Oral Administration of a Select Mixture of Lactobacillus and Bacillus Alleviates Inflammation and Maintains Mucosal Barrier Integrity in the Ileum of Pigs Challenged with Salmonella Infantis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Characterization of B. licheniformis BL1721 and B. subtilis BS1715 In Vitro
2.3. Ethics Statement
2.4. Animals
2.5. Animal Experiments
2.6. Clinical Examinations and Microbiological Analyses
2.7. Differential Blood Leukocyte Count
2.8. Histopathologic Scoring
2.9. Alcian Blue/Periodic Acid–Schiff Staining and Subsequent Quantification
2.10. Analysis of DNA Fragmentation
2.11. Western Blotting
2.12. Statistical Analysis
3. Results
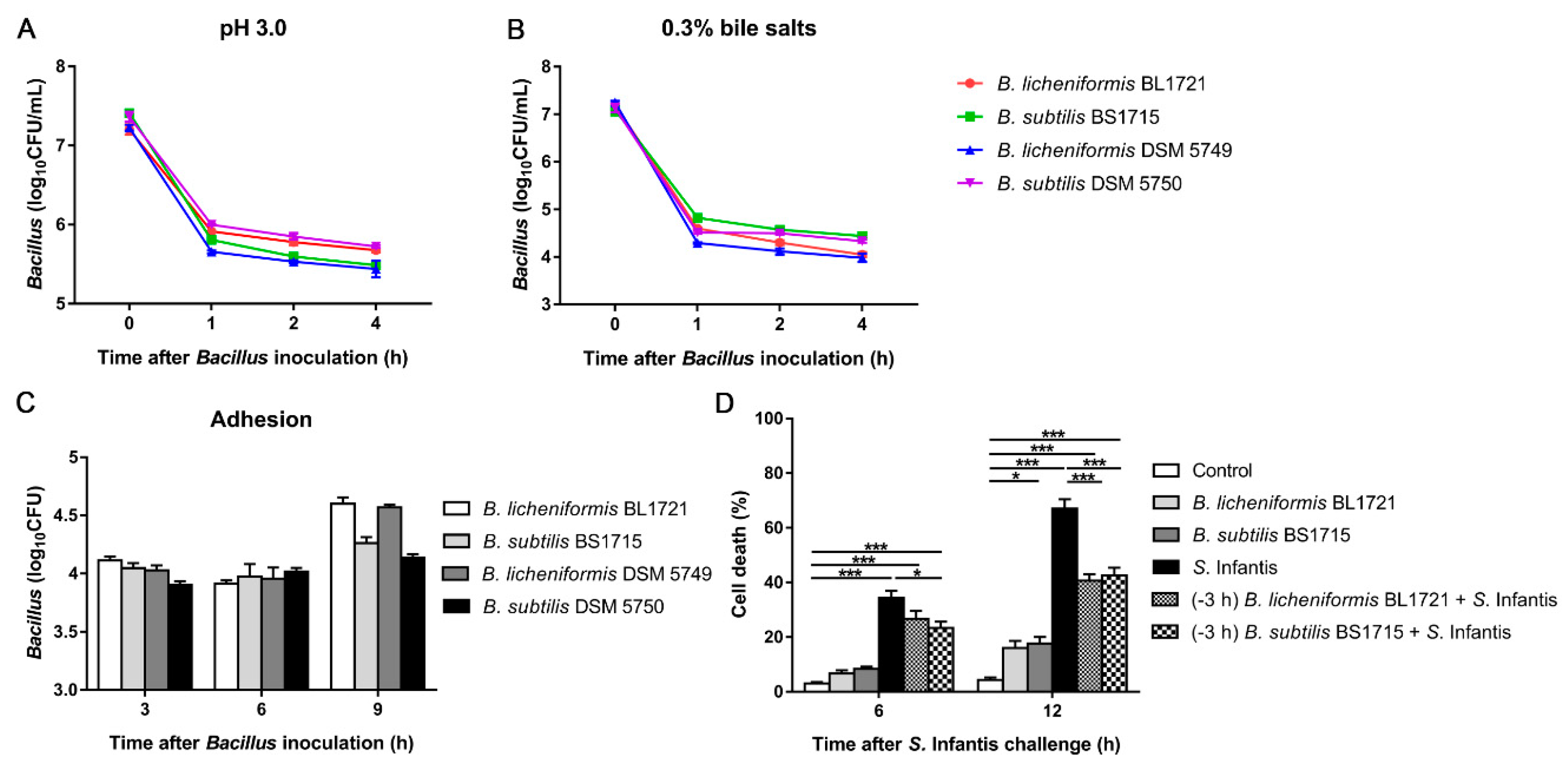
3.1. In Vitro Probiotic Characteristics of B. licheniformis BL1721 and B. subtilis BS1715
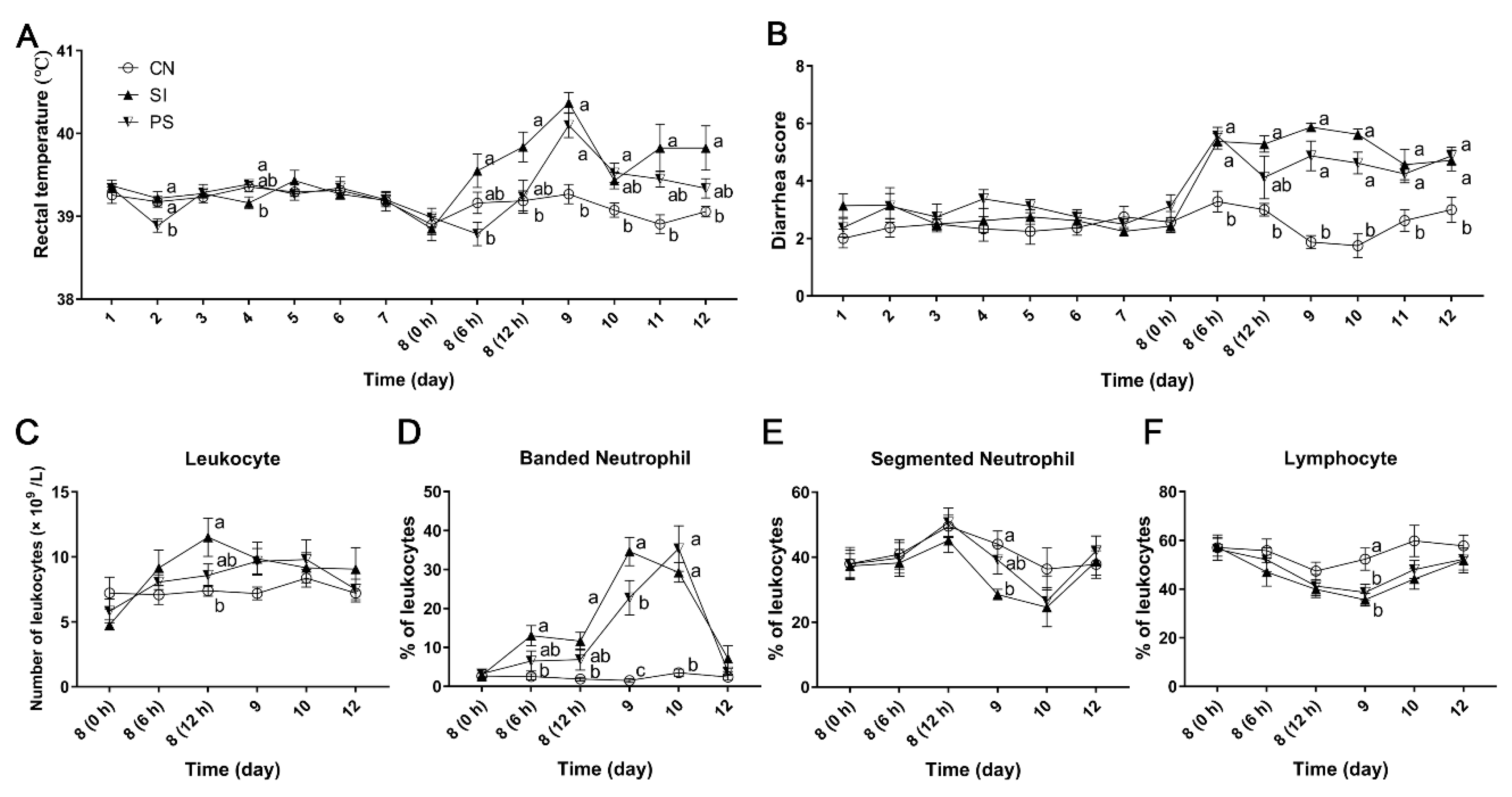
3.2. Clinical Symptoms and Growth Performance
3.3. The Effects of Oral Administration of LBB-Mix on Blood Leukocyte Count and Population Distribution
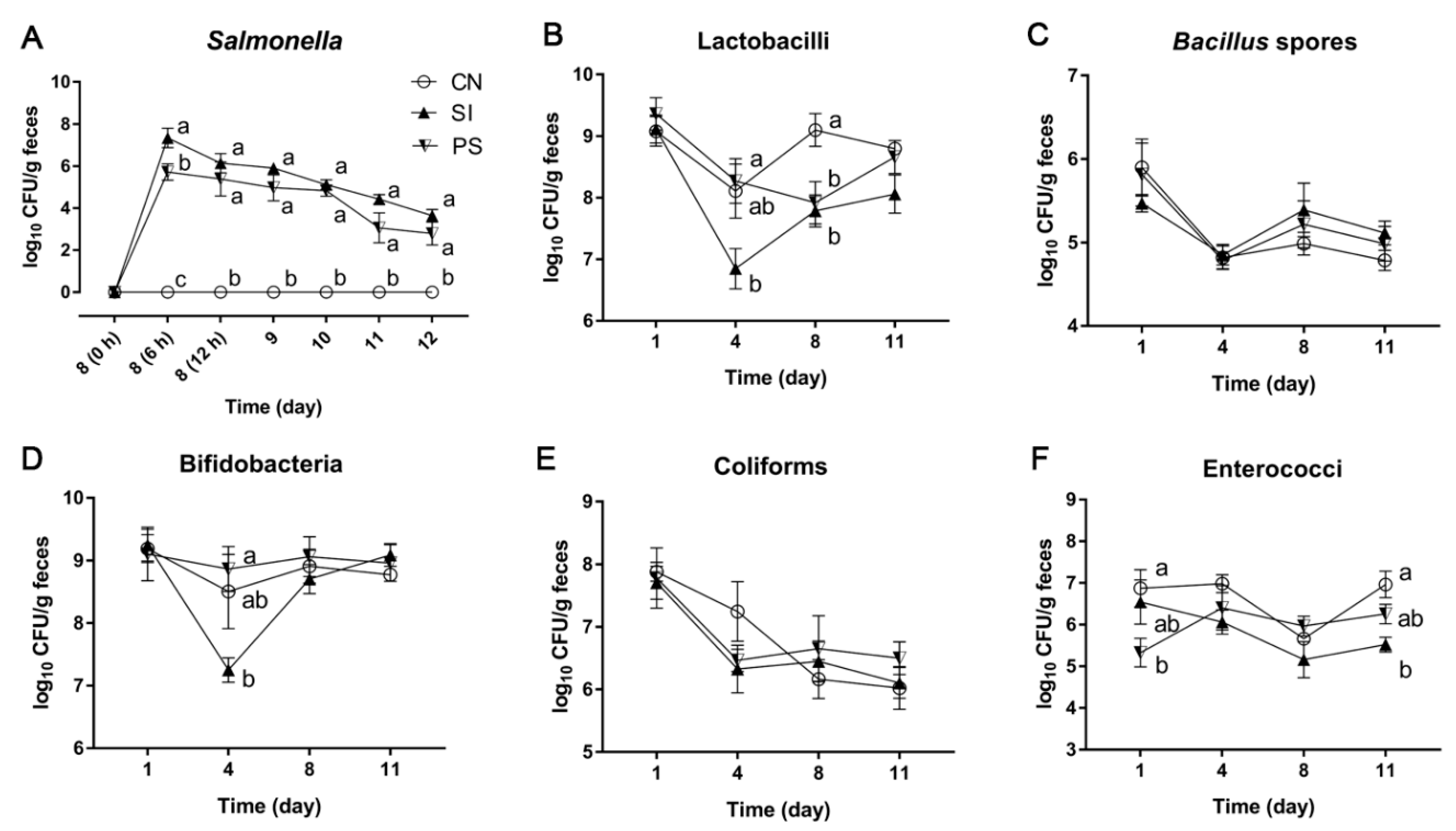
3.4. The LBB-Mix Changes the Composition of Fecal Microbiota during Salmonella Infection
3.5. Oral Administration of LBB-Mix Attenuated the Severity of Intestinal Damage and Inflammation Induced by S. Infantis
3.6. Orally Fed LBB-Mix Increased Goblet Cell Count and Claudin 1 Protein Expression during S. Infantis Infection
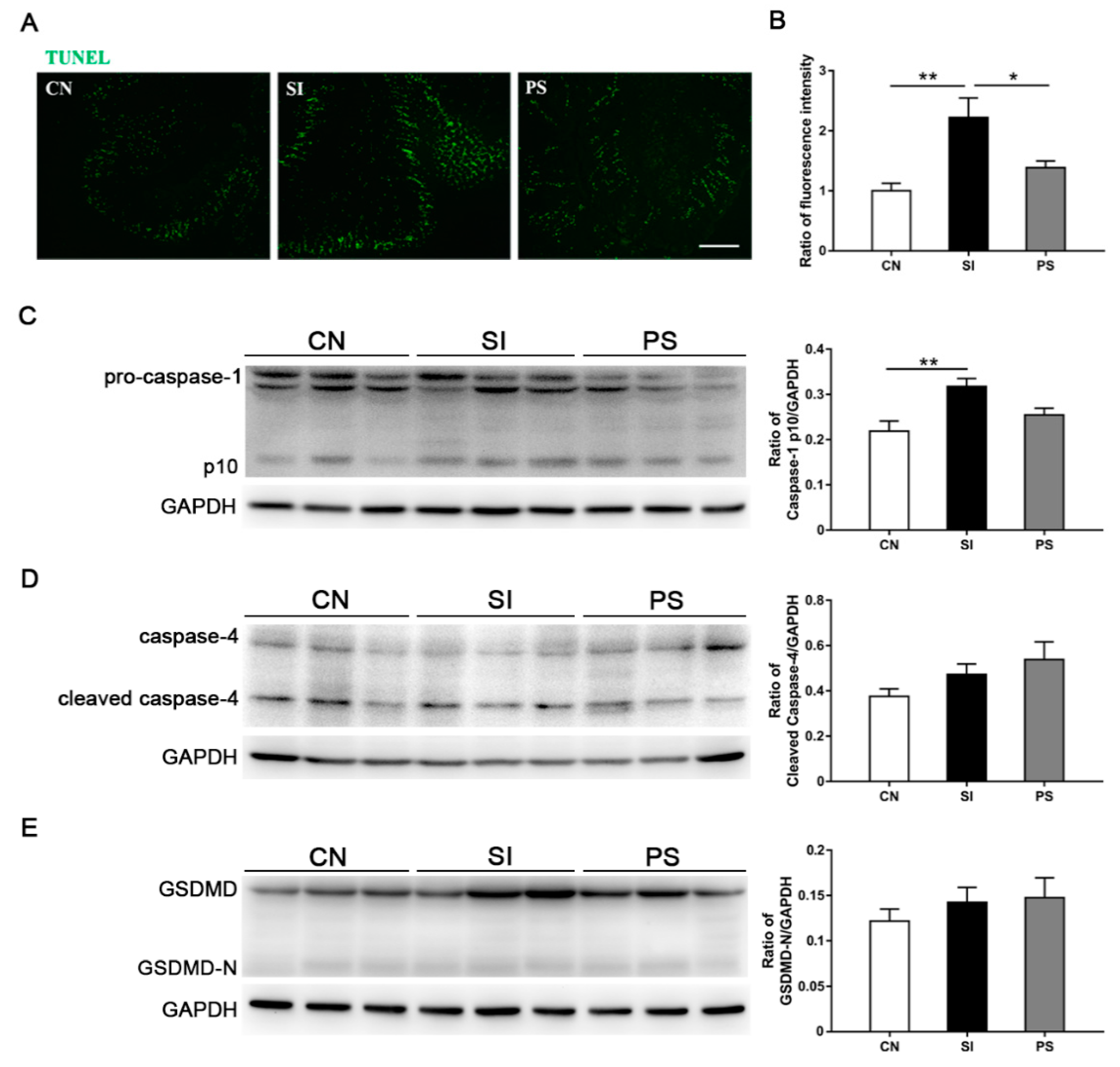
3.7. Orally Fed LBB-Mix Reduces Cell Death in the Intestine after S. Infantis Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- European Food Safety Authority and European Centre for Disease Prevention and Control (EFSA and ECDC). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J. 2018, 16, e05500. [Google Scholar]
- Schroeder, S.; Harries, M.; Prager, R.; Höfig, A.; Ahrens, B.; Hoffmann, L.; Rabsch, W.; Mertens, E.; Rimek, D. A prolonged outbreak of Salmonella Infantis associated with pork products in central Germany, April–October 2013. Epidemiol. Infect. 2016, 144, 1429–1439. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.; Krull, A.; Wang, C.; Erdman, M.; Fedorka-Cray, P.; Logue, C.; O’Connor, A.M. Changes in the prevalence of Salmonella serovars associated swine production and correlations of avian, bovine and swine-associated serovars with human-associated serovars in the United States (1997–2015). Zoonoses Public Health 2018, 65, 648–661. [Google Scholar] [CrossRef] [PubMed]
- Su, J.-H.; Zhu, Y.-H.; Ren, T.-Y.; Guo, L.; Yang, G.-Y.; Jiao, L.-G.; Wang, J.-F. Distribution and Antimicrobial Resistance of Salmonella Isolated from Pigs with Diarrhea in China. Microorganisms 2018, 6, 117. [Google Scholar] [CrossRef] [PubMed]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilhelm, B.J.; Young, I.; Cahill, S.; Nakagawa, R.; Desmarchelier, P.; Rajić, A. Rapid systematic review and meta-analysis of the evidence for effectiveness of primary production interventions to control Salmonella in beef and pork. Prev. Vet. Med. 2017, 147, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Chapman, C.; Gibson, G.R.; Rowland, I. Health benefits of probiotics: Are mixtures more effective than single strains? Eur. J. Nutr. 2011, 50, 1–17. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Zhu, Y.-H.; Yu, J.; Xia, B.; Liu, X.; Yang, G.-Y.; Su, J.-H.; Guo, L.; Wang, M.-L.; Wang, J.-F. Lactobacillus johnsonii L531 reduces pathogen load and helps maintain short-chain fatty acid levels in the intestines of pigs challenged with Salmonella enterica Infantis. Vet. Microbiol. 2019, 230, 187–194. [Google Scholar] [CrossRef]
- Yang, G.-Y.; Zhu, Y.-H.; Zhang, W.; Zhou, D.; Zhai, C.-C.; Wang, J.-F. Influence of orally fed a select mixture of Bacillus probiotics on intestinal T-cell migration in weaned MUC4 resistant pigs following Escherichia coli challenge. Vet. Res. 2016, 47, 71. [Google Scholar] [CrossRef]
- Zhang, W.; Zhu, Y.-H.; Zhou, D.; Wu, Q.; Song, D.; Dicksved, J.; Wang, J.-F. Oral administration of a select mixture of Bacillus probiotics affects the gut microbiota and goblet cell function following Escherichia coli challenge in newly weaned pigs of genotype MUC4 that are supposed to be enterotoxigenic E. coli F4ab/ac receptor negative. Appl. Environ. Microbiol. 2017, 83, e02747-16. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.; Davis, E.; Spencer, J.; Moser, R.; Rehberger, T. The effect of a Bacillus-based direct-fed microbial supplemented to sows on the gastrointestinal microbiota of their neonatal piglets. J. Anim. Sci. 2013, 91, 3390–3399. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Ye, L.; Lu, X.; Xie, S.; Yang, Q.; Yu, Q. Lactobacillus acidophilus Alleviated Salmonella-Induced Goblet Cells Loss and Colitis by Notch Pathway. Mol. Nutr. Food Res. 2018, 62, 1800552. [Google Scholar] [CrossRef] [PubMed]
- Groschwitz, K.R.; Hogan, S.P. Intestinal barrier function: Molecular regulation and disease pathogenesis. J. Allergy Clin. Immunol. 2009, 124, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Landy, J.; Ronde, E.; English, N.; Clark, S.K.; Hart, A.L.; Knight, S.C.; Ciclitira, P.J.; Al-Hassi, H.O. Tight junctions in inflammatory bowel diseases and inflammatory bowel disease associated colorectal cancer. World J. Gastroenterol. 2016, 22, 3117. [Google Scholar] [CrossRef]
- Lépine, A.F.; de Wit, N.; Oosterink, E.; Wichers, H.; Mes, J.; de Vos, P. Lactobacillus acidophilus attenuates Salmonella-induced stress of epithelial cells by modulating tight-junction genes and cytokine responses. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Rhayat, L.; Maresca, M.; Nicoletti, C.; Perrier, J.; Brinch, K.S.; Christian, S.; Devillard, E.; Eckhardt, E. Effect of Bacillus subtilis strains on intestinal barrier function and inflammatory response. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Shalini, S.; Dorstyn, L.; Dawar, S.; Kumar, S. Old, new and emerging functions of caspases. Cell Death Differ. 2015, 22, 526. [Google Scholar] [CrossRef]
- Tapia, V.S.; Daniels, M.J.; Palazón-Riquelme, P.; Dewhurst, M.; Luheshi, N.M.; Rivers-Auty, J.; Green, J.; Redondo-Castro, E.; Kaldis, P.; Lopez-Castejon, G. The three cytokines IL-1β, IL-18, and IL-1α share related but distinct secretory routes. J. Biol. Chem. 2019. [Google Scholar] [CrossRef]
- Shi, J.; Zhao, Y.; Wang, Y.; Gao, W.; Ding, J.; Li, P.; Hu, L.; Shao, F. Inflammatory caspases are innate immune receptors for intracellular LPS. Nature 2014, 514, 187. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zhao, Y.; Wang, K.; Shi, X.; Wang, Y.; Huang, H.; Zhuang, Y.; Cai, T.; Wang, F.; Shao, F. Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature 2015, 526, 660. [Google Scholar] [CrossRef]
- Kayagaki, N.; Stowe, I.B.; Lee, B.L.; O’Rourke, K.; Anderson, K.; Warming, S.; Cuellar, T.; Haley, B.; Roose-Girma, M.; Phung, Q.T. Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling. Nature 2015, 526, 666. [Google Scholar] [CrossRef]
- He, W.-T.; Wan, H.; Hu, L.; Chen, P.; Wang, X.; Huang, Z.; Yang, Z.-H.; Zhong, C.-Q.; Han, J. Gasdermin D is an executor of pyroptosis and required for interleukin-1β secretion. Cell Res. 2015, 25, 1285. [Google Scholar] [CrossRef]
- Wu, Q.; Zhu, Y.-H.; Xu, J.; Liu, X.; Duan, C.; Wang, M.-J.; Wang, J.-F. Lactobacillus rhamnosus GR-1 Ameliorates Escherichia coli-induced Activation of NLRP3 and NLRC4 Inflammasomes with Differential Requirement for ASC. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.-Y.; Yu, J.; Su, J.-H.; Jiao, L.-G.; Liu, X.; Zhu, Y.-H. Oral administration of Lactobacillus rhamnosus GG ameliorates Salmonella Infantis-induced inflammation in a pig model via activation of the IL-22BP/IL-22/STAT3 pathway. Front. Cell. Infect. Microbiol. 2017, 7, 323. [Google Scholar] [CrossRef] [PubMed]
- Birchenough, G.M.; Johansson, M.E.; Gustafsson, J.K.; Bergström, J.H.; Hansson, G.C. New developments in goblet cell mucus secretion and function. Mucosal Immunol. 2015, 8, 712. [Google Scholar] [CrossRef] [PubMed]
- Barthel, M.; Hapfelmeier, S.; Quintanilla-Martínez, L.; Kremer, M.; Rohde, M.; Hogardt, M.; Pfeffer, K.; Rüssmann, H.; Hardt, W.-D. Pretreatment of mice with streptomycin provides a Salmonella enterica serovar Typhimurium colitis model that allows analysis of both pathogen and host. Infect. Immunity 2003, 71, 2839–2858. [Google Scholar] [CrossRef]
- Yang, F.; Wang, A.; Zeng, X.; Hou, C.; Liu, H.; Qiao, S. Lactobacillus reuteri I5007 modulates tight junction protein expression in IPEC-J2 cells with LPS stimulation and in newborn piglets under normal conditions. BMC Microbiol. 2015, 15, 32. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Xiao, K.; Ke, Y.; Jiao, L.; Hu, C.; Diao, Q.; Shi, B.; Zou, X. Effect of a probiotic mixture on intestinal microflora, morphology, and barrier integrity of broilers subjected to heat stress. Poult. Sci. 2014, 93, 581–588. [Google Scholar] [CrossRef] [Green Version]
- Cummins, P.M. Occludin: One protein, many forms. Mol. Cell. Biol. 2012, 32, 242–250. [Google Scholar] [CrossRef]
- Martins, R.P.; Aguilar, C.; Graham, J.E.; Carvajal, A.; Bautista, R.; Claros, M.G.; Garrido, J.J. Pyroptosis and adaptive immunity mechanisms are promptly engendered in mesenteric lymph-nodes during pig infections with Salmonella enterica serovar Typhimurium. Vet. Res. 2013, 44, 120. [Google Scholar] [CrossRef] [PubMed]
- Mariathasan, S.; Newton, K.; Monack, D.M.; Vucic, D.; French, D.M.; Lee, W.P.; Roose-Girma, M.; Erickson, S.; Dixit, V.M. Differential activation of the inflammasome by caspase-1 adaptors ASC and IPAF. Nature 2004, 430, 213. [Google Scholar] [CrossRef] [PubMed]
- Broz, P.; Newton, K.; Lamkanfi, M.; Mariathasan, S.; Dixit, V.M.; Monack, D.M. Redundant roles for inflammasome receptors NLRP3 and NLRC4 in host defense against Salmonella. J. Exp. Med. 2010, 207, 1745–1755. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, L.; Kling, D.; Lorca, G.; Gonzalez, C. Lactobacillus johnsonii N6. 2 diminishes caspase-1 maturation in the gastrointestinal system of diabetes prone rats. Benef. Microbes 2018, 9, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Knodler, L.A.; Crowley, S.M.; Sham, H.P.; Yang, H.; Wrande, M.; Ma, C.; Ernst, R.K.; Steele-Mortimer, O.; Celli, J.; Vallance, B.A. Noncanonical inflammasome activation of caspase-4/caspase-11 mediates epithelial defenses against enteric bacterial pathogens. Cell Host Microbe 2014, 16, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Baker, P.J.; Boucher, D.; Bierschenk, D.; Tebartz, C.; Whitney, P.G.; D’Silva, D.B.; Tanzer, M.C.; Monteleone, M.; Robertson, A.A.; Cooper, M.A. NLRP3 inflammasome activation downstream of cytoplasmic LPS recognition by both caspase-4 and caspase-5. Eur. J. Immunol. 2015, 45, 2918–2926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, E.A.; Mao, D.P.; Yudkovsky, N.; Bonneau, R.; Lorang, C.G.; Warren, S.E.; Leaf, I.A.; Aderem, A. Innate immune detection of the type III secretion apparatus through the NLRC4 inflammasome. Proc. Natl. Acad. Sci. USA 2010, 107, 3076–3080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, E.A.; Leaf, I.A.; Treuting, P.M.; Mao, D.P.; Dors, M.; Sarkar, A.; Warren, S.E.; Wewers, M.D.; Aderem, A. Caspase-1-induced pyroptosis is an innate immune effector mechanism against intracellular bacteria. Nat. Immunol. 2010, 11, 1136. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Wang, K.; Liu, W.; She, Y.; Sun, Q.; Shi, J.; Sun, H.; Wang, D.-C.; Shao, F. Pore-forming activity and structural autoinhibition of the gasdermin family. Nature 2016, 535, 111. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item 1 | Treatments 2 | SEM | p Value | ||
|---|---|---|---|---|---|
| CN | SI | PS | |||
| Pre-challenge (days 1 to 7) | |||||
| BW, kg (day 1) | 4.656 | 4.894 | 5.263 | 0.177 | 0.387 |
| ADG, g/d | 136.607 | 143.75 | 149.107 | 10.588 | 0.898 |
| G:F, g/g | 0.615 | 0.701 | 0.612 | 0.048 | 0.707 |
| Post-challenge (days 8 to 13) | |||||
| BW, kg (day 13) | 6.3 | 5.85 | 6.443 | 0.211 | 0.379 |
| ADG, g/d | 137.5 a | −10 b | 55 ab | 19.476 | 0.003 |
| G:F, g/g | 0.495 a | −0.049 b | 0.21 ab | 0.083 | 0.019 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Xia, B.; He, T.; Li, D.; Su, J.-H.; Guo, L.; Wang, J.-f.; Zhu, Y.-H. Oral Administration of a Select Mixture of Lactobacillus and Bacillus Alleviates Inflammation and Maintains Mucosal Barrier Integrity in the Ileum of Pigs Challenged with Salmonella Infantis. Microorganisms 2019, 7, 135. https://doi.org/10.3390/microorganisms7050135
Liu X, Xia B, He T, Li D, Su J-H, Guo L, Wang J-f, Zhu Y-H. Oral Administration of a Select Mixture of Lactobacillus and Bacillus Alleviates Inflammation and Maintains Mucosal Barrier Integrity in the Ileum of Pigs Challenged with Salmonella Infantis. Microorganisms. 2019; 7(5):135. https://doi.org/10.3390/microorganisms7050135
Chicago/Turabian StyleLiu, Xiao, Bing Xia, Ting He, Dan Li, Jin-Hui Su, Liang Guo, Jiu-feng Wang, and Yao-Hong Zhu. 2019. "Oral Administration of a Select Mixture of Lactobacillus and Bacillus Alleviates Inflammation and Maintains Mucosal Barrier Integrity in the Ileum of Pigs Challenged with Salmonella Infantis" Microorganisms 7, no. 5: 135. https://doi.org/10.3390/microorganisms7050135
APA StyleLiu, X., Xia, B., He, T., Li, D., Su, J. -H., Guo, L., Wang, J. -f., & Zhu, Y. -H. (2019). Oral Administration of a Select Mixture of Lactobacillus and Bacillus Alleviates Inflammation and Maintains Mucosal Barrier Integrity in the Ileum of Pigs Challenged with Salmonella Infantis. Microorganisms, 7(5), 135. https://doi.org/10.3390/microorganisms7050135