Epibiotic Fungal Communities of Three Tomicus spp. Infesting Pines in Southwestern China



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Library Construction
2.4. Data Analysis
3. Results
3.1. Data Characteristics
3.2. Epibiotic Community Diversity
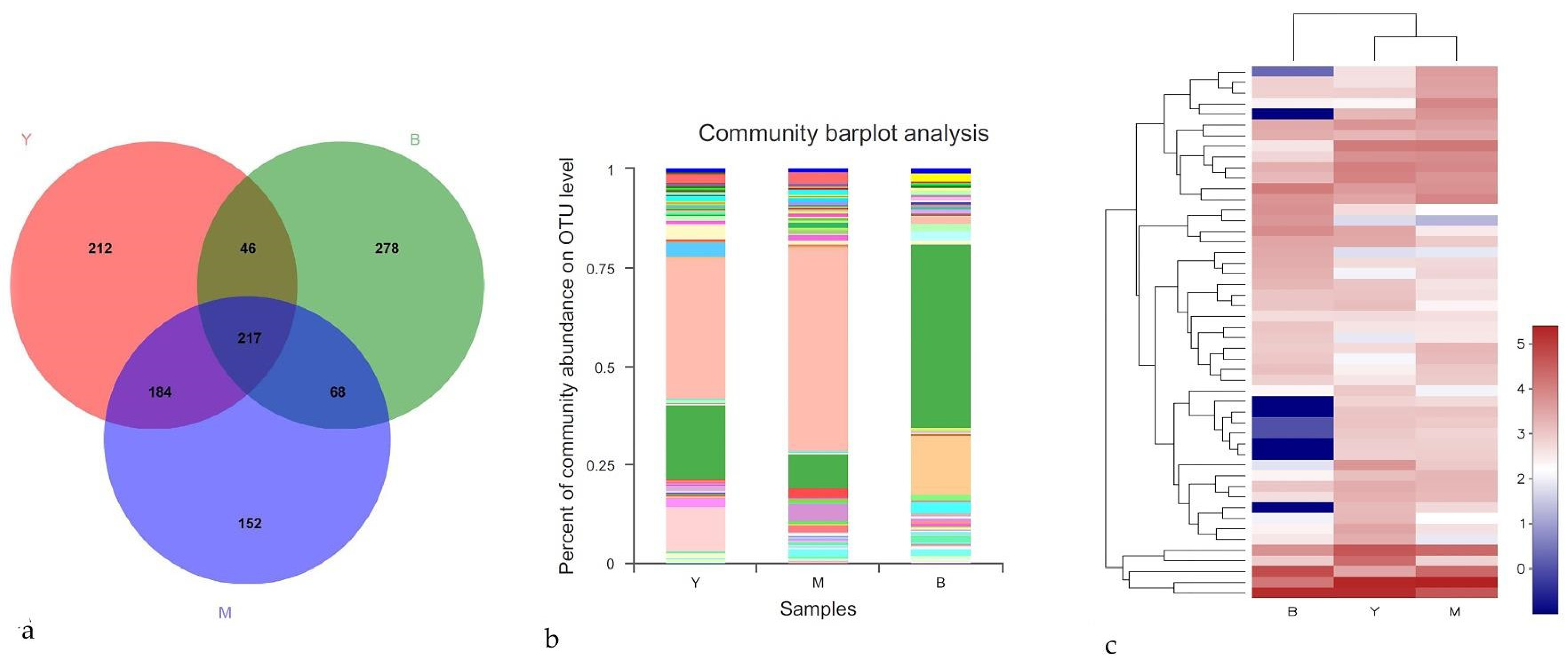
3.3. Taxonomic Identities of Fungal Communities
3.4. Factors Influencing Fungal Communities and Abundance Among P. yunnanensis Samples
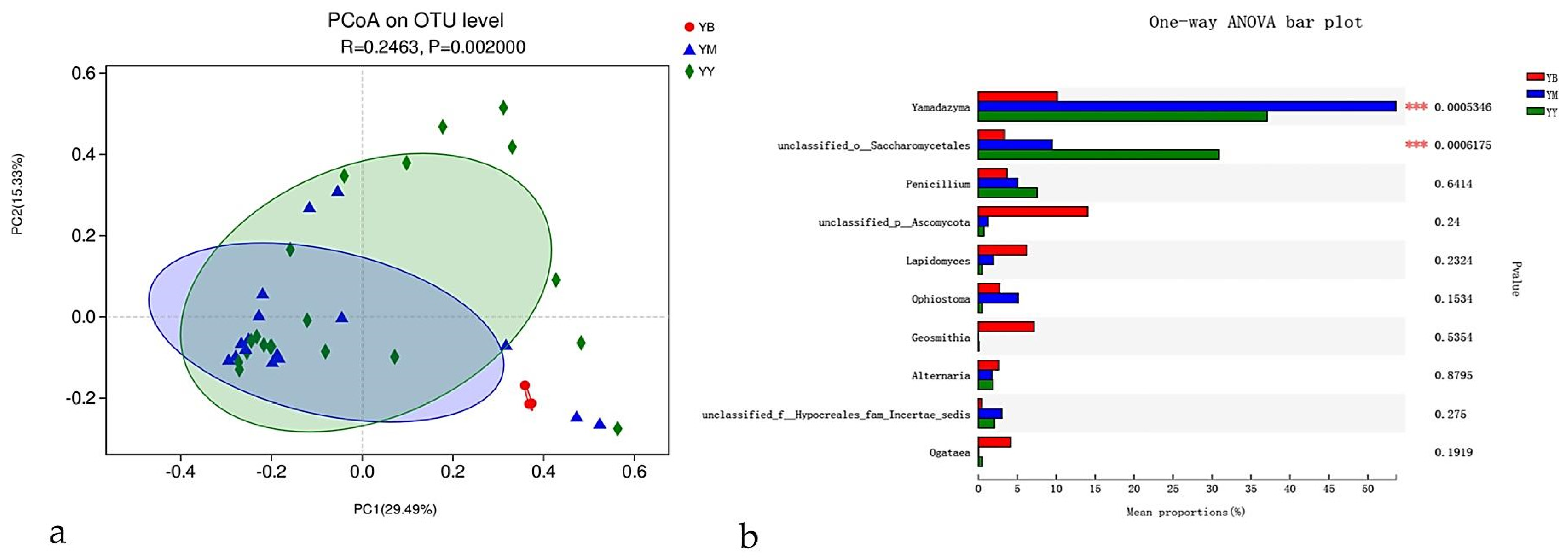
3.4.1. Beetle Species
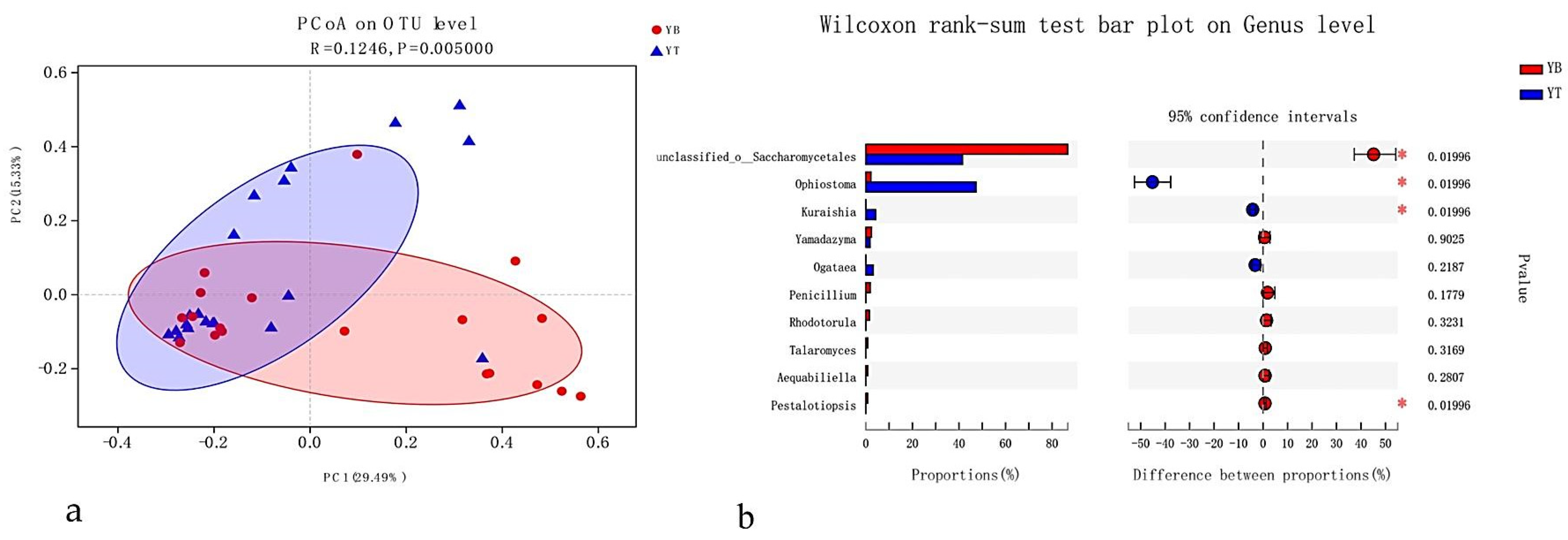
3.4.2. Infectious Section
3.4.3. Location
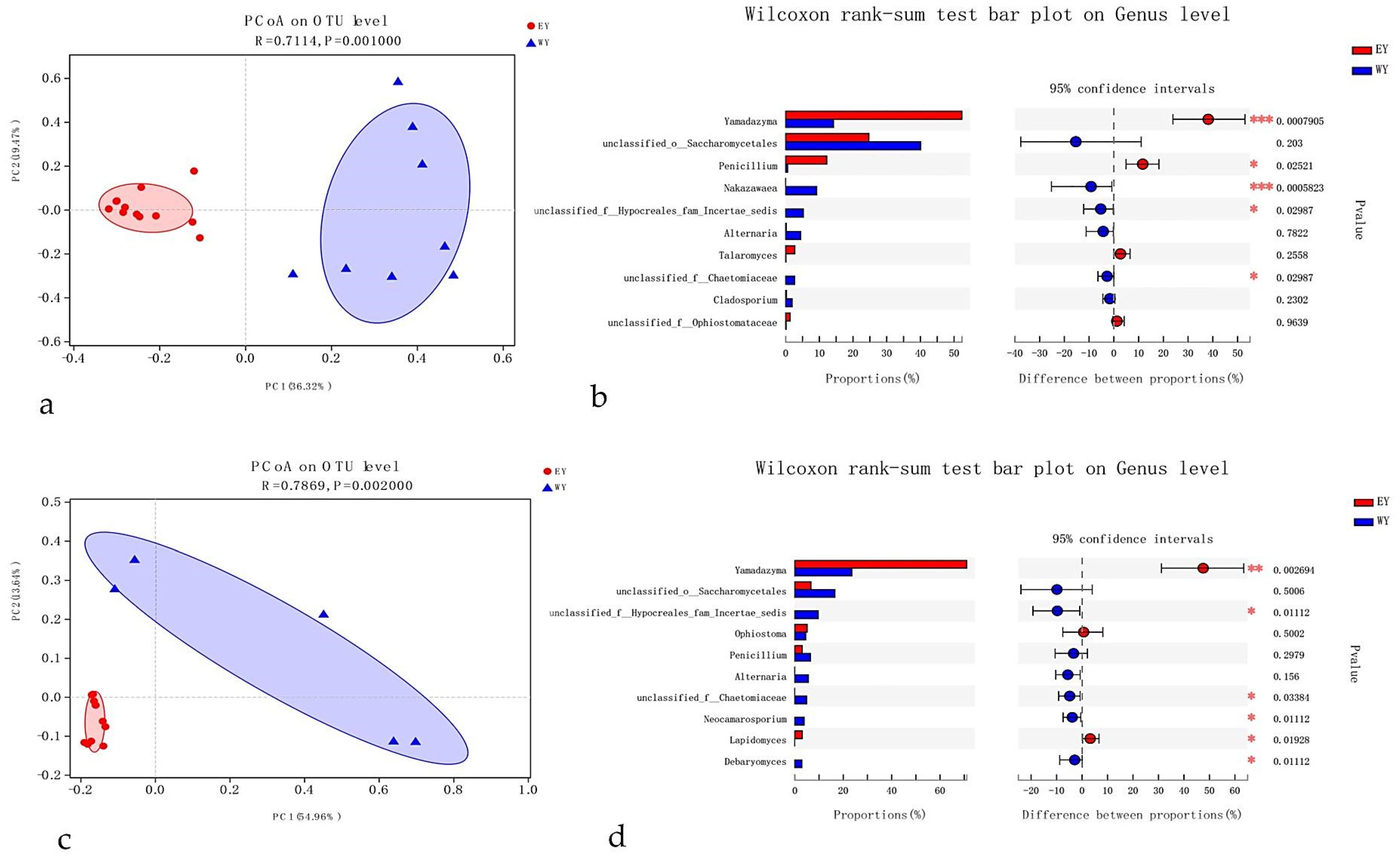
3.5. Hosts Influencing Fungal Communities and Abundance Associated with T. brevipilosus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hulcr, J.; Dunn, R.R. The sudden emergence of pathogenicity in insect-fungus symbioses threatens naïve forest ecosystems. Proc. R Soc. B. 2011, 278, 2866–2873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linnakoski, R.; de Beer, Z.W.D.; Niemelä, P.; Wingfield, M.J. Associations of conifer-infesting bark beetles and fungi in fennoscandia. Insects 2012, 3, 200–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Six, D.L.; Poulsen, M.; Hansen, A.K.; Wingfield, M.J.; Roux, J.; Eggleton, P.; Slippers, B.; Paine, T.D. Anthropogenic effects on interaction outcomes: Examples from insectmicrobial symbioses in forest and savanna ecosystems. Symbiosis 2011, 53, 101–121. [Google Scholar] [CrossRef]
- Wingfield, M.; Garnas, J.; Hajek, A.; Hurley, B.; Beer, Z.; Taerum, S. Novel and co-evolved associations between insects and microorganisms as drivers of forest pestilence. Biol. Invasions 2016, 18, 1045–1056. [Google Scholar] [CrossRef]
- Wingfield, M.J.; Slippers, B.; Wingfield, B.D. Novel associations between pathogens, insects and tree species threaten world forests. N. Z. J. For. Sci. 2010, 40, S95–S104. [Google Scholar]
- Dohet, L.; Grégoire, J.C.; Berasategui, A.; Kaltenpoth, M.; Biedermann, P.H. Bacterial and fungal symbionts of parasitic Dendroctonus bark beetles. FEMS Microbiol. Ecol. 2016, 92. [Google Scholar] [CrossRef] [Green Version]
- KnÍŽek, M.; Beaver, R. Taxonomy and Systematics of Bark and Ambrosia Beetles. In Bark and Wood Boring Insects in Living Trees in Europe, a Synthesis; Springer: Dordrecht, The Nederland, 2007. [Google Scholar]
- Von Schrenk, H. The bluing and the red rot of the western yellow pine with special reference to the Black hills forest reserve. US Bur. Plant Ind. Bull. 1903, 36, 1–40. [Google Scholar]
- Bleiker, K.; Six, D.L. Dietary benefits of fungal associates to an eruptive herbivore: Potential implications of multiple associates on host population dynamics. Environ. Entomol. 2007, 36, 1384–1396. [Google Scholar] [CrossRef]
- Lieutier, F.; Yart, A.; Salle, A. Stimulation of tree defenses by Ophiostomatoid fungi can explain attack success of bark beetles on conifers. Ann. For. Sci. 2009, 66, 801. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A. The microbial dimension in insect nutritional ecology. Funct. Ecol. 2009, 23, 38–47. [Google Scholar] [CrossRef]
- Gibson, C.M.; Hunter, M.S. Extraordinarily widespread and fantastically complex: Comparative biology of endosymbiotic bacterial and fungal mutualists of insects. Ecol. Lett. 2010, 13, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Paine, T.D.; Raffa, K.F.; Harrington, T.C. Interactions among Scolytid bark beetles, their associated fungi, and live host conifers. Annu. Rev. Entomol. 1997, 42, 179–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostovcik, M.; Bateman, C.C.; Kolarik, M.; Stelinski, L.L.; Jordal, B.H.; Hulcr, J. The ambrosia symbiosis is specific in some species and promiscuous in others: Evidence from community pyrosequencing. ISME J. 2015, 9, 126. [Google Scholar] [CrossRef] [Green Version]
- Wood, S.L.; Bright, D.E. A catalog of Scolytidae and Platypodidae (Coleoptera), Part 2: Taxonomic index. Great Basin Nat. Mem. 1992, 13, 1–6. [Google Scholar]
- Ye, H.; Ding, X.S. Impacts of Tomicus minor on distribution and reproduction of Tomicus piniperda (Col., Scolytidae) on the trunk of the living Pinus yunnanensis trees. J. Appl. Entomol. 1999, 123, 329–333. [Google Scholar]
- Jankowiak, R.; Kacprzyk, M.; Mlynarczyk, M. Diversity of ophiostomatoid fungi associated with bark beetles (Coleoptera: Scolytidae) colonizing branches of norway spruce (Picea abies) in southern Poland. Biologia 2009, 64, 1170–1177. [Google Scholar] [CrossRef] [Green Version]
- Raffa, K.F.; Phillips, T.W.; Salom, S.M. Strategies and mechanisms of host colonization by bark beetles. In Beetlepathogen Interactions in Conifer Forests; Schowalter, R.D., Filip, G.M., Eds.; Academic Press: London, UK, 1993; pp. 103–128. [Google Scholar]
- Liu, Z.; Xin, Y.; Xu, B.; Raffa, K.F.; Sun, J. Sound-triggered production of antiaggregation pheromone limits overcrowding of Dendroctonus valens attacking pine trees. Chem. Sens. 2017, 42, 59–67. [Google Scholar]
- Wood, S. The Bark and Ambrosia beetles of North and Central America (Coleoptera: Scolytidae): A Taxonomic Monograph; Great Basin Nat. Mem.; Brigham Young University Press: Provo, UT, USA, 1982; p. 1359. [Google Scholar]
- Lu, J.; Zhao, T.; Ye, H. The shoot-feeding ecology of three Tomicus species in Yunnan Province, southwestern China. J. Insect Sci. 2014, 14, 37. [Google Scholar] [CrossRef]
- Ye, H. Tomicus Yunnanensis; Yunnan Science and Technology Publishing Press: Kunming, Yunnan, China, 2011. [Google Scholar]
- Chen, P.; Lu, J.; Haack, R.A.; Ye, H. Attack Pattern and Reproductive Ecology of Tomicus brevipilosus (Coleoptera: Curculionidae) on Pinus yunnanensis in Southwestern China. J. Insect Sci. 2015, 15, 43. [Google Scholar] [CrossRef] [Green Version]
- Lu, R.; Wang, H.; Zhang, Z.; Byers, J.A.; Jin, Y.J.; Wen, H.F.; Shi, W.J. Coexistence and competition between Tomicus yunnanensis and T. minor (Coleoptera: Scolytinae) in Yunnan pine. A J. Entomol. 2012, 2012. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Li, L.; Liu, H. Host preference and competion in barkbeetles, Tomicus yunnanensis and T. minor in breeding period. J. West China For. Sci. 2010, 39, 15–20. [Google Scholar]
- Chen, P.; Li, L.; Liu, H. Interspecific competition between Tomicus yunnanensis and T. minor (Col., Scolytidae) during shoot-feeding period in Yunnan of China. J. West China For. Sci. 2009, 38, 52–60. [Google Scholar]
- Li, L.S.; Wang, H.L.; Chai, X.; Shu, N. Study on the Biological Characteristics of Tomicus piniperda and Its Damage; Yunnan Forestry Science and Technology: Kunming, Yunnan, China, 1997; pp. 1–7. [Google Scholar]
- Li, L.S.; Liu, H.P.; Chen, P.; Li, H.R.; Långström, B.; Francoies, L. Comparison study on ecological characteristics of Tomicus piniperda and T. minor. J. West China For. Sci. 2006, 35, 1–5. [Google Scholar]
- Ji, M.; Dong, X.Q.; Liu, H.P.; Li, L.S.; Xu, H.; Yang, X.P.; Li, H.R.; Ze, S.Z. Preliminary Study on Remote Sensing Detection of Yunnan Pine Forest Damaged by Tomicus piniperda. J. West China For. Sci. 2007, 36, 87–90. [Google Scholar]
- Ye, H.; Dang, C.L. The study on the feature of the bark beetle (Blastophagus piniperda L.) injuring the pine tree (Pinus yunnanensis). J. Yunnan Univ. 1986, 8, 218–221. [Google Scholar]
- Ceriani-Nakamurakare, E.; Ramos, S.; Robles, C.; Novas, M.; D´Jonsiles, M.; Gonzalez-Audino, P.; Carmarán, C. Metagenomic approach of associated fungi with Megaplatypus mutatus (Coleoptera: Platypodinae). Silva Fenn. 2018, 52. [Google Scholar] [CrossRef] [Green Version]
- Malacrinò, A.; Rassati, D.; Schena, L.; Mehzabin, R.; Battisti, A.; Palmeri, V. Fungal communities associated with bark and ambrosia beetles trapped at international harbours. Fungal Ecol. 2017, 28, 44–52. [Google Scholar] [CrossRef]
- Miller, K.E.; Hopkins, K.; Inward, D.J.G.; Vogler, A.P. Metabarcoding of fungal communities associated with bark beetles. Ecol. Evol. 2016, 6, 1590–1600. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.M.; Wang, Z.; Liu, F.; Wu, C.X.; Zhang, S.F.; Kong, X.B.; Decock, C.; Lu, Q.; Zhang, Z. Differential patterns of ophiostomatoid fungal communities associated with three sympatric Tomicus species infesting pines in southwestern China, with description of four new species. MycoKeys 2019, 50, 93–133. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Application; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- UPARSE. High-accuracy, high-throughput OTU clustering. Version 7.1. Available online: http://drive5.com/uparse (accessed on 6 October 2019).
- RDP. Classifier algorithm. Available online: http://rdp.cme.msu.edu (accessed on 6 October 2019).
- Unite Database. UNITE Mothur Release. Version Release 6.0. Available online: http://unite.ut.ee/index.php (accessed on 6 October 2019).
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Venny. Available online: https://omictools.com/venny-tool (accessed on 6 October 2019).
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Skelton, J.; Jusino, M.A.; Carlson, P.S.; Smith, K.; Banik, M.T.; Lindner, D.L.; Palmer, J.M.; Hulcr, J. Relationships among wood-boring beetles, fungi, and the decomposition of forest biomass. Mol. Ecol. 2019, 28. [Google Scholar] [CrossRef] [PubMed]
- Skelton, J.; Jusino, M.A.; Li, Y.; Bateman, C.; Thai, P.H.; Wu, C.X.; Lindner, D.L.; Hulcr, J. Detecting Symbioses in complex communities: The Fungal Symbionts of Bark and Ambrosia Beetles within Asian Pines. Microb. Ecol. 2018, 76, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.D.; Jacobs, K.; Morelet, M.; Ye, H.; Lieutier, F.; Wingfield, M.J. A new Leptographium species associated with Tomicus piniperda in South Western China. Mycoscience 2000, 41, 573–578. [Google Scholar] [CrossRef]
- Chang, R.; Duong, T.A.; Taerum, S.J.; Wingfield, M.J.; Zhou, X.D.; De Beer, Z.W. Ophiostomatoid fungi associated with conifer-infesting beetles and their phoretic mites in Yunnan, China. MycoKeys 2017, 28, 19. [Google Scholar] [CrossRef] [Green Version]
- Lieutier, F.; Ye, H.; Yart, A. Shoot damage by Tomicus sp. (Coleoptera: Scolytidae) and effect on Pinus yunnanensis resistance to subsequent reproductive attacks in the stem. Agric. For. Entomol. 2003, 5, 227–233. [Google Scholar] [CrossRef]
- Pan, Y.; Chen, P.; Lu, J.; Zhou, X.D.; Ye, H. First report of blue-stain in Pinus yunnanensis caused by Ophiostoma tingens associated with Tomicus minor in China. J. Plant Pathol. 2017, 99, 805. [Google Scholar]
- Zhou, X.D.; de Beer, Z.W.; Wingfield, M.J. Ophiostomatoid fungi associated with conifer infecting bark beetles in China. In Ophiostomatoid Fungi: Expanding Frontiers; Seifert, K.A., de Beer, Z.W., Wingfield, M.J., Eds.; CBS: Utrecht, The Netherlands, 2013; pp. 91–98. [Google Scholar]
- Kubátová, A.; Novotný, D.; Práil, K.; Mrácek, Z. The nematophagous hyphomycete Esteya vermicola found in the Czech Republic. Czech Mycol. 2000, 52, 227–235. [Google Scholar]
- Liou, J.Y.; Shih, J.Y.; Tzean, S.S. Esteya, a new nematophagous genus from Taiwan, attacking the pinewood nematode (Bursaphelenchus xylophilus). Mycol. Res. 1999, 103, 242–248. [Google Scholar] [CrossRef]
- Wang, C.Y.; Fang, Z.M.; Sun, B.S.; Gu, L.J.; Wang, Z.; Gu, L.J.; Sun, B.S.; Zhang, D.L.; Sung, C.K. High infection activities of two Esteya vermicola isolates against pinewood nematode. Afr. J. Microbiol. Res. 2009, 3, 581–584. [Google Scholar]
- Wang, X.; Wang, T.; Wang, J.; Guan, T.; Li, H. Morphological, molecular and biological characterization of Esteya vermicola, a nematophagous fungus isolated from intercepted wood packing materials exported from Brazil. Mycoscience 2014, 55, 367–377. [Google Scholar] [CrossRef]
- Yamaoka, Y.; Masuya, H.; Chung, W.H.; Goto, H.; To-Anun, C.; Tokumasu, S.; Zhou, X.; Wingfield, M.J. The teleomorph of Leptographium yunnanense, discovered in crosses among isolates from Thailand, China, and Japan. Mycoscience 2008, 49, 233–240. [Google Scholar] [CrossRef]
- Ye, H.; Zhou, X.; Lu, J.; Yang, L.; Ding, H. A preliminary study on the occurrence and pathogenicity of the fungi associated with Tomicus. For. Res. Beijing 2000, 13, 451–454. [Google Scholar]
- Ganter, P.F. Yeast and invertebrate associations. In Biodiversity and Ecophysiology of Yeasts; Pe´ter, G., Rosa, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 303–370. [Google Scholar]
- Davis, T.S. The ecology of yeasts in the bark beetle holobiont: A century of research revisited. Microb. Ecol. 2015, 69, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Lou, Q.Z.; Lu, M.; Sun, J.H. Yeast diversity associated with invasive Dendroctonus valens killing Pinus tabuliformis in China using culturing and molecular methods. Microb. Ecol. 2014, 68, 397–415. [Google Scholar] [CrossRef] [PubMed]
- Rivera, F.N.; Gonzalez, E.; Gomez, Z.; Lopez, N.; Herna’ndez-Rodrı’guez, C.; Berkov, A. Gut-associated yeast in bark beetles of the genus Dendroctonus Erichson (Coleoptera: Curculionidae: Scolytinae). Biol. J. Linn. Soc. 2009, 98, 325–342. [Google Scholar] [CrossRef] [Green Version]
- Six, D.L.; Paine, T.D. Effects of mycangial fungi and host tree species on progeny survival and emergence of Dendroctonus ponderosae (Coleoptera: Scolytidae). Environ. Entomol. 1998, 27, 1393–1401. [Google Scholar] [CrossRef]
- Whitney, H.S. Association of Dendroctonus ponderosae (Coleoptera: Scolytidae) with blue stain fungi and yeasts during brood development in lodgepole pine. Can. Entomol. 1971, 103, 1495–1503. [Google Scholar] [CrossRef]
- Whitney, H.S.; Farris, S.H. Maxillary mycangium in the mountain pine beetle. Science 1970, 167, 54–55. [Google Scholar] [CrossRef]
- Leufvén, A.; Bergström, G.; Falsen, E. Interconversion of verbenols and verbenone by identified yeasts isolated fromthe spruce bark beetle Ips typographus. J. Chem. Ecol. 1984, 10, 1349–1361. [Google Scholar] [CrossRef]
- Leufvén, A.; Bergström, G.; Falsen, E. Oxygenated monoterpenes produced by yeasts, isolated from Ips typographus (Coleoptera: Scolytidae) and grown in phloem medium. J. Chem. Ecol. 1988, 14, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Callaham, R.Z.; Shifrine, M. The yeasts associated with bark beetles. For. Sci. 1960, 6, 146–154. [Google Scholar]
- Strongman, D.B. A method for rearing Dendroctonus ponderosae Hopk. (Coleoptera: Scolytidae) from eggs to pupae on host tissue with or without a fungal complement. Can. Entomol. 1987, 119, 207–208. [Google Scholar] [CrossRef]
- Brand, J.M.; Schultz, J.; Barras, S.J.; Edson, L.J.; Payne, T.L.; Hedden, R.L. Bark-beetle pheromones: Enhancement of Dendroctonus frontalis (Coleoptera: Scolytidae) aggregation pheromone by yeast metabolites in laboratory bioassays. J. Chem. Ecol. 1977, 3, 657–666. [Google Scholar] [CrossRef]
- Hunt, D.W.A.; Borden, J.H. Conversion of verbenols to verbenone by yeasts isolated from Dendroctonus ponderosae (Coleoptera: Scolytidae). J. Chem. Ecol. 1990, 16, 1385–1397. [Google Scholar] [CrossRef]
- Davis, T.S.; Hofstetter, R.W. Reciprocal interactions between the bark beetle-associated yeast Ogataea pini and host plant phytochemistry. Mycologia 2011, 103, 1201–1207. [Google Scholar] [CrossRef] [Green Version]
- Lewinsohn, D.; Lewinsohn, E.; Bertagnolli, C.L.; Patridge, A.D. Blue-stain fungi and their transport structures on the Douglas-fir beetle. Can. J. For. Res. 1994, 24, 2275–2283. [Google Scholar] [CrossRef]
- Vaz, A.B.M.; Fonseca, P.L.C.; Leite, L.R.; Badotti, F.; Salim, A.C.M.; Araujo, F.M.G.; Cuadros -Orellana, S.; Duarte, A.A.; Rosa, C.A.; Oliveira, G.; et al. Using Next-Generation Sequencing (NGS) to uncover diversity of wood-decaying fungi in neotropical Atlantic forests. Phytotaxa 2017, 295, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Kasson, M.T.; O’Donnell, K.; Rooney, A.P.; Sink, S.; Ploetz, R.C.; Ploetz, J.N.; Konkol, J.L.; Carrillo, C.; Freeman, S.; Mendel, Z.; et al. An inordinate fondness for Fusarium: Phylogenetic diversity of fusaria cultivated by ambrosia beetles in the genus Euwallacea on Avocado and other plant hosts. Fungal Genet. Biol. 2013, 56, 147–157. [Google Scholar] [CrossRef]
- Romon, P.; Troya, M.; Fernandez de Gamarra, M.E.; Eguzkitza, A.; Iturrondobeitia, J.; Goldarazena, A. Fungal communities associated with pitch canker disease of Pinus radiata caused by Fusarium circinatum in northern Spain: Association with insects and pathogen-saprophyte antagonistic interactions. Can. J. Plant Pathol. 2008, 30, 241–253. [Google Scholar] [CrossRef]
- Ceriani-Nakamurakare, E.; Slodowicz, M.; Gonzalez-Audino, P.; Dolinko, A.; Carmarán, C. Mycobiota associated with the ambrosia beetle Megaplatypus mutatus: Threat to poplar plantations. Forestry 2016, 89, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Freeman, S.; Sharon, M.; Maymon, M.; Mendel, Z.; Protasov, A.; Aoki, T.; Eskalen, A.; O’Donell, K. Fusarium euwallaceae sp. nov.—A symbiotic fungus of Euwallacea sp., an invasive ambrosia beetle in Israel and California. Mycologia 2013, 105, 1595–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Short, D.P.; O’Donnell, K.; Stajich, J.E.; Hulcr, J.; Kijimoto, T.; Berger, M.C.; Macias, A.M.; Spahr, E.J.; Bateman, C.C.; Eskalen, A.; et al. PCR multiplexes discriminate Fusarium symbionts of invasive Euwallacea ambrosia beetles that inflict damage on numerous tree species throughout the United States. Plant Dis. 2017, 101, 233–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, H.; Wang, J.; Endoh, R.; Takeuchi, Y.; Tarno, H.; Futai, K. Pathogenicity of microorganisms isolated from the Oak platypodid, Platypus quercivorus (Murayama) (Coleoptera: Platypodidae). Appl. Entomol. Zool. 2011, 46, 201–210. [Google Scholar] [CrossRef]
- Scully, E.D.; Hoover, K.; Carlson, J.; Tien, M.; Geib, S.M. Proteomic analysis of Fusarium solani isolated from the Asian longhorned beetle, Anoplophora glabripennis. PLoS ONE 2012, 7, e32990. [Google Scholar] [CrossRef] [Green Version]
- Belhoucine, L.; Bouhraoua, R.T.; Meijer, M.; Houbraken, J.; Harrak, M.J.; Samson, R.A.; Equihua-Martinez, A.; Pujade-Villar, J. Mycobiota associated with Platypus cylindrus (Coleoptera: Curculionidae, Platypodidae) in cork oak stands of north west Algeria, Africa. Afr. J. Microbiol. Res. 2011, 5, 4411–4423. [Google Scholar] [CrossRef]
- Repe, A.; Kirisits, T.; Piskur, B.; De Groot, M.; Kump, B.; Jurc, M. Ophiostomatoid fungi associated with three spruce-infesting bark beetles in Slovenia. Ann. For. Sci. 2013, 70, 717–727. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
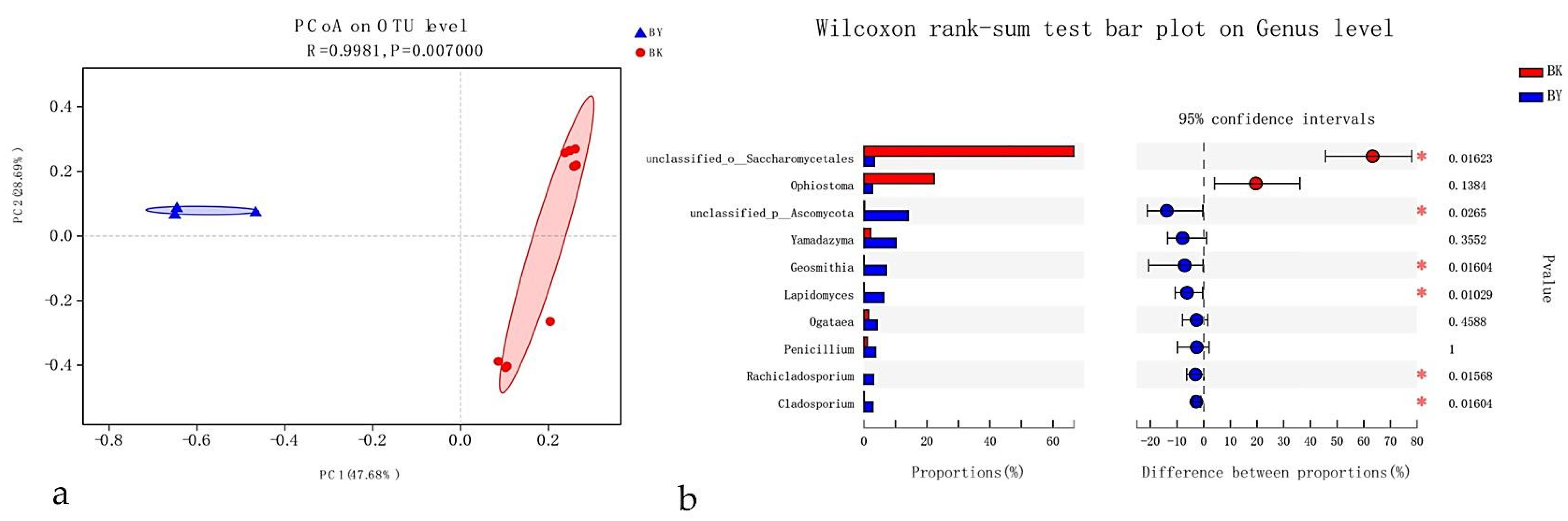{kind=link}
| Sample\Estimators | Samples Number | Chao 1 | Shannon | ||||||
|---|---|---|---|---|---|---|---|---|---|
| B | T | B-T | B | T | B-T | B | T | B-T | |
| Tomicus brevipilosus | 7 | 5 | 12 | 118.58 ± 29.43 | 146.47 ± 33.46 | 139.9 ± 40.45 | 1.57 ± 0.97 | 2 ± 0.82 | 1.75 ± 0.56 |
| T. minor | 9 | 7 | 16 | 173.84 ± 70.26 | 64.01 ± 25.91 | 125.79 ± 77.71 | 2.12 ± 0.89 | 1.28 ± 0.41 | 1.75 ± 0.83 |
| T. yunnanensis | 8 | 12 | 20 | 146.04 ± 69.63 | 71.6 ± 24.82 | 101.38 ± 60.07 | 2.08 ± 1.17 | 1.31 ± 0.27 | 1.62 ± 0.86 |
| Taxon | Host | Beetle | References |
|---|---|---|---|
| Esteya vermicola | P. yunnanensis | Tomicus yunnanensis | [34] |
| Graphilbum anningense | P. yunnanensis | T. yunnanensis, T. minor | [34] |
| Gra. fragrans | P. yunnanensis | T. minor | [34] |
| Leptographium yunnanensis | P. yunnanensis, P. kesiya | T. yunnanensis, T. brevipilosus | [34,44] |
| Ophiostoma aggregatum | P. yunnanensis | T. yunnanensis, T. minor | [34] |
| O. brevipilosi | P. kesiya | T. brevipilosus | [34,46] |
| O. canum | P. yunnanensis | T. yunnanensis, T. minor | [34] |
| O. ips | P. yunnanensis | T. yunnanensis | [48] |
| O. minus | P. yunnanensis | T. yunnanensis | [54] |
| O. quercus | P. yunnanensis | T. yunnanensis | [48] |
| Ophiostoma sp. 1 | P. yunnanensis | T. yunnanensis | [34] |
| O. tingens | P. yunnanensis | T. yunnanensis, T. minor | [34,47] |
| Sporothrix abietina | P. yunnanensis | T. yunnanensis | [43] |
| S. macroconidia | P. yunnanensis, P. kesiya | T. yunnanensis, T. brevipilosus | [34] |
| S. nebularis | P. yunnanensis | T. yunnanensis | [45] |
| S. pseudoabietina | P. yunnanensis | T. minor | [34] |
| Order | Genus | Possible Species | OTU | Reads No. |
|---|---|---|---|---|
| Ophiostomatales | Ceratocystiopsis | C. minima | OTU379, OTU243 | 41 |
| Graphilbum | Gra. microcarpum | OTU434 | 31 | |
| Graphilbum anningense | OTU326 | 1255 | ||
| Grosmannia | Grosmannia aurea | OTU677 | 7 | |
| G. yunnanense | OTU433 | 527 | ||
| Ophiostoma | O. macrosporum\ O. tingens | OTU391, OTU1178 | 6349 | |
| O. brevipilosi | OTU273, OTU272, OTU277, OTU370, OTU278, OTU375, OTU194, OTU281, OTU286, OTU304, OTU306, OTU308, OTU353, OTU332, OTU342, OTU349, OTU348, OTU269, OTU266, OTU263, OTU294, OTU222, OTU221, OTU227, OTU224, OTU223, OTU367, OTU255, OTU59, OTU254, OTU318, OTU317, OTU259, OTU350, OTU354, OTU359, OTU347, OTU210, OTU217, OTU215, | 56,779 | ||
| O. tapionis | OTU343, OTU327, OTU248, OTU298, OTU369, OTU360, OTU338, OTU262, OTU203, OTU290, OTU296, OTU1286, OTU393, | 27 | ||
| O. canum | OTU393, OTU175 | 30,863 | ||
| O. nigrocarpum | OTU1286 | 4 | ||
| O. pusillum | OTU363 | 37 | ||
| O. minus | OTU963, OTU432 | 1256 | ||
| Sporothrix | Sporothrix sp. | OTU218 | 4979 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.-M.; Liu, F.; Zhang, S.-F.; Kong, X.-B.; Lu, Q.; Zhang, Z. Epibiotic Fungal Communities of Three Tomicus spp. Infesting Pines in Southwestern China. Microorganisms 2020, 8, 15. https://doi.org/10.3390/microorganisms8010015
Wang H-M, Liu F, Zhang S-F, Kong X-B, Lu Q, Zhang Z. Epibiotic Fungal Communities of Three Tomicus spp. Infesting Pines in Southwestern China. Microorganisms. 2020; 8(1):15. https://doi.org/10.3390/microorganisms8010015
Chicago/Turabian StyleWang, Hui-Min, Fu Liu, Su-Fang Zhang, Xiang-Bo Kong, Quan Lu, and Zhen Zhang. 2020. "Epibiotic Fungal Communities of Three Tomicus spp. Infesting Pines in Southwestern China" Microorganisms 8, no. 1: 15. https://doi.org/10.3390/microorganisms8010015
APA StyleWang, H. -M., Liu, F., Zhang, S. -F., Kong, X. -B., Lu, Q., & Zhang, Z. (2020). Epibiotic Fungal Communities of Three Tomicus spp. Infesting Pines in Southwestern China. Microorganisms, 8(1), 15. https://doi.org/10.3390/microorganisms8010015