterization of leukotoxin-producing Staphylococcus intermedius in isolates from dogs and pigeons. J. Clin. Microbiol. 2004, 42, 5324–5326. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.W.; Lee, G.J.; Lee, S.Y.; Park, C.; Yoo, J.H.; Park, H.M. Prevalence of genes for enterotoxins, toxic shock syndrome toxin 1 and exfoliative toxin among clinical isolates of Staphylococcus pseudintermedius from canine origin. Vet. Dermatol. 2010, 21, 484–489. [Google Scholar] [CrossRef]
- Lautz, S.; Kanbar, T.; Alber, J.; Lämmler, C.; Weiss, R.; Prenger-Berninghoff, E.; Zschock, M. Dissemination of the gene encoding exfoliative toxin of Staphylococcus intermedius among strains isolated from dogs during routine microbiological diagnostics. J. Vet. Med. Ser. B Infect. Dis. Vet. Public Health 2006, 53, 434–438. [Google Scholar] [CrossRef]
- Futagawa-Saito, K.; Makino, S.; Sunaga, F.; Kato, Y.; Sakurai-Komada, N.; Ba-Thein, W.; Fukuyasu, T. Identification of first exfoliative toxin in Staphylococcus pseudintermedius. FEMS Microbiol. Lett. 2009, 301, 176–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyori, K.; Hisatsune, J.; Kawakami, T.; Shibata, S.; Murayama, N.; Ide, K.; Nagata, M.; Fukata, T.; Iwasaki, T.; Oshima, K. Identification of a novel Staphylococcus pseudintermedius exfoliative toxin gene and its prevalence in isolates from canines with pyoderma and healthy dogs. FEMS Microbiol. Lett. 2010, 312, 169–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Zubeir, I.E.M.; Kanbar, T.; Alber, J.; Lämmler, C.; Akineden, Ö.; Weiss, R.; Zschock, M. Phenotypic and genotypic characteristics of methicillin/oxacillin-resistant Staphylococcus intermedius isolated from clinical specimens during routine veterinary microbiological examinations. Vet. Microbiol. 2007, 121, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Stepanovic, S.; Vukovic, D.; Dakic, I.; Savic, B.; Svabic-Vlahovic, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Stepanović, S.; Vuković, D.; Hola, V.; Di Bonaventura, G.; Djukić, S.; Cirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. Apmis 2007, 115, 891–899. [Google Scholar] [CrossRef]
- Melchior, M.B.; Van Osch, M.H.J.; Graat, R.M.; Van Duijkeren, E.; Mevius, D.J.; Nielen, M.; Gaastra, W.; Fink-Gremme, J. Biofilm formation and genotyping of Staphylococcus aureus bovine mastitis isolates: Evidence for lack of penicillin-resistance in Agr-type II strains. Vet. Microbiol. 2009, 137, 83–89. [Google Scholar] [CrossRef]
- Landis, J.R.; Koch, G.G. The Measurement of Observer Agreement for Categorical Data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef] [Green Version]
- Kmieciak, W.; Szewczyk, E.M. Are zoonotic Staphylococcus pseudintermedius strains a growing threat for humans? Folia Microbiol. (Praha) 2018, 63, 743–747. [Google Scholar] [CrossRef] [Green Version]
- Somayaji, R.; Rubin, J.E.; Priyantha, M.A.; Church, D. Exploring Staphylococcus pseudintermedius: An emerging zoonotic pathogen? Future Microbiol. 2016, 11, 1371–1374. [Google Scholar] [CrossRef] [Green Version]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Moyaert, H.; de Jong, A.; Simjee, S.; Rose, M.; Youala, M.; Garch, F.E.; Vila, T.; Klein, U.; Rzewuska, M.; Morrissey, I. Survey of antimicrobial susceptibility of bacterial pathogens isolated from dogs and cats with respiratory tract infections in Europe: ComPath results. J. Appl. Microbiol. 2019, 127, 29–46. [Google Scholar] [CrossRef] [PubMed]
- Damborg, P.; Moodley, A.; Aalbæk, B.; Ventrella, G.; dos Santos, T.P.; Guardabassi, L. High genotypic diversity among methicillin-resistant Staphylococcus pseudintermedius isolated from canine infections in Denmark. BMC Vet. Res. 2016, 12, 131–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergot, M.; Martins-Simoes, P.; Kilian, H.; Châtre, P.; Worthing, K.A.; Norris, J.M.; Madec, J.-Y.; Laurent, F.; Haenni, M. Evolution of the Population Structure of Staphylococcus pseudintermedius in France. Front. Microbil. 2018, 9, 3055. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, T.P.; Damborg, P.; Moodley, A.; Guardabassi, L. Systematic review on global epidemiology of methicillin-resistant staphylococcus pseudintermedius: Inference of population structure from multilocus sequence typing data. Front. Microbiol. 2016, 7, 1599. [Google Scholar] [CrossRef] [Green Version]
- Rinsky, J.L.; Nadimpalli, M.; Wing, S.; Hall, D.; Baron, D.; Price, L.B.; Larsen, J.; Stegger, M.; Stewart, J.; Heaney, C.D. Livestock-Associated Methicillin and Multidrug Resistant Staphylococcus aureus Is Present among Industrial, Not Antibiotic-Free Livestock Operation Workers in North Carolina. PLoS ONE 2013, 8, e67641. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Genes | Sequence (5’-3’) | Amplicon Size (bp) | PCR Conditions | References | ||
|---|---|---|---|---|---|---|
| Antibiotic-resistance genes | mecA | F | AAAATCGATGGTAAAGGTTGGC | 532 | 95 °C × 4 min, 30 × (95 °C × 1 min, 58 °C × 1 min, 72 °C × 1 min) 72 °C × 7 min, 4 °C | [25] |
| R | AGTTCTGCAGTACCGGATTTGC | |||||
| blaZ | F | TGACCACTTTTATCAGCAACC | 750 | |||
| R | GCCATTTCAACACCTTCTTTC | |||||
| tetK | F | GTAGCGACAATAGGTAATAGT | 360 | 94 °C × 3 min, 30 × (94 °C × 30 s, 55 °C × 30 s, 72 °C × 30 s) 72 °C × 7 min, 4 °C | [26] | |
| R | GTAGTGACAATAAACCTCCTA | |||||
| tetM | F | AGTGGAGCGATTACAGAA | 158 | |||
| R | CATATGTCCTGGCGTGTCTA | |||||
| aacA-aphD | F | TAATCCAAGAGCAATAAGGGC | 227 | |||
| R | GCCACACTATCATAACCACTA | |||||
| Biofilm genes | icaA | F | ACTGTTTCGGGGACAAGCAT | 134 | 94 °C × 3 min, 35 × (94 °C × 15 s, 60 °C × 20 s, 72 °C × 20 s) 72 °C × 7 min, 4 °C | [14] |
| R | ATTGAGGCTGTAGGGCGTTG | |||||
| icaD | F | CGTTAATGCCTTCTTTCTTATTGCG | 166 | 94 °C × 3 min, 35 × (94 °C × 15 s, 56 °C × 20 s, 72 °C × 20 s) 72 °C × 7 min, 4 °C | ||
| R | ATTAGCGCACATTCGGTGTT | |||||
| Quorum-sensing genes | pan-agr | F | ATGCACATGGTGCACATGC | 94 °C × 3 min, 35 × (94 °C × 15 s, 56 °C × 20 s, 72 °C × 20 s) 72 °C × 7 min, 4 °C | [27] | |
| agrI | R | GTCACAAGTACTATAAGCTGCGAT | ||||
| agrII | R | GTATTACTAATTGAAAAGTGCCATAGC | ||||
| agrIII | R | CTGTTGAAAAAGTCAACTAAAAGCTC | ||||
| agrIV | R | CGATAATGCCGTAATACCCG | ||||
| Virulence factors | luk-F | F | CCTGTCTATGCCGCTAATCCA | 572 | 94 °C × 3 min, 35 × (94 °C × 1 min, 57 °C × 1 min, 72 °C × 1 min) 72 °C × 7 min, 4 °C | [28] |
| R | AGGTCATGGAAGCTATCTCGA | |||||
| luk-S | F | TGTAAGCAGCAGAAAATGGGG | 503 | |||
| R | GCCCGATAGGACTTCTTACAA | |||||
| seC | F | GGCGGCAATATTGGCGCTCG | 271 | 95 °C × 2 min, 30 × (95 °C × 1 min, 55 °C × 1 min, 72 °C × 2 min) 72 °C × 5 min, 4 °C | [29] | |
| R | TTACTGTCAATGCTCTGACC | |||||
| nuc | F | TRGGCAGTAGGATTCGTTAA | 926 | 95 °C × 2 min, 30 × (95 °C × 30 s, 52 °C × 30 s, 72 °C × 30 s) 72 °C × 2 min, 4 °C | [19] | |
| R | CTTTTGTGCTYCMTTTTGG | |||||
| siet | F | ATGGAAAATTTAGCGGCATCTGG | 359 | 94 °C × 3 min, 30 × (94 °C × 30 s, 56 °C × 30 s, 72 °C × 1 min) 72 °C × 5 min, 4 °C | [30] | |
| R | CCATTACTTTTCGCTTGTTGTGC | |||||
| expA | F | GTKTTAATTGGWAAAAATACA | 413 | 94 °C × 3 min, 30 × (94 °C × 1 min, 42 °C × 1 min, 72 °C × 1 min) 72 °C × 4 min, 4°C | [31] | |
| R | ATNCCWGAKCCTGAATTWCC | |||||
| expB | F | GGGCATGCACATATGATGAAGCC | 820 | 95 °C × 3 min, 30 × (95 °C × 1 min, 53 °C × 1 min, 72 °C × 1 min) 72 °C × 4 min, 4 °C | [32] | |
| R | CCAGATCTATCTTCTGATTCAGC |
| MLST | SCCmec Types | No. of Isolates (%) |
|---|---|---|
| ST 71 | II-III | 24 (68.5%) |
| ST 258 | IV | 9 (25.7%) |
| ST 106 | IV | 2 (5.7%) |
| Genes | not MDR Strains | MDR Strains | p-Value | |
|---|---|---|---|---|
| Antibiotic-resistance genes | mecA | 0 | 35/42 (83%) | <0.0001 |
| blaZ | 7/31 (23%) | 42/42 (100%) | <0.0001 | |
| tetK | 2/31 (6.4%) | 13/42 (31%) | 0.0171 | |
| tetM | 0 | 22/42 (50%) | <0.0001 | |
| aacA-aphD | 5/31 (16.6%) | 32/42 (76%) | <0.0001 | |
| Biofilm genes | icaA | 30/31 (97%) | 29/42 (69%) | 0.0026 |
| icaD | 30/31 (97%) | 41/42 (97%) | >0.05 | |
| Virulence factors | luk-I | 30/31 (97%) | 40/42 (95%) | >0.05 |
| seC | 14/31 (45%) | 31/42 (74%) | 0.016 | |
| nuc | 31/31 (100%) | 42/42 (100%) | >0.05 | |
| siet | 31/31 (100%) | 42/42 (100%) | >0.05 | |
| expA | 0 | 3/42 (7%) | >0.05 | |
| expB | 0 | 5/42 (12%) | >0.05 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meroni, G.; Soares Filipe, J.F.; Drago, L.; Martino, P.A. Investigation on Antibiotic-Resistance, Biofilm Formation and Virulence Factors in Multi Drug Resistant and Non Multi Drug Resistant Staphylococcus pseudintermedius. Microorganisms 2019, 7, 702. https://doi.org/10.3390/microorganisms7120702
Meroni G, Soares Filipe JF, Drago L, Martino PA. Investigation on Antibiotic-Resistance, Biofilm Formation and Virulence Factors in Multi Drug Resistant and Non Multi Drug Resistant Staphylococcus pseudintermedius. Microorganisms. 2019; 7(12):702. https://doi.org/10.3390/microorganisms7120702
Chicago/Turabian StyleMeroni, Gabriele, Joel F. Soares Filipe, Lorenzo Drago, and Piera A. Martino. 2019. "Investigation on Antibiotic-Resistance, Biofilm Formation and Virulence Factors in Multi Drug Resistant and Non Multi Drug Resistant Staphylococcus pseudintermedius" Microorganisms 7, no. 12: 702. https://doi.org/10.3390/microorganisms7120702
APA StyleMeroni, G., Soares Filipe, J. F., Drago, L., & Martino, P. A. (2019). Investigation on Antibiotic-Resistance, Biofilm Formation and Virulence Factors in Multi Drug Resistant and Non Multi Drug Resistant Staphylococcus pseudintermedius. Microorganisms, 7(12), 702. https://doi.org/10.3390/microorganisms7120702