Phenotypic and Molecular Analyses of Rhizoctonia spp. Associated with Rice and Other Hosts


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Isolation
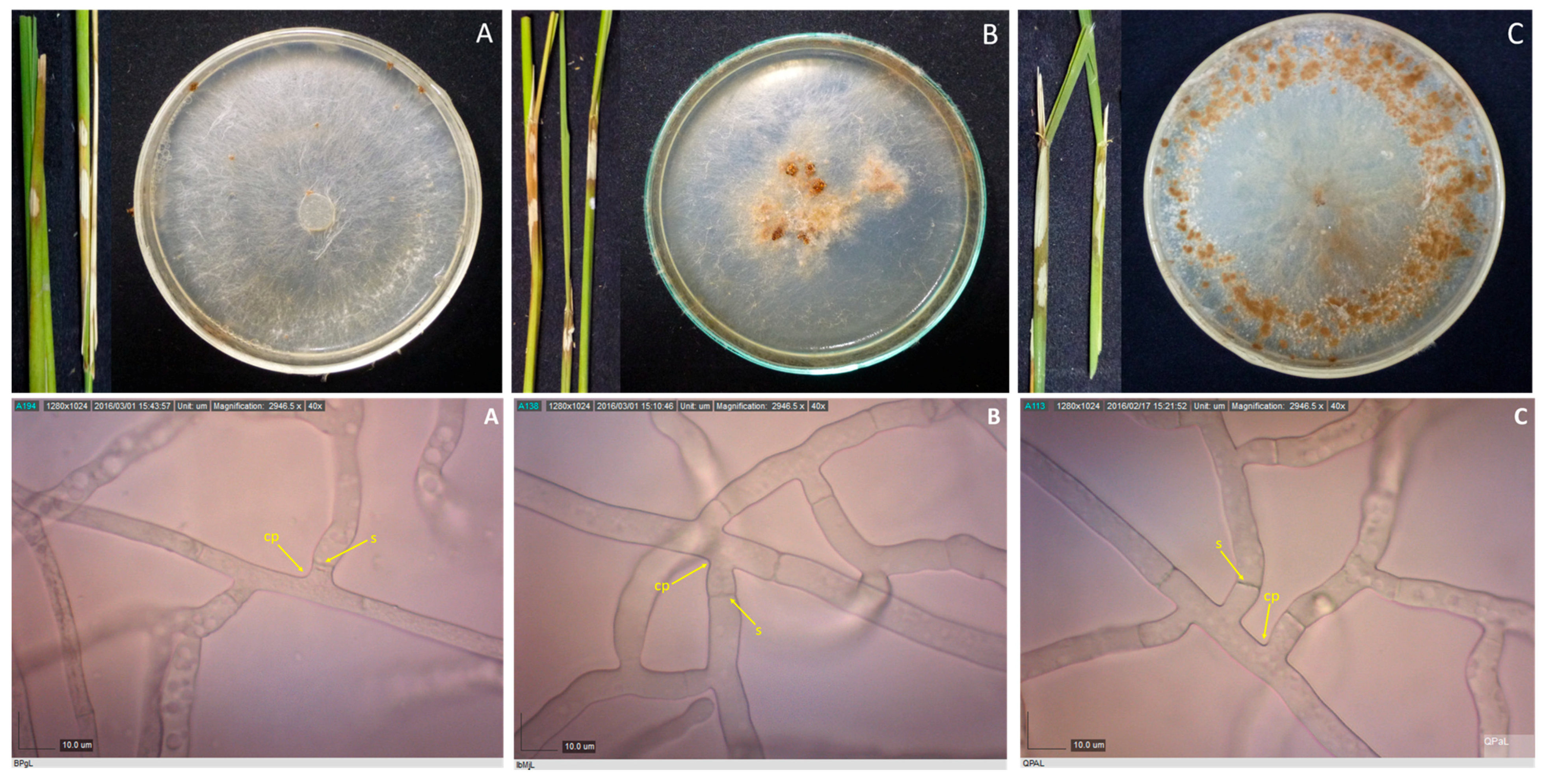
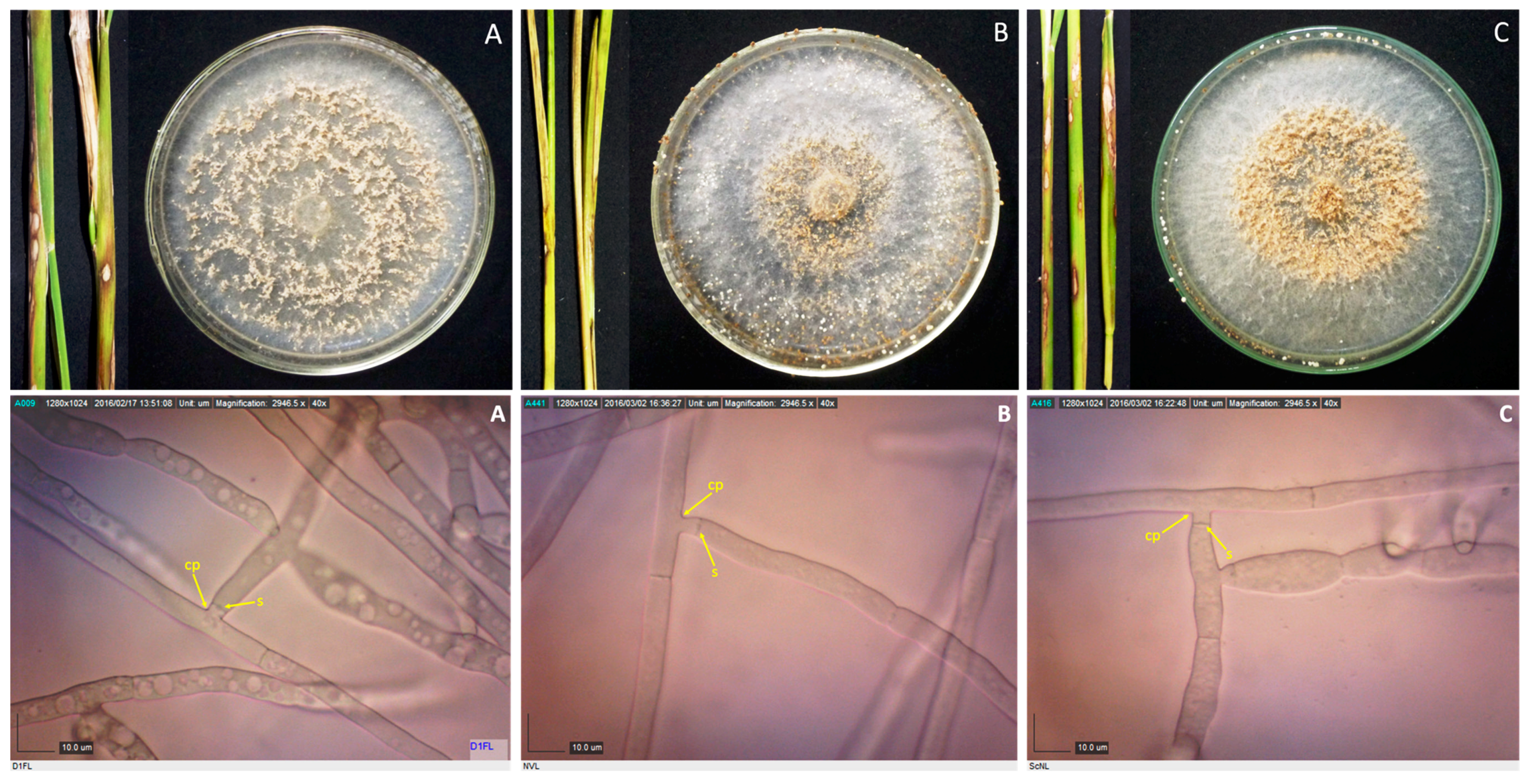
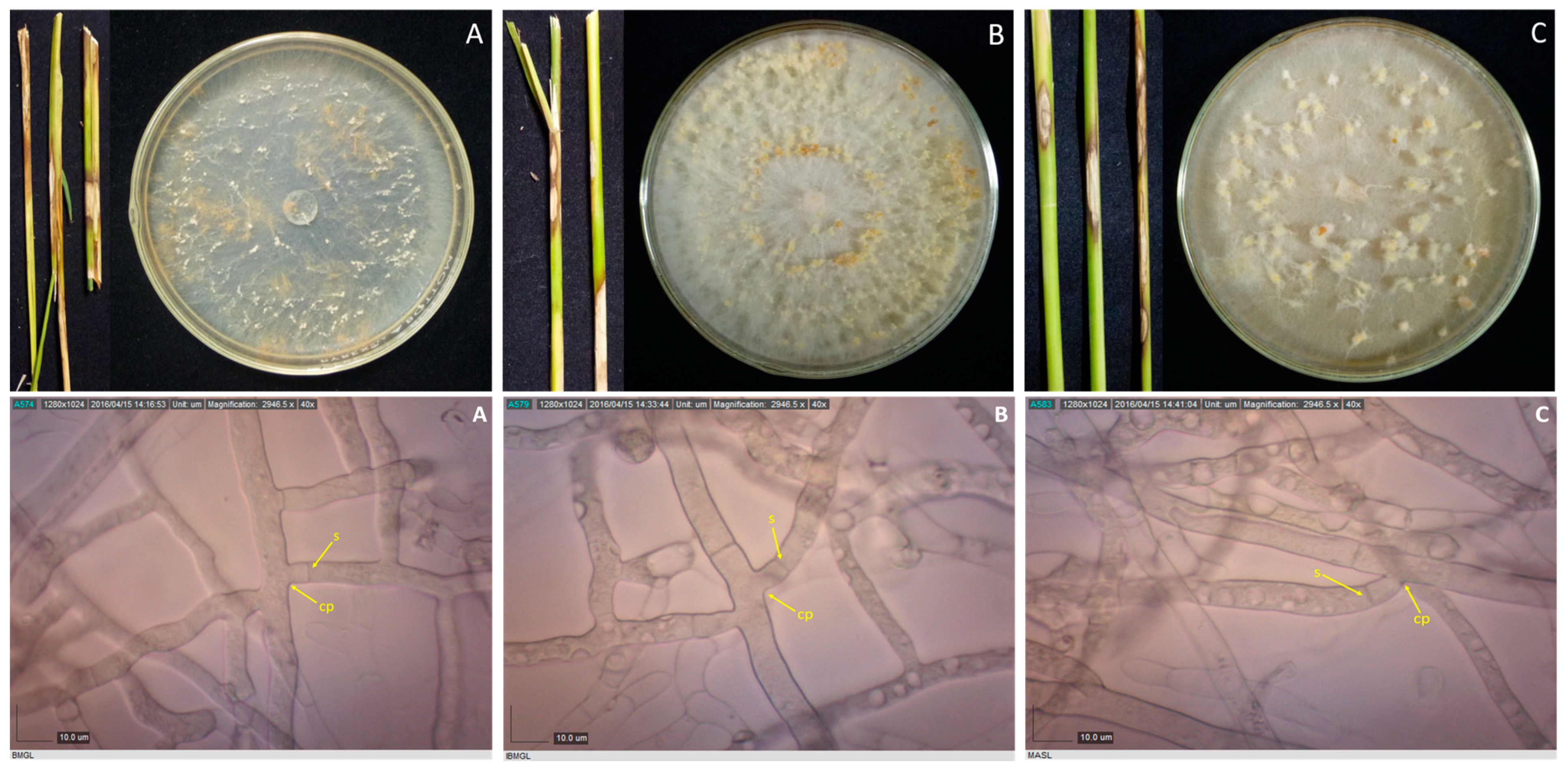
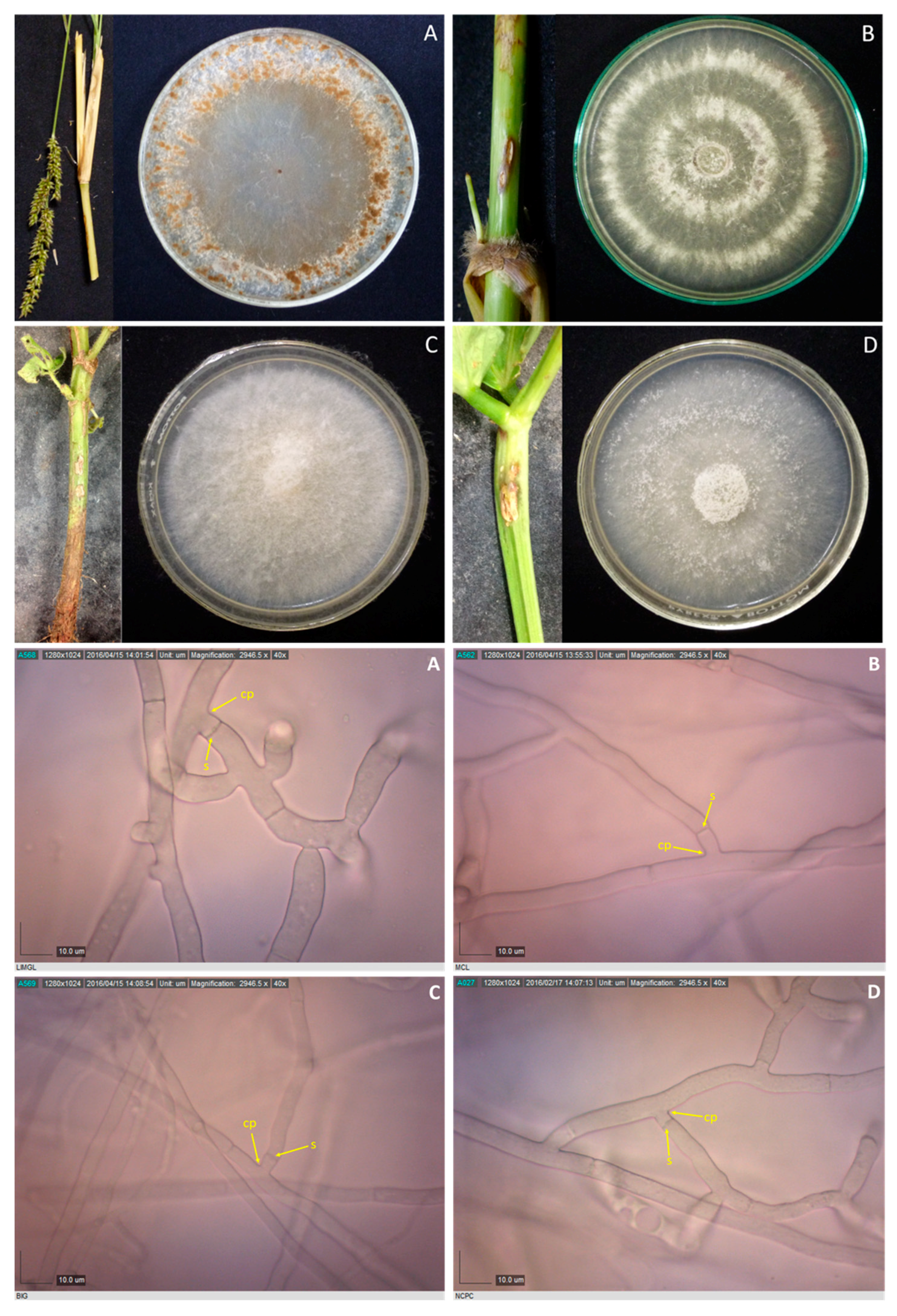
2.2. Culture Characterization
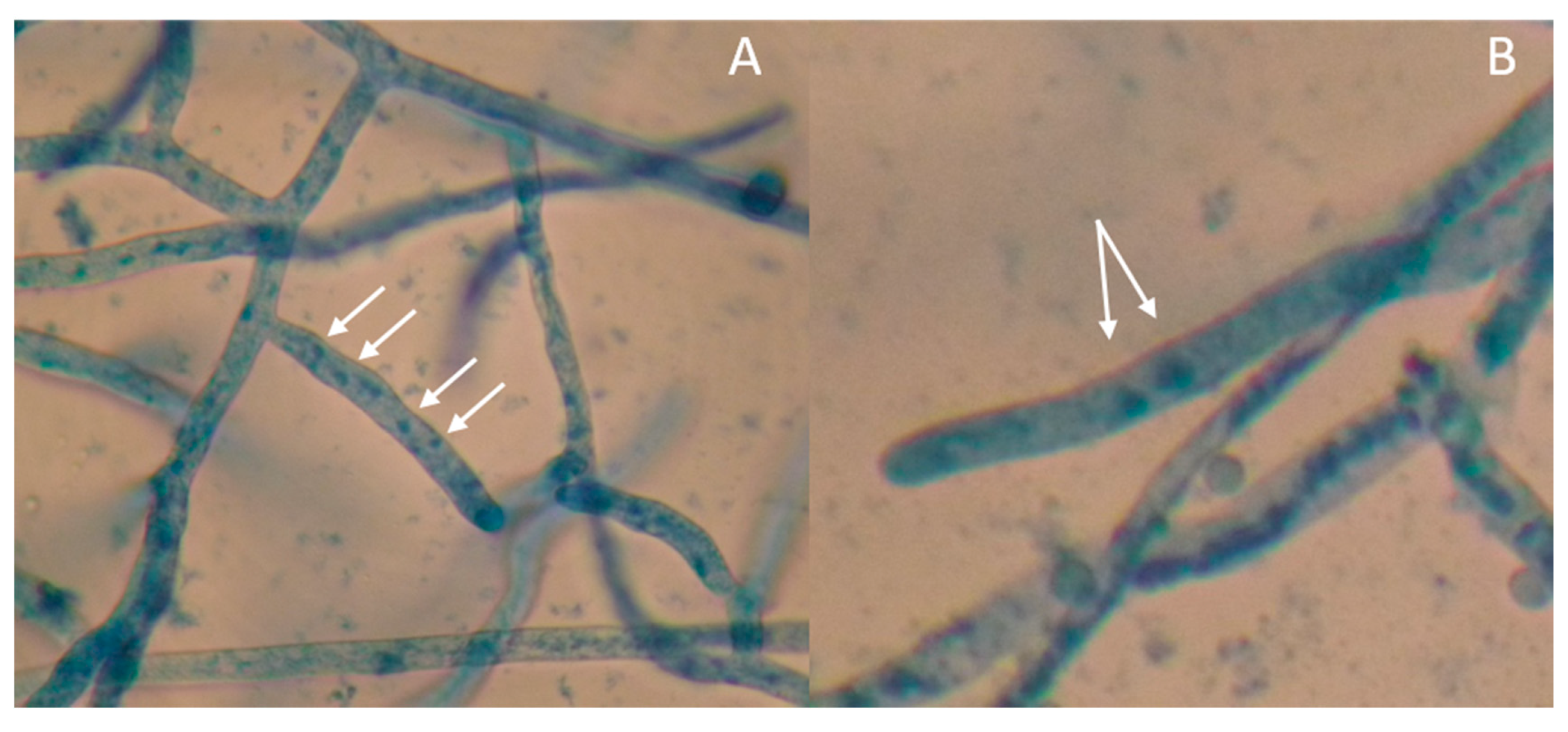
2.3. Microscopic Examination
2.4. DNA Extraction
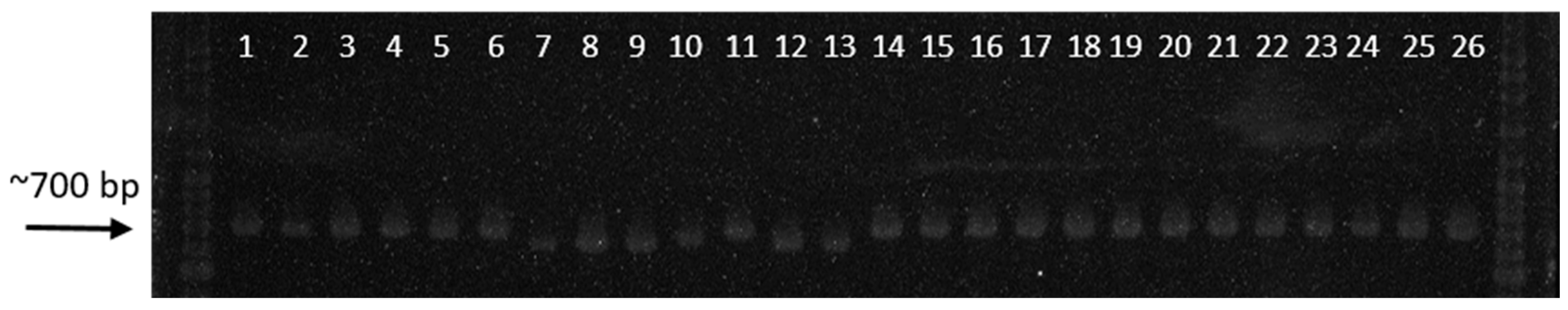
2.5. Detection of AG1-IA Isolates Using Specific Primers
2.6. Amplification of ITS 1–5.8S–ITS 2 Regions
2.7. Analysis of ITS Sequences
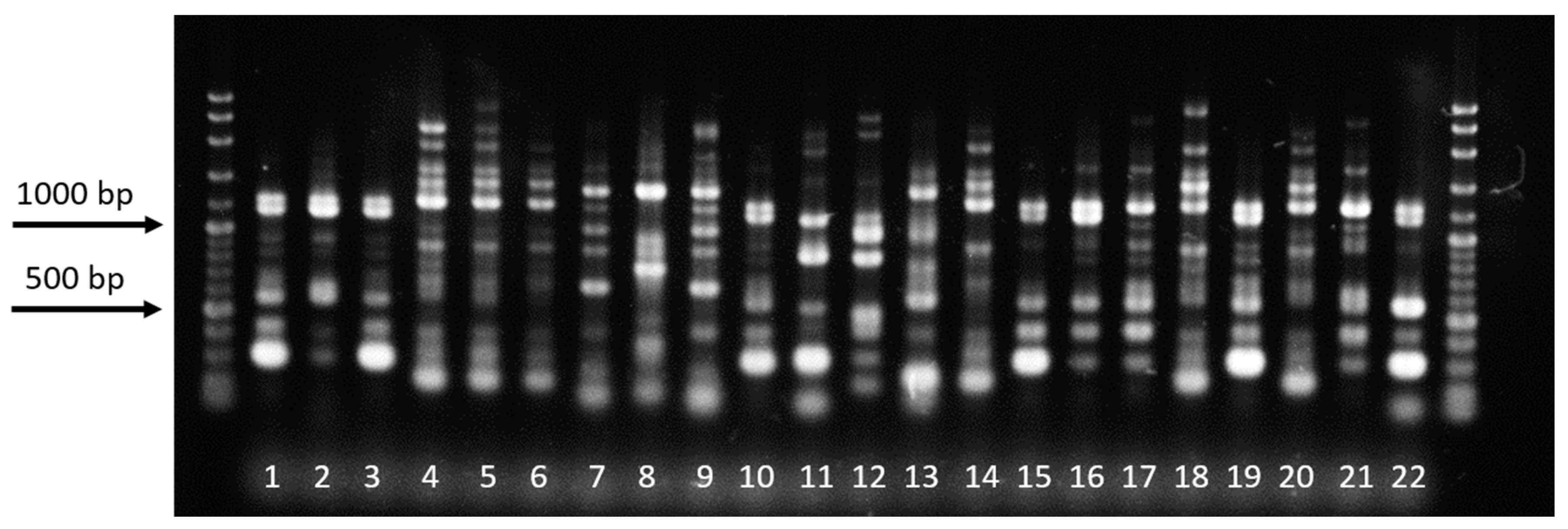
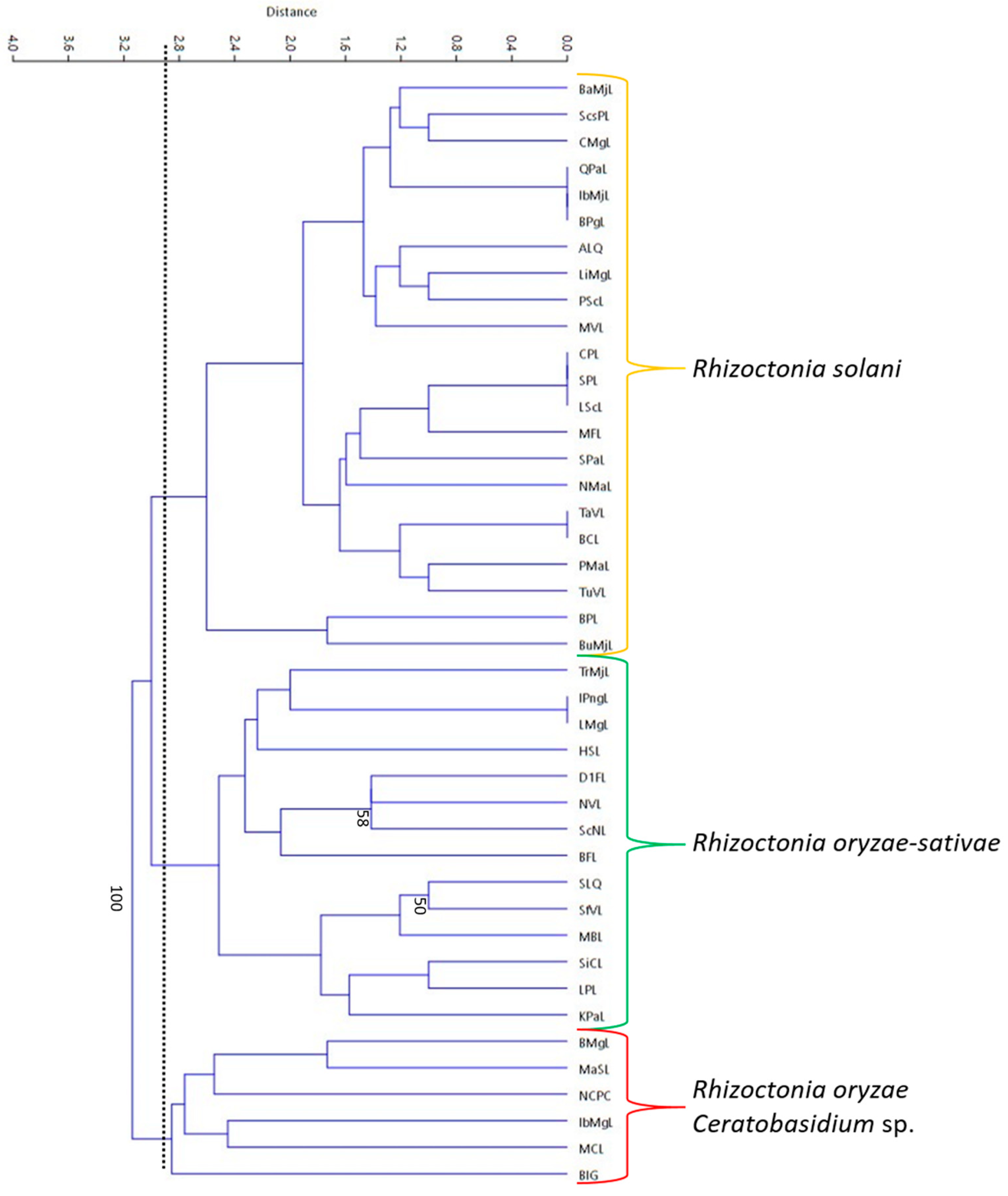
2.8. DNA Fingerprinting of Rhizoctonia Isolates Using Repetitive-Element PCR (Rep-PCR)
2.9. Pathogenicity Test
3. Results and Discussion
3.1. Collection and Isolation
3.2. Culture Characterization
3.3. Sclerotia Characterization
3.4. Direct Detection Using Specific Primers
3.5. Amplification of ITS 1–5.8S–ITS 2 Regions
3.6. Rep-PCR
3.7. Pathogenicity Test
Author Contributions
Funding
Conflicts of Interest
References
- Santiaguel, A.F. Nourishing a nation. Rice Today. Int. Rice Res. Inst. 2013, 12, 11–14. [Google Scholar]
- Philippines Statistics Authority (formerly Bureau of Agricultural Statistics). BAS 2015. Available online: http://countrystat.bas.gov.ph (accessed on 25 May 2015).
- Adhipathi, P.; Singh, V.; Meena, S.C. Virulence diversity of Rhizoctonia solani causing sheath blight disease in rice and its host pathogen interaction. Bioscan 2013, 8, 949–952. [Google Scholar]
- IRRI Rice Knowledge Bank. IRRI (2015) Sheath Blight. Available online: http://www.knowledgebank.irri.org/training/fact-sheets/pest-management/diseases/item/sheath-blight (accessed on 25 May 2015).
- Gianessi, L.P. Importance of Pesticides for Growing Rice in South and South East Asia. International Pesticide Benefit Case Study 108. Crop Life Foundation. Available online: https://croplife.org/case-study/importance-of-pesticides-for-growing-rice-in-south-and-south-east-asia/ (accessed on 18 March 2019).
- Dilla, E.M. Yield Loss Due to Sheath Blight in Direct Seeded Rice as Affected by Plant Density, Nitrogen Level, and Amount of Inoculum; University of the Philippines Los Baños: Los Baños, Philippines, Unpublished work; 1993. [Google Scholar]
- Lanoiselet, V.M.; Cother, E.J.; Ash, G.J. Aggregate sheath spot and sheath spot of rice. Crop Prot. 2007, 26, 799–808. [Google Scholar] [CrossRef]
- Inagaki, K.; Tamura, M.; Makino, M. Overwintering in plant residues of rice sclerotial disease fungi, Rhizoctonia and Sclerotium species. Proc. Kansai Plant Prot. Soc. 1997, 29, 27–29. [Google Scholar]
- Arakawa, M.; Inagaki, K. Molecular markers for genotyping anastomosis groups and understanding population biology of Rhizoctonia species. J. Gen. Plant Pathol. 2014, 80, 401–407. [Google Scholar] [CrossRef]
- Agrios, G.N. Plant Pathology, 5th ed.; Elsevier Academic Press: New York, NY, USA, 2005; p. 952. [Google Scholar]
- Nadarajah, K.; Omar, N.S.; Rosli, M.M.; Tze, O.S. Molecular Characterization and Screening for Sheath Blight Resistance Using Malaysian Isolates of Rhizoctonia solani. Biomed Res. Intern. 2014, 2014, 434257. [Google Scholar] [CrossRef]
- Aye, S.S.; Matsumoto, M. Genetic characterization by REP-PCR of Myanmar isolates of Rhizoctonia spp., causal agents of rice sheath diseases. J. Plant Pathol. 2010, 92, 255–260. [Google Scholar]
- Dubey, S.C.; Tripathi, A.; Upadhyay, B.K. Molecular diversity analysis of Rhizoctonia solani isolates infecting various pulse crops in different agro-ecological regions in India. Folia Microbiol. (Praha) 2012, 57, 513–524. [Google Scholar] [CrossRef]
- Khodayari, M.; Safaie, N.; Shamsbakhsh, M. Genetic Diversity of Iranian AG1-IA Isolates of Rhizoctonia solani, the cause of rice sheath blight, using morphological and molecular markers. J. Phytopathol. 2009, 157, 708–714. [Google Scholar] [CrossRef]
- Lal, M.; Singh, V.; Kandhari, J.; Sharma, P.; Kumar, V.; Murti, S. Diversity analysis of Rhizoctonia solani causing sheath blight of rice in India. Afr. J. Biotechnol. 2014, 13, 4594–4605. [Google Scholar]
- Toda, T.; Hyakumachi, M.; Arora, D.K. Genetic relatedness among and within different Rhizoctonia solani anastomosis groups as assessed by RAPD, ERIC, and REP-PCR. Microbiol. Res. 1999, 154, 247–258. [Google Scholar] [CrossRef]
- Goswami, S.K.; Singh, V.; Kashap, P.L. Population genetic structure of Rhizoctonia solani AG1IA from rice field in North India. Phytoparasitica 2017, 45, 299–316. [Google Scholar] [CrossRef]
- Moni, Z.R.; Ali, M.A.; Alam, M.S.; Rahman, M.S.; Bhuiyan, M.R.; Mian, M.S.; Iftekharuddaula, K.M.; Latif, M.A.; Khan, M.A.I. Morphological and genetical variability among Rhizoctonia solani isolates causing sheath blight disease of rice. Rice Sci. 2016, 23, 42–50. [Google Scholar] [CrossRef]
- Wang, L.; Liu, L.M.; Hou, Y.X.; Li, L.; Huang, S.W. Pathotypic and genetic diversity in the population of Rhizoctonia solani AG1-IA causing rice sheath blight in China. Plant Pathol. 2015, 64, 718–728. [Google Scholar] [CrossRef]
- Pascual, C.B.; Hyakumachi, M. Distribution of vegetatively compatible populations in a field planted with different host species. J. Gen. Plant Pathol. 2000, 66, 206–209. [Google Scholar] [CrossRef]
- Herr, L.J. Practical nuclear staining procedure for Rhizoctonia-like fungi. Phytopathology 1979, 69, 958–961. [Google Scholar] [CrossRef]
- Cenis, J.L. Rapid Extraction of Fungal DNA for PCR Amplification. Nucleic Acid. Res. 1992, 20, 2380. [Google Scholar] [CrossRef]
- Matsumoto, M. Trials for direct detection and identification of Rhizoctonia solani AG1 and AG 2 subgroups using specifically primed PCR analysis. Mycoscience 2002, 43, 185–189. [Google Scholar] [CrossRef]
- Johanson, A.; Turner, H.C.; McKay, G.J.; Brown, A.E. A PCR-based method to distinguish fungi of the sheath-blight complex, Rhizoctonia solani, R. oryzae and R. oryzae-sativae. FEMS Microbiol. Lett. 1998, 162, 289–294. [Google Scholar] [CrossRef]
- Versalovic, J.; Schneider, M.; De Bruijn, F. Genomic fingerprinting of bacteria using repetitive sequence-based polymerase chain reaction. Methods Mol. Cell Biol. 1994, 5, 25–40. [Google Scholar]
- Jia, Y.; Correa-Victoria, F.; Mcclung, A.; Zhu, L.; Liu, G.; Wamishe, Y.; Xie, J.; Marchetti, M.A.; Pinson, S.R.M.; Rutger, J.N.; Correll, J.C. Rapid determination of rice cultivar responses to the sheath blight pathogen Rhizoctonia solani using micro-chamber screening method. Plant Dis. 2007, 91, 485–489. [Google Scholar] [CrossRef]
- Castroagudin, V.L. Genetic Diversity, Pathogenicity/Virulence Variation and Population Structure of Rhizoctonia spp. Associated with Rice Sheath Blight in Arkansas and QTL Mapping for Disease Resistance; University of Arkansas: Fayetteville, NC, USA, Unpublished work; 2012. [Google Scholar]
- Duggar, B.M. Rhizoctonia crocorum (Pers.) DC and Rhizoctonia solani Kühn (Corticium vagum (B & C) with notes and other species. Ann. Mo. Bot. Gard. 1915, 2, 403–458. [Google Scholar]
- Butler, E.E.; Bracker, C.E. Morphology and cytology of Rhizoctonia solani. In Rhizoctonia solani, Biology and Pathology; Parmeter, J.R., Ed.; University of California Press: Los Angeles, CA, USA, 1970; pp. 32–51. [Google Scholar]
- Banniza, S.; Holderness, M. Rice sheath blight—pathogen biology and diversity. In Major Fungal Diseases of Rice; Sveenivasaprasad, S., Johnson, R., Eds.; Springer Science and Business Media: Dordrecht, The Netherlands, 2011; pp. 201–211. [Google Scholar]
- Gnanamanickam, S.S. Biological Control of Rice Diseases, Progress in Biological Control 8; Springer Science and Business Media: Dordrecht, The Netherlands, 2009. [Google Scholar]
- Shahjahan, A.K.M.; Mew, T.M. Sheath spot of rice in the Philippines. Int. Rice Res. Newsl. 1986, 11, 17. [Google Scholar]
- CABI Echinochloa crus-galli (Barnyard Grass) Data Sheet. Available online: http://www.cabi.org/isc/datasheet/20367 (accessed on 15 March 2016).
- Furoc, R.E.; Javier, E.Q. Integration of fodder production with intensive croppings involving rice. 1. Grass production from irrigated lowland rice field. 2. Herbage weeds during juvenile stage of the rice crop. Philippine J. Crop Sci. 1976, 1, 146–148. [Google Scholar]
- Singh, A.K.; Srivastava, J.N. Sheath blight disease of paddy and their management. In Recent Advances in the Diagnosis and Management of Plant Diseases; Awasthi, L.P., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; p. 294. [Google Scholar]
- Oplinger, E.S.; Hardman, L.L.; Kaminski, A.R.; Combs, S.M.; Doll, J.D. Mung Bean: Alternative Field Crops Manual. Available online: https://hort.purdue.edu/newcrop/afcm/mungbean.html (accessed on 18 March 2019).
- PCAARRD (2003) PCARRD Commodities: Mungbean. Available online: http://www.pcaarrd.dost.gov.ph/home/joomla/index.php?option=com_content&task=view&id=544&Itemid=418 (accessed on 18 April 2016).
- Butranu, W. Carrying capacity of component crops on Rhizoctonia solani Kuhn inoculum in relation to multiple cropping. Master’s Thesis, University of the Philippines Los Baños, Los Baños, Philippines, 1988. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Isolate | Host | Plant Part | Place of Collection | GPS Coordinates | Colony Color a | Growth Pattern b | Colony Growth Diameter (mm) c | Mean Diameter (mm) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | |||||||||
| 1 | ALQ | Rice | sheath | Ayuti, Lucban, Quezon | 14.10876° N 121.537007° E | light brown | abundant | 51.67 | 90.00 | 90.00 | 77.22 |
| 2 | SLQ | Rice | sheath | Samil, Lucban, Quezon | 14.109835° N 121.525667° E | light brown | moderate | 32.00 | 61.83 | 90.00 | 61.28 |
| 3 | BaMjL | Rice | sheath | Bakya, Majayjay | 14.16061° N 121.490699° E | light brown | moderate | 61.83 | 90.00 | 90.00 | 80.61 |
| 4 | BuMjL | Rice | sheath | Buharaw, Majayjay | 14.114552° N 121.50457° E | light brown | moderate | 50.33 | 90.00 | 90.00 | 76.78 |
| 5 | IbMjL | Rice | sheath | Ilayang Banga, Majayjay | 14.15° N 121.48° E | light brown | moderate | 61.17 | 74.33 | 74.67 | 70.06 |
| 6 | TrMjL | Rice | sheath | Talortor, Majayjay | 14.15° N 121.46° E | light brown | moderate | 33.33 | 69.33 | 90.00 | 64.22 |
| 7 | BMgL | Rice | sheath | Burlungan, Magdalena | 14.18434° N 121.442995° E | light salmon | scarce | 40.33 | 85.17 | 90.00 | 71.83 |
| 8 | CMgL | Rice | sheath | Cigapas, Magdalena | 14.22821° N 121.435905° E | light brown | moderate | 56.33 | 87.83 | 90.00 | 78.06 |
| 9 | IbMgL | Rice | sheath | Ibabang Bugtong, Magdalena | 10.178268° N −74.495031° W | yellowish brown | moderate | 54.67 | 90.00 | 90.00 | 78.22 |
| 10 | LiMgLg | Barnyard grass | sheath | Libunan, Magdalena | 10.178268° N −74.495031° W | light brown | abundant | 66.67 | 90.00 | 90.00 | 82.22 |
| 11 | LMgL | Rice | sheath | Libunan, Magdalena | 10.178268° N −74.495031° W | light brown | moderate | 35.17 | 66.33 | 90.00 | 63.83 |
| 12 | BPgL | Rice | sheath | Binan, Pagsanjan | 14.271785° N 121.436265° E | light brown | moderate | 43.33 | 77.67 | 89.17 | 70.06 |
| 13 | BPL | Rice | sheath | Buboy, Pagsanjan | 14.24° N 121.43° E | light brown | abundant | 60.33 | 90.00 | 90.00 | 80.11 |
| 14 | CPL | Rice | sheath | Cabanbanan, Pagsanjan | 14.24694° N 121.430908° E | light brown | moderate | 52.67 | 89.67 | 90.00 | 77.44 |
| 15 | SPL | Rice | sheath | Sabang, Pagsanjan | 14.26° N 121.43° E | brown | abundant | 55.83 | 83.33 | 89.17 | 76.11 |
| 16 | LScL | Rice | sheath | Labuin, Santa Cruz | 14.254683° N 121.395612° E | light brown | moderate | 61.83 | 90.00 | 90.00 | 80.61 |
| 17 | PScL | Rice | sheath | Patimbao, Santa Cruz | 14.266121° N 121.414189° E | light brown | moderate | 43.50 | 86.50 | 90.00 | 73.33 |
| 18 | LPL | Rice | sheath | Labuin, Pila | 14.248139° N 121.369887° E | light brown | moderate | 37.67 | 77.33 | 90.00 | 68.33 |
| 19 | ScsPL | Rice | sheath | Santa Clara Sur, Pila | 14.226484° N 121.368458° E | light brown | moderate | 60.00 | 90.00 | 90.00 | 80.00 |
| 20 | MVL | Rice | sheath | Masapang, Victoria | 14.192465° N 121.337007° E | light brown | moderate | 48.00 | 89.33 | 90.00 | 75.78 |
| 21 | NVL | Rice | sheath | Naninaya, Victoria | 14.192465° N 121.337007° E | light brown | moderate | 30.00 | 56.83 | 86.00 | 57.61 |
| 22 | SfVL | Rice | sheath | San Francisco, Victoria | 14.215199° N 121.339867° E | whitish brown | scarce | 33.83 | 67.00 | 90.00 | 63.61 |
| 23 | TaVL | Rice | sheath | Tangsa, Victoria | 14.202753° N 121.337007° E | light brown | moderate | 53.83 | 86.67 | 89.17 | 76.56 |
| 24 | TuVL | Rice | sheath | Tabuan, Victoria | 14.202753° N 121.337007° E | light brown | moderate | 50.67 | 90.00 | 90.00 | 76.89 |
| 25 | BCL | Rice | sheath | Bangyas, Calauan | 14.18482° N 121.309839° E | light brown | moderate | 49.33 | 86.67 | 90.00 | 75.33 |
| 26 | MCL | nknown grass | basal stem | Masiit, Calauan | 14.163578° N 121.305549° E | yellowish brown | moderate | 34.50 | 69.00 | 90.00 | 64.50 |
| 27 | SiCL | Rice | sheath | San Isidro, Calauan | 14.157863° N 121.318419° E | light brown | scarce | 35.83 | 68.5 | 90.00 | 64.78 |
| 28 | MBL | Rice | sheath | Maitim, Bae | 14.182059° N 121.275512° E | light brown | moderate | 36.00 | 69.83 | 90.00 | 65.28 |
| 29 | ScNL | Rice | sheath | Santa Clara, Nagcarlan | 14.149029° N 121.388467° E | light brown | moderate | 37.33 | 70.33 | 90.00 | 65.89 |
| 30 | KPaL | Rice | sheath | Kwatro, Paete | 14.367518° N 121.529954° E | light brown | moderate | 37.00 | 73.17 | 90.00 | 66.72 |
| 31 | QPaLr | Rice | sheath | Quinale, Paete | 14.360923° N 121.574104° E | light brown | moderate | 68.50 | 90.00 | 90.00 | 82.83 |
| 32 | SPaL | Rice | sheath | Syete, Paete | 14.367518° N 121.529954° E | light brown | moderate | 57.33 | 90.00 | 90.00 | 79.11 |
| 33 | IPngL | Rice | sheath | Isla, Pangil | 14.403307° N 121.469898° E | light brown | moderate | 32.50 | 66.17 | 90.00 | 62.89 |
| 34 | NMaL | Rice | sheath | Nanguma, Mabitac | 14.444753° N 121.422663° E | light brown | moderate | 64.50 | 90.00 | 90.00 | 81.50 |
| 35 | PMaL | Rice | sheath | Paagahan, Mabitac | 14.450122° N 121.405615° E | light brown | moderate | 60.00 | 90.00 | 90.00 | 80.00 |
| 36 | BFL | Rice | sheath | Batuhan, Famy | 14.436914° N 121.442761° E | light brown | moderate | 30.33 | 63.83 | 90.00 | 61.39 |
| 37 | D1FL | Rice | sheath | Dunghak 1, Famy | 14.473008° N 121.484245° E | light brown | scarce | 35.83 | 66.17 | 89.67 | 63.89 |
| 38 | MFL | Rice | sheath | Mayputat, Famy | 14.473008° N 121.484245° E | light brown | moderate | 59.83 | 90.00 | 90.00 | 79.94 |
| 39 | HSL | Rice | sheath | Halayhayin, Siniloan | 14.428391° N 121.465614° E | light brown | moderate | 33.67 | 63.67 | 90.00 | 62.44 |
| 40 | MaSL | Rice | sheath | Makatad, Siniloan | 14.436155° N 121.482751° E | yellowish brown | moderate | 54.83 | 89.33 | 90.00 | 78.06 |
| 41 | BIG | Mungbean | stem | Pili drive, UPLB | 14.163158° N 121.249706° E | yellowish brown | moderate | 35.17 | 65.50 | 90.00 | 63.56 |
| 42 | NCPC | Mungbean | stem | NCPC, UPLB | 14.167535° N 121.243282° E | yellowish brown | moderate | 29.50 | 55.67 | 80.33 | 55.17 |
| # | Isolate | Color of Sclerotia | Sclerotia Formation Pattern | Mean Sclerotia Diameter (mm) a | Sclerotial Production b | Mean Hyphal Width (µm) a | Nuclear Count | Binucleate (BNR)/ Multinucleate (MNR) | Molecular Identification |
|---|---|---|---|---|---|---|---|---|---|
| 1 | ALQ | brown/dark brown | scattered | 1.69 | fair | 7.57 | 5 | MNR | R. solani AG1-IA |
| 2 | SLQ | white/brown | scattered | 0.81 | excellent | 6.90 | 2 | BNR | ND |
| 3 | BaMjL | brown/dark brown | scattered | 2.48 | fair | 8.04 | 5 | MNR | R. solani AG1-IA |
| 4 | BuMjL | brown/dark brown | scattered | 1.66 | moderate | 8.07 | 9 | MNR | R. solani AG1-IA |
| 5 | IbMjL | brown/dark brown | central | 1.68 | fair | 7.85 | 8 | MNR | R. solani f. sp. sasakii |
| 6 | TrMjL | white/brown | scattered | 0.77 | excellent | ND | 2 | BNR | ND |
| 7 | BMgL | salmon | scattered | ND | excellent | 7.37 | 7 | MNR | R. oryzae |
| 8 | CMgL | brown/dark brown | scattered | 2.06 | good | 7.56 | 6 | MNR | R. solani AG1-IA |
| 9 | IbMgL | salmon | central | ND | excellent | 7.26 | 8 | MNR | R. oryzae |
| 10 | LiMgLg | brown/dark brown | peripheral | 1.76 | very good | 7.05 | 6 | MNR | R. solani AG1-IA |
| 11 | LMgL | white/brown | scattered | 0.63 | excellent | 7.16 | 2 | BNR | ND |
| 12 | BPgL | brown/dark brown | scattered | 1.48 | moderate | 7.48 | 9 | MNR | R. solani AG1-IA |
| 13 | BPL | brown/dark brown | scattered | 1.97 | moderate | 7.84 | 10 | MNR | R. solani AG1-IA |
| 14 | CPL | brown/dark brown | scattered | 2.69 | fair | 7.29 | 6 | MNR | R. solani AG1-IA |
| 15 | SPL | brown/dark brown | scattered | 1.83 | fair | ND | 6 | MNR | R. solani AG1-IA |
| 16 | LScL | NA | NA | NA | poor | 9.43 | 6 | MNR | R. solani AG1-IA |
| 17 | PScL | NA | NA | NA | poor | 7.69 | 7 | MNR | R. solani AG1-IA |
| 18 | LPL | white/brown | scattered | 0.86 | excellent | ND | 2 | BNR | ND |
| 19 | ScsPL | brown/dark brown | scattered | 1.68 | fair | 7.93 | 7 | MNR | R. solani AG1-IA |
| 20 | MVL | brown/dark brown | scattered | 1.89 | moderate | 6.12 | 7 | MNR | R. solani AG1-IA |
| 21 | NVL | white/brown | scattered | 0.81 | excellent | 6.33 | 2 | BNR | R. oryzae-sativae |
| 22 | SfVL | white/brown | central | 0.8 | excellent | 6.04 | 2 | BNR | ND |
| 23 | TaVL | NA | NA | NA | poor | 7.31 | 6 | MNR | R. solani AG1-IA |
| 24 | TuVL | brown/dark brown | peripheral | 2.92 | good | 7.93 | 6 | MNR | R. solani AG1-IA |
| 25 | BCL | brown/dark brown | scattered | 1.21 | moderate | 7.94 | 7 | MNR | R. solani AG1-IA |
| 26 | MCL | white to gray | ring | 1.39 | excellent | 5.07 | 8 | MNR | R. solani AG1-IA |
| 27 | SiCL | white/brown | scattered | 0.87 | excellent | 7.20 | 2 | BNR | ND |
| 28 | MBL | white/brown | central | 0.71 | excellent | 6.27 | 2 | BNR | ND |
| 29 | ScNL | white/brown | central | 0.54 | excellent | 6.28 | 2 | BNR | R. oryzae-sativae |
| 30 | KPaL | white/brown | scattered | 0.77 | excellent | 6.45 | 2 | BNR | ND |
| 31 | QPaL | brown/dark brown | peripheral | 2.52 | good | 6.96 | 7 | MNR | R. solani AG1-IA |
| 32 | SPaL | brown/dark brown | scattered | 1.52 | fair | 7.51 | 7 | MNR | R. solani AG1-IA |
| 33 | IPngL | white to brown | scattered | 0.78 | excellent | 5.82 | 2 | BNR | ND |
| 34 | NMaL | brown/dark brown | scattered | 2.54 | fair | 8.08 | 8 | MNR | R. solani AG1-IA |
| 35 | PMaL | brown/dark brown | scattered | 2.05 | fair | 7.35 | 6 | MNR | R. solani AG1-IA |
| 36 | BFL | white/brown | scattered | 0.64 | excellent | ND | 2 | BNR | ND |
| 37 | D1FL | white/brown | scattered | 0.61 | excellent | 6.09 | 2 | BNR | R. oryzae-sativae |
| 38 | MFL | brown/dark brown | peripheral | 1.91 | very good | 7.28 | 6 | MNR | R. solani AG1-IA |
| 39 | HSL | white/brown | scattered | 0.84 | excellent | ND | 2 | BNR | ND |
| 40 | MaSL | salmon | ring | ND | excellent | 8.62 | 10 | MNR | R. oryzae |
| 41 | BIG | NA | NA | NA | poor | 5.02 | 2 | BNR | Ceratobasidium sp. AG-L |
| 42 | NCPC | NA | NA | NA | poor | 4.60 | 2 | BNR | Ceratobasidium sp. AG-A |
| Isolate | Percent Similarity (%) | Species Identification | Accession No of Standard Isolates | Country of Origin |
|---|---|---|---|---|
| IbMjL | 99 | Rhizoctonia solani f. sp. sasakii | KF570312.1 | India |
| D1FL | 100 | Ceratorhiza oryzae-sativae | DQ307249.1 | China |
| NVL | 97 | Ceratorhiza oryzae-sativae | FJ667260.1 | Japan |
| ScNL | 100 | Ceratorhiza oryzae-sativae | DQ307249.1 | China |
| LiMgLg | 97 | Rhizoctonia solani | JF701789.1 | India |
| MCLgb | 98 | Ceratobasidium sp. AG-Ba | KF176604.1 | China |
| NCPC | 99 | Ceratobasidium sp. AG-L | FJ515884.1 | China |
| BIG | 99 | Ceratobasidium sp. AG-A | KC782943.1 | Italy |
| BMgL | 100 | Waitea circinata | EF429315.1 | China |
| IbMgL | 90 | Waitea circinata | HM597138.1 | USA |
| MaSL | 100 | Waitea circinata | EF429315.1 | China |
| Isolate | Rice | Barnyard Grass | Tomato | Mungbean |
|---|---|---|---|---|
| BPgL | 41.85 b | 58.78 a | 73.00 d | 24.08 a |
| IbMjL | 40.54 b | 47.23 a,b | 62.75 d | 25.42 a |
| QPaL | 40.48 b | 58.09 a | 62.75 d | 23.00 a |
| D1FL | 16.72 c | 1.66 b | 20.50 a | 0 |
| NVL | 20.45 c | 0.27 b | 20.50 a | 0 |
| ScNL | 15.36 c | 1.24 b | 20.50 a | 0 |
| BMgL | 32.47 b | 18.68 a,b | 26.00 a,b | 0 |
| IbMgL | 39.69 b | 25.168 a,b | 48.00 b,c,d | 0 |
| MaSL | 41.24 b | 36.44 a,b | 31.50 a,b,c | 0 |
| LiMgL | 51.42 a | 49.40 a,b | 20.50 a | 25.50 a |
| MCL | ND | 1.36 b | 37.00 a,b,c | 0 |
| NCPC | ND | 3.44 b | 37.00 a,b,c | ND |
| BIG | ND | ND | 53.50 c,d | ND |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sandoval, R.F.C.; Cumagun, C.J.R. Phenotypic and Molecular Analyses of Rhizoctonia spp. Associated with Rice and Other Hosts. Microorganisms 2019, 7, 88. https://doi.org/10.3390/microorganisms7030088
Sandoval RFC, Cumagun CJR. Phenotypic and Molecular Analyses of Rhizoctonia spp. Associated with Rice and Other Hosts. Microorganisms. 2019; 7(3):88. https://doi.org/10.3390/microorganisms7030088
Chicago/Turabian StyleSandoval, Regina Faye C., and Christian Joseph R. Cumagun. 2019. "Phenotypic and Molecular Analyses of Rhizoctonia spp. Associated with Rice and Other Hosts" Microorganisms 7, no. 3: 88. https://doi.org/10.3390/microorganisms7030088
APA StyleSandoval, R. F. C., & Cumagun, C. J. R. (2019). Phenotypic and Molecular Analyses of Rhizoctonia spp. Associated with Rice and Other Hosts. Microorganisms, 7(3), 88. https://doi.org/10.3390/microorganisms7030088