Exploring Beneficial Properties of the Bacteriocinogenic Enterococcus faecium ST10Bz Strain Isolated from Boza, a Bulgarian Cereal-Based Beverage

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Bacteriocinogenic Strains and their Inhibitory Activity
2.2. Selection and Identification of Bacterial Isolates
2.3. Detection of Known Bacteriocin Genes in the Genome of the Studied Strain
2.4. Evaluation of Bacteriocin Production
2.4.1. Growth Production Experiment
2.4.2. Stability of Antimicrobial Peptides
2.4.3. Optimization of Culture Condition for Bacteriocin Production
2.4.4. Bacteriocin Mode of Action
2.4.5. Adsorption of Bacteriocin to Listeria monocytogenes ATCC®15313™
2.4.6. Spectrum of Activity
2.5. Safety Tests and Detection of Additional Beneficial Properties
2.5.1. Antibiotic Susceptibility Test
2.5.2. Hemolysis Test
2.5.3. Biogenic Amine Production Test
2.5.4. Detection of Adhesion and Gamma Aminobutyric Acid (GABA) Production Related Genes
2.5.5. Gastrointestinal Simulation
2.5.6. Enzyme Profile
3. Results
3.1. Isolation and Molecular Characterization of the Bacteriocinogenic Strain
3.2. Bacteriocin Production of the Strain
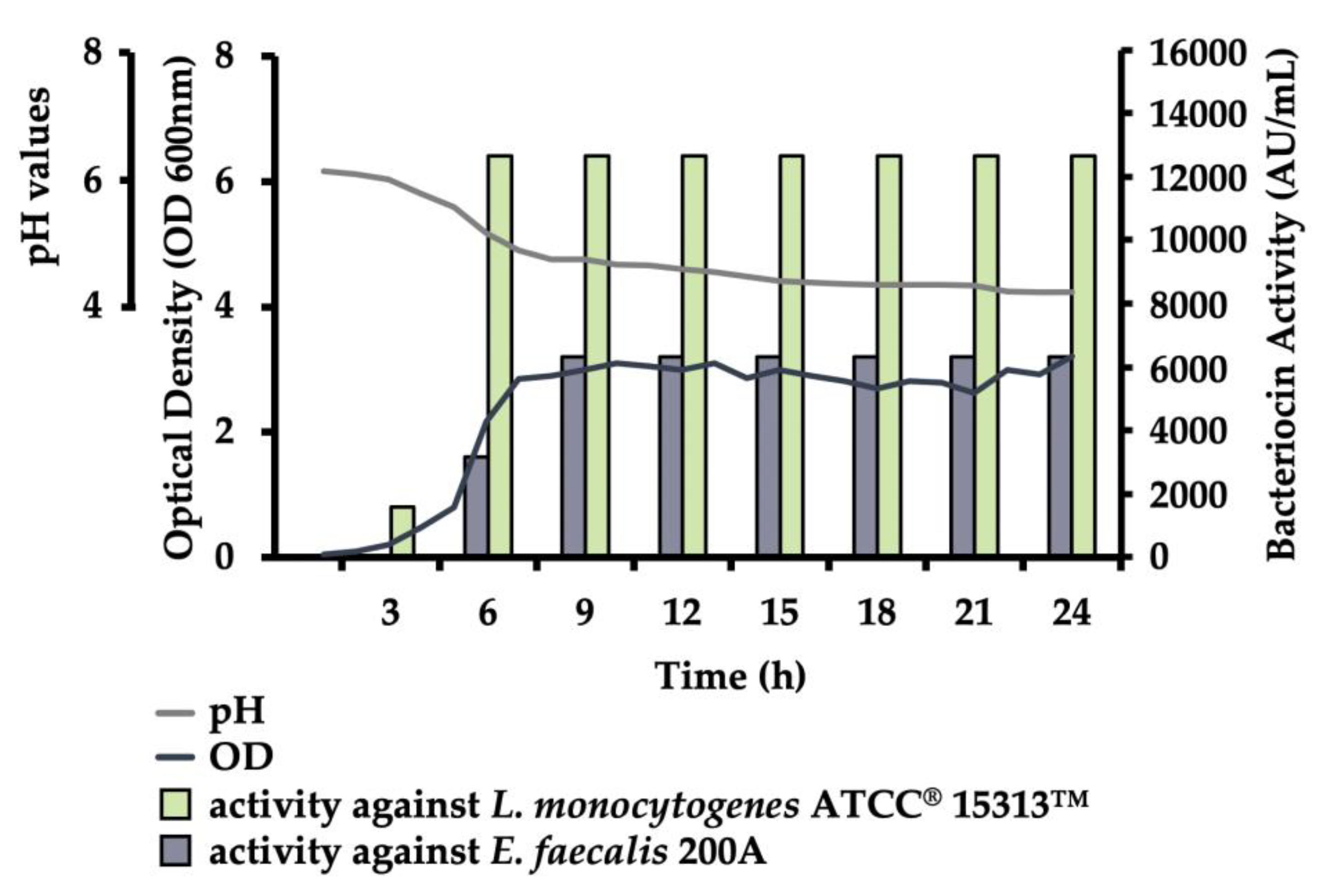
3.2.1. Growth Production
3.2.2. Stability of Bacteriocins
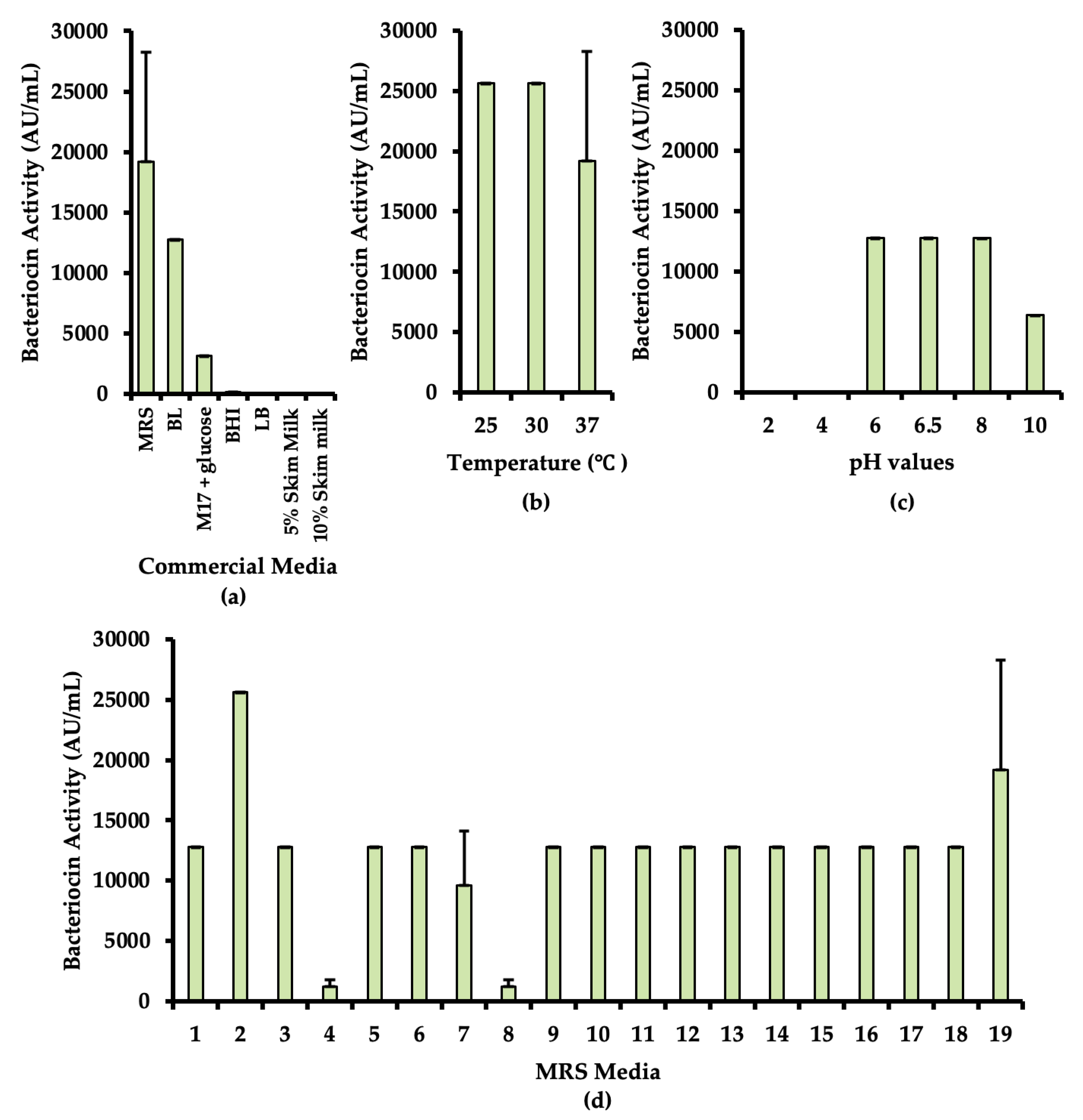
3.2.3. Optimization of Culture Medium
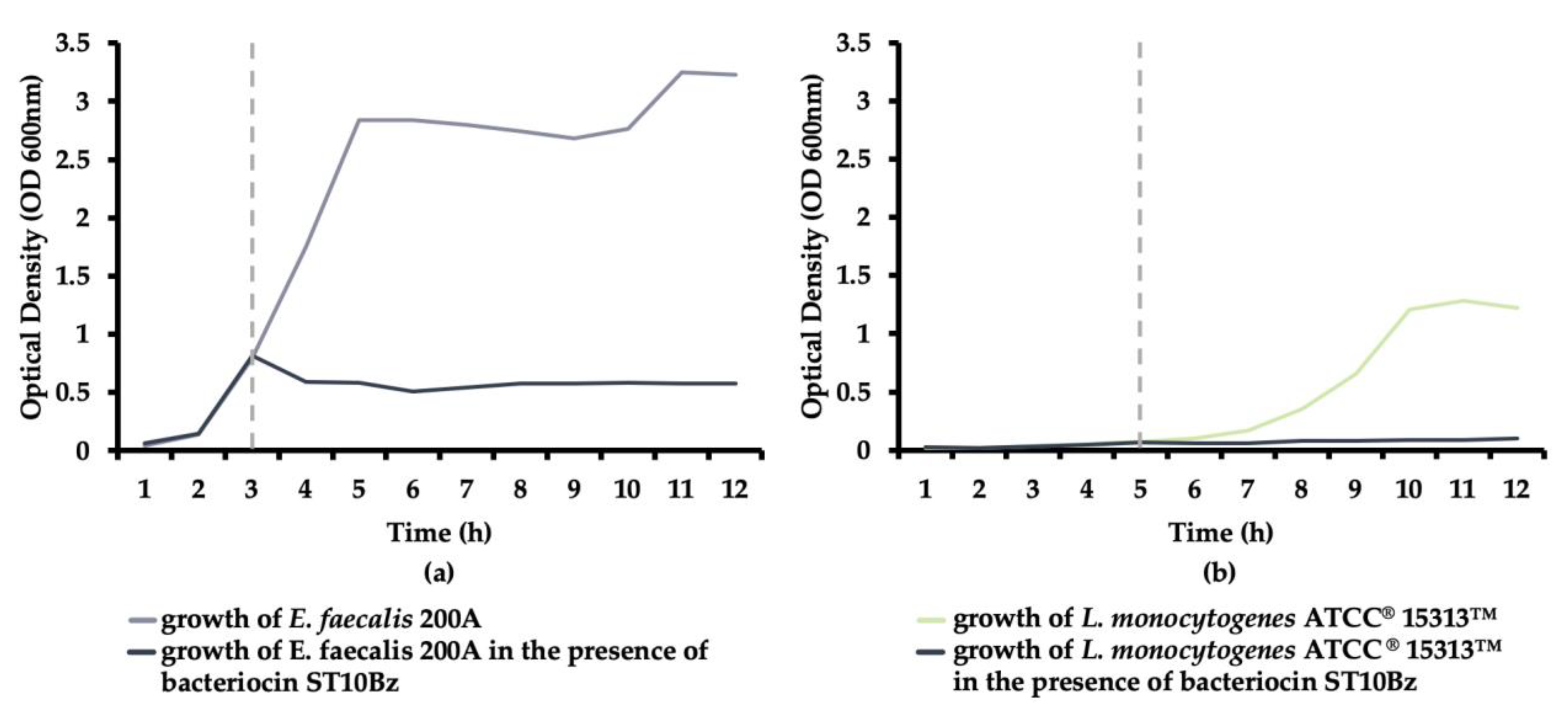
3.2.4. Bacteriocin Mode of Action
3.2.5. Spectrum of Activity
3.3. Safety Tests and Detection of Additional Beneficial Properties
3.3.1. Antibiotic Resistance, Hemolysis, and Biogenic Amine Production
3.3.2. Detection of Adhesion and Gamma Aminobutyric Acid (GABA) Production Related Genes
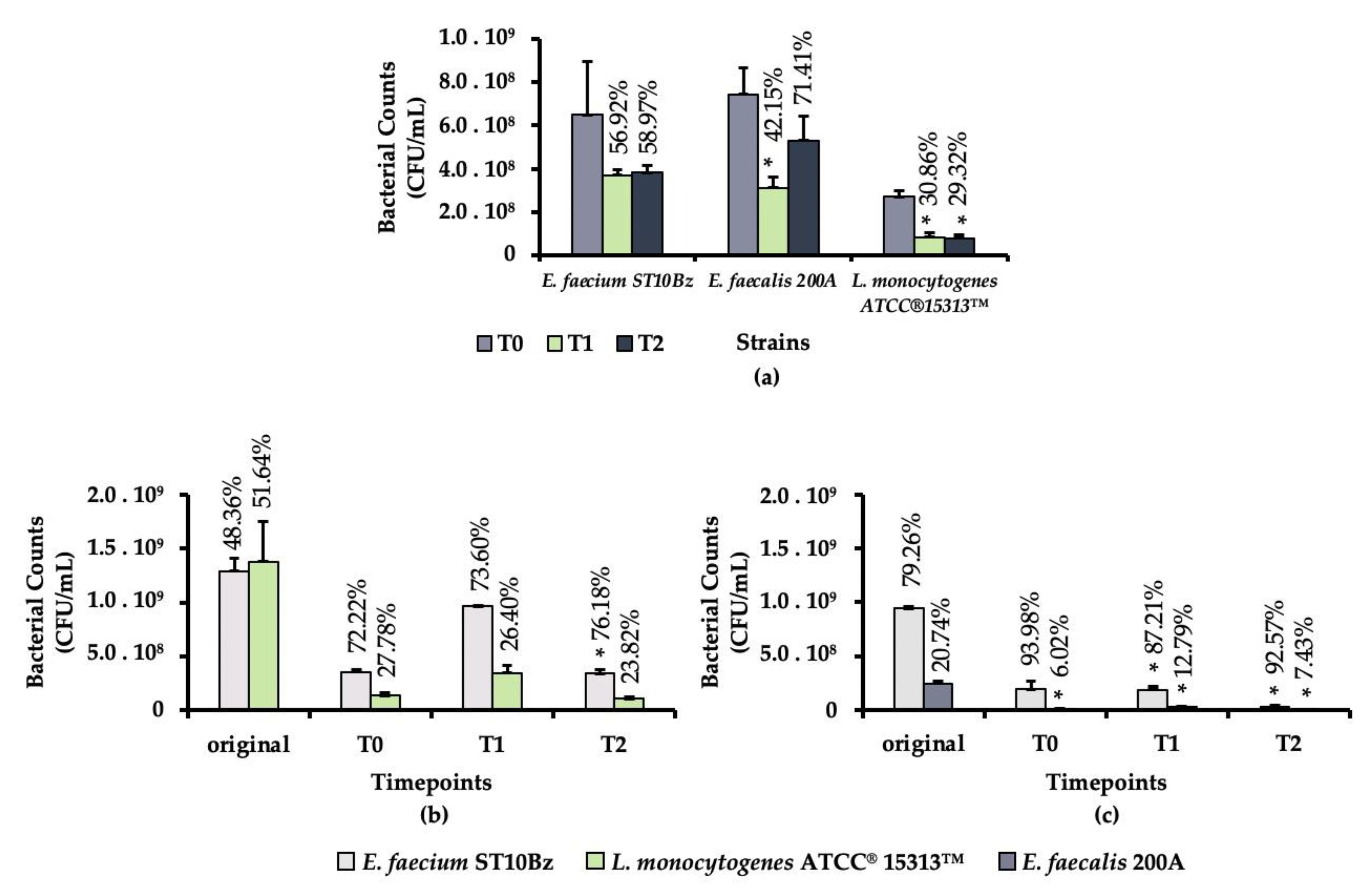
3.3.3. GIT Model
3.3.4. Enzymes Production Profile
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lahtinen, S.; Ouwehand, A.; Salminen, S.; Wright, A.V. Lactic Acid Bacteria: Microbiological and Functional aspects, 4th ed.; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar]
- Atanasova, J.; Dalgalarrondo, M.; Iliev, I.; Moncheva, P.; Todorov, S.D.; Ivanova, I.V. Formation of free amino acids and bioactive peptides during ripening of Bulgarian white brined cheeses. Probiotics Antimicrob. Proteins 2020. [Google Scholar] [CrossRef] [PubMed]
- Sulistiani; Handayani, R. Application biopreservatives produced by lactic acid bacteria (LAB) for preservation boiled-salted (pindang) tuna (Euthynnusaffinis Cantor, 1849). AIP Conf. Proc. 2018, 2002. [Google Scholar] [CrossRef]
- Wegh, C.A.M.; Geerlings, S.Y.; Knol, J.; Roeselers, G.; Belzer, C. Postbiotics and their potential applications in early life nutrition and beyond. Int. J. Mol. Sci. 2019, 20, 4673. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez Ronquillo, M.; Angeles Hernandez, J.C. Antibiotic and synthetic growth promoters in animal diets: Review of impact and analytical methods. Food Control. 2017, 72, 255–267. [Google Scholar] [CrossRef]
- Antibiotic Resistance. Available online: https://www.who.int/news-room/fact-sheets/detail/antibiotic-resistance (accessed on 15 June 2020).
- Todorov, S.; Franco, B.D.M.; Tagg, J. Bacteriocins of Gram-positive bacteria having activity spectra extending beyond closely-related species. Benef. Microbes 2019, 10, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Jordan, K.; Leong, D.; Ordóñez, A.Á. Listeria monocytogenes in the Food Processing Environment; Springer: Cham, Switzerland, 2015. [Google Scholar] [CrossRef]
- Lopetuso, L.; Giorgio, M.; Saviano, A.; Scaldaferri, F.; Gasbarrini, A.; Cammarota, G. Bacteriocins and bacteriophages: Therapeutic weapons for gastrointestinal diseases. Int. J. Mol. Sci. 2019, 20, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chikindas, M.L.; Weeks, R.; Drider, D.; Chistyakov, V.A.; Dicks, L.M.T. Functions and emerging applications of bacteriocins. Curr. Opin. Biotechnol. 2018, 49, 23–28. [Google Scholar] [CrossRef]
- Inglis, R.F.; Scanlan, P.; Buckling, A. Iron availability shapes the evolution of bacteriocin resistance in Pseudomonas aeruginosa. ISME J. 2016, 10, 2060–2066. [Google Scholar] [CrossRef]
- Listeriosis. Available online: https://www.who.int/news-room/fact-sheets/detail/listeriosis#targetText=It%20is%20a%20relatively%20rare,a%20significant%20public%20health%20concern (accessed on 15 June 2020).
- Roesel, K.; Grace, D. Food Safety and Informal Markets: Animal Products in Sub-Saharan Africa; Routledge: London, UK, 2015. [Google Scholar]
- Listeria (Listeriosis). Available online: https://www.fda.gov/food/foodborne-pathogens/listeria-listeriosis (accessed on 15 June 2020).
- Camargo, A.C.; Moura, A.; Avillan, J.; Herman, N.; McFarland, A.P.; Sreevatsan, S.; Call, D.R.; Woodward, J.J.; Lecuit, M.; Nero, L.A. Whole-genome sequencing reveals Listeria monocytogenes diversity and allows identification of long-term persistent strains in Brazil. Environ. Microbiol. 2019, 21, 4478–4487. [Google Scholar] [CrossRef]
- Terzić-Vidojević, A.; Veljović, K.; Begović, J.; Filipić, B.; Popović, D.; Tolinački, M.; Miljković, M.; Kojić, M.; Golić, N. Diversity and antibiotic susceptibility of autochthonous dairy enterococci isolates: Are they safe candidates for autochthonous starter cultures? Front. Microbiol. 2015, 6, 954. [Google Scholar] [CrossRef] [Green Version]
- Hanchi, H.; Mottawea, W.; Sebei, K.; Hammami, R. The genus Enterococcus: Between probiotic potential and safety concerns—An update. Front. Microbiol. 2018, 9, 1791. [Google Scholar] [CrossRef] [PubMed]
- Cetinkaya, Y.; Falk, P.; Mayhall, C.G. Vancomycin-Resistant Enterococci. Clin. Microbiol. Rev. 2001, 13, 686–707. [Google Scholar] [CrossRef]
- Dos Santos, K.M.O.; de Matos, C.R.; Salles, H.O.; Franco, B.D.G.M.; Arellano, K.; Holzapfel, W.H.; Todorov, S.D. Exploring beneficial/virulence properties of two dairy-related strains of Streptococcus infantarius subsp. infantarius. Probiotics Antimicrob. Proteins 2020. [Google Scholar] [CrossRef] [PubMed]
- De Vos, P.; Garrity, G.M.; Jones, D.; Krieg, N.R.; Ludwig, W.; Rainey, F.A.; Schleifer, K.-H.; Whitman, W.B. Bergeys Manual of Systematic Bacteriology: The Firmicutes; Springer: London, UK, 2009. [Google Scholar]
- De Moraes, G.M.D.; de Abreu, L.R.; do Egito, A.S.; Salles, H.O.; da Silva, L.M.F.; Nero, L.A.; Todorov, S.D.; dos Santos, K.M.O. Functional properties of Lactobacillus mucosae strains isolated from Brazilian goat milk. Probiotics Antimicrob. Proteins 2016, 9, 235–245. [Google Scholar] [CrossRef]
- Aymerich, T.; Holo, H.; Havarstein, L.S.; Hugas, M.; Garriga, M.; Nes, I.F. Biochemical and genetic characterization of enterocin A from Enterococcus faecium, a new antilisterial bacteriocin in the pediocin family of bacteriocins. Appl. Environ. Microbiol. 1996, 62, 1676–1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du Toit, M.; Franz, C.M.; Dicks, L.M.T.; Holzapfel, W.H. Preliminary characterization of bacteriocins produced by Enterococcus faecium and Enterococcus faecalis isolated from pig faeces. J. Appl. Microbiol. 2000, 88, 482–494. [Google Scholar] [CrossRef]
- Gutierrez, J.; Criado, R.; Citti, R.; Martin, M.; Herranz, C.; Nes, I.F.; Cintas, L.M.; Hernandez, P.E. Cloning, production and functional expression of enterocin P, a sec-dependent bacteriocin produced by Enterococcus faecium P13, in Escherichia coli. Int. J. Food. Microbiol. 2005, 103, 239–250. [Google Scholar] [CrossRef]
- Todorov, S.D.; Wachsman, M.; Tomé, E.; Dousset, X.; Destro, M.T.; Dicks, L.M.T.; Franco, B.D.G.M.; Vaz-Velho, M.; Drider, D. Characterisation of an antiviral pediocin-like bacteriocin produced by Enterococcus faecium. Food Microbiol. 2010, 27, 869–879. [Google Scholar] [CrossRef]
- Favaro, L.; Basaglia, M.; Casella, S.; Hue, I.; Dousset, X.; Franco, B.D.G.M.; Todorov, S.D. Bacteriocinogenic potential and safety evaluation of non-starter Enterococcus faecium strains isolated from homemade white brine cheese. Food Microbiol. 2014, 38, 228–239. [Google Scholar] [CrossRef]
- Todorov, S.D.; Dicks, L.M.T. Parameters affecting the adsorption of plantaricin 423, a bacteriocin produced by Lactobacillus plantarum 423 isolated from sorghum beer. Biotechnol. J. 2006, 1, 405–409. [Google Scholar] [CrossRef]
- Arellano, K.; Vazquez, J.; Park, H.; Lim, J.; Ji, Y.; Kang, H.-J.; Cho, D.; Jeong, H.W.; Holzapfel, W.H. Safety evaluation and whole-genome annotation of Lactobacillus plantarum strains from different sources with special focus on isolates from green tea. Probiotics Antimicrob. Proteins 2019, 12, 1057–1070. [Google Scholar] [CrossRef]
- European Food Safety Authority. Guidance on the assessment of bacterial susceptibility to antimicrobials of human and veterinary importance. EFSA J. 2012, 10, 1–10. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 11th ed.; CLSI: Wayne, PA, USA, 2018. [Google Scholar]
- Manson, J.M.; Keis, S.; Smith, J.M.B.; Cook, G.M. A Clonal Lineage of VanA-Type Enterococcus faecalis Predominates in Vancomycin-Resistant Enterococci Isolated in New Zealand. Antimicrob. Agents Chemother. 2003, 47, 204–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iweriebor, B.C.; Obi, L.C.; Okoh, A.I. Virulence and antimicrobial resistance factors of Enterococcus spp. isolated from fecal samples from piggery farms in Eastern Cape, South Africa. BMC Microbiol. 2015, 15, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depardieu, F.; Perichon, B.; Courvalin, P. Detection of the van Alphabet and Identification of Enterococci and Staphylococci at the Species Level by Multiplex PCR. J. Clin. Microbiol. 2004, 42, 5857–5860. [Google Scholar] [CrossRef] [Green Version]
- Morandi, S.; Silvetti, T.; Brasca, M. Biotechnological and safety characterization of Enterococcus lactis, a recently described species of dairy origin. Antonie van Leeuwenhoek 2012, 103, 239–249. [Google Scholar] [CrossRef]
- Du, F.; Lv, X.; Duan, D.; Wang, L.; Huang, J. Characterization of a Linezolid- and Vancomycin-Resistant Streptococcus suis Isolate That Harbors optrA and vanG Operons. Front. Microbiol. 2019, 10, 2026. [Google Scholar] [CrossRef]
- Todorov, S.D.; Dicks, L.M.T. Evaluation of lactic acid bacteria from kefir, molasses and olive brine as possible probiotics based on physiological properties. Ann. Microbiol. 2008, 58, 661–670. [Google Scholar] [CrossRef]
- De Castilho, N.P.A.; Colombo, M.; de Oliveira, L.L.; Todorov, S.D.; Nero, L.A. Lactobacillus curvatus UFV-NPAC1 and other lactic acid bacteria isolated from calabresa, a fermented meat product, present high bacteriocinogenic activity against Listeria monocytogenes. BMC Microbiol. 2019, 19, 63–75. [Google Scholar] [CrossRef] [Green Version]
- Bajić, S.S.; Đokić, J.; Dinić, M.; Tomić, S.; Popović, N.; Brdarić, E.; Goli, N.; Tolinački, M. GABA potentiate the immunoregulatory effects of Lactobacillus brevis BGZLS10-17 via ATG5-dependent autophagy in vitro. Sci. Rep. 2020, 10, 1347. [Google Scholar] [CrossRef]
- Ray, R.C.; Montet, D. Fermented Foods, Part. II Technological Interventions; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Todorov, S.D.; Dicks, L.M.T. Screening for bacteriocin-producing lactic acid bacteria from boza, a traditional cereal beverage from Bulgaria. Process. Biochem. 2006, 41, 11–19. [Google Scholar] [CrossRef]
- Makhloufi, K.M.; Carré-Mlouka, A.; Peduzzi, J.; Lombard, C.; van Reenen, C.A.; Dicks, L.M.T.; Rebuffat, S. Characterization of leucocin B-KM432Bz from Leuconostoc pseudomesenteroides isolated from boza, and comparison of its efficiency to pediocin PA-1. PLoS ONE 2013, 8, e70484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Mollendorff, J.W.; Todorov, S.D.; Dicks, L.M.T. Optimization of growth medium for production of bacteriocins produced by Lactobacillus plantarum JW3BZ and JW6BZ, and Lactobacillus fermentum JW11BZ and JW15BZ isolated from Boza. Trakia J. Sci. 2009, 7, 22–23. [Google Scholar]
- Charyyev, M.G.; Tuncer, B.Ö.; Kankaya, D.A.; Tuncer, Y. Bacteriocinogenic properties and safety evaluation of Enterococcus faecium YT52 isolated from boza, a traditional cereal based fermented beverage. J. Consum. Prot. Food Saf. 2019, 14, 41–53. [Google Scholar] [CrossRef]
- Barbosa, M.S.; Todorov, S.D.; Ivanova, I.; Chobert, J.-M.; Haertlé, T.; Franco, B.D.G.M. Improving safety of salami by application of bacteriocins produced by an autochthonous Lactobacillus curvatus isolate. Food Microbiol. 2015, 46, 254–262. [Google Scholar] [CrossRef] [Green Version]
- Sawa, N.; Okamura, K.; Zendo, T.; Himeno, K.; Nakayama, J.; Sonomoto, K. Identification and characterization of novel multiple bacteriocins produced by Leuconostoc pseudomesenteroides QU 15. J. Appl. Microbiol. 2010, 109, 282–291. [Google Scholar] [CrossRef]
- Garcia-Gutierrez, E.; O’Connor, P.M.; Colquhoun, I.J.; Vior, N.M.; Rodriguez, J.M.; Mayer, M.J.; Cotter, P.D.; Narbad, A. Production of multiple bacteriocins, including the novel bacteriocin gassericin M, by Lactobacillus gasseri LM19, a strain isolated from human milk. Appl. Microbiol. Biotechnol. 2020, 104, 3869–3884. [Google Scholar] [CrossRef] [Green Version]
- Mirkovic, N.; Polovic, N.; Vukotic, G.; Jovcic, B.; Miljkovic, M.; Radulovic, Z.; Diep, D.B.; Kojic, M. Lactococcus lactis LMG2081 produces two bacteriocins, a nonlantibiotic and a novel lantibiotic. Appl. Environ. Microb. 2016, 82, 2555–2562. [Google Scholar] [CrossRef] [Green Version]
- Himeno, K.; Fujita, K.; Zendo, T.; Wilaipun, P.; Ishibashi, N.; Masuda, Y.; Yoneyama, F.; Leelawatcharamas, V.; Nakayama, J.; Sonomoto, K. Identification of enterocin NKR-5-3C, a novel class IIa bacteriocin produced by a multiple bacteriocin producer, Enterococcus faecium NKR-5-3. Biosci. Biotechnol. Biochem. 2012, 76, 1245–1247. [Google Scholar] [CrossRef]
- Aguilar-Galvez, A.; Guillermo, S.; Dubois-Dauphin, R.; Campos, D.; Thonart, P. The influence of growth conditions on enterocin-like production by Enterococcus faecium CWBI-B1430 and Enterococcus mundtii CWBI-B1431 isolates from artisanal Peruvian cheeses. Ann. Microbiol. 2011, 61, 955–964. [Google Scholar] [CrossRef]
- Han, S.-K.; Shin, M.-S.; Park, H.-E.; Kim, S.-Y.; Lee, W.-K. Screening of bacteriocin-producing Enterococcus faecalis strains for antagonistic activities against Clostridium perfringens. Korean J. Food Sci. Anim. 2014, 34, 614–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yusuf, M.A.; Hamid, T.H.A.T.A. Optimization of temperature and pH for the growth and bacteriocin production of Enterococcus faecium. B3L3. IOSR J. Pharm. 2012, 2, 49–59. [Google Scholar] [CrossRef]
- Leroy, F.; De Vuyst, L. Bacteriocin production by Enterococcus faecium RZS C5 is cell density limited and occurs in the very early growth phase. Int. J. Food Microbiol. 2002, 72, 155–164. [Google Scholar] [CrossRef]
- Todorov, S.D.; Dicks, L.M.T. Optimization of bacteriocin ST311LD production by Enterococcus faecium ST311LD, isolated from spoiled black olives. J. Microbiol. 2005, 43, 370–374. [Google Scholar] [PubMed]
- O’Beirne, D.; Francis, G.A. Reducing pathogen risks in MAP-prepared produce. In Novel Food Packaging Techniques; Woodhead Publishig: Cambridge, UK, 2003; pp. 231–275. [Google Scholar] [CrossRef]
- Gaálvez, A.; Valdivia, E.; Abriouel, H. Isolation and characterisation of enterocin EJ97, a bacteriocin produced by Enterococcus faecalis EJ97. Arch. Microbiol. 1998, 171, 59–65. [Google Scholar] [CrossRef]
- Ivanova, I.; Kabadjova, P.; Pantev, A.; Danova, S.; Dousset, X. Detection, purification and partial characterization of a novel bacteriocin substance produced by Lactococcus lactis subsp. lactis B14 isolated from boza-Bulgarian traditional cereal beverage. Biocatal-Vest. Moskov Univ. Kimia. 2000, 41, 47–53. [Google Scholar]
- Urso, R.; Rantsiou, K.; Cantoni, C.; Comi, G.; Cocolin, L. Technological characterization of a bacteriocin-producing Lactobacillus sakei and its use in fermented sausages production. Int. J. Food Microbiol. 2006, 110, 232–239. [Google Scholar] [CrossRef]
- Abdelsamei, H.; Ibrahim, E.; Saad, M.; Sohaimy, S.E. Effect of storage on the activity of the bacteriocin extracted from Lactobacillus acidophilus. Benha Vet. Med. J. 2015, 28, 216–222. [Google Scholar] [CrossRef] [Green Version]
- Atrih, A.; Rekhif, N.; Moir, A.J.; Lebrihi, A.; Lefebvre, G. Mode of action, purification and amino acid sequence of plantaricin C19, an anti-Listeria bacteriocin produced by Lactobacillus plantarum C19. Int. J. Food Microbiol. 2001, 68, 93–104. [Google Scholar] [CrossRef]
- Sidek, N.L.M.; Halim, M.; Tan, J.S.; Abbasiliasi, S.; Mustafa, S.; Ariff, A.B. Stability of bacteriocin-like inhibitory substance (BLIS) produced by Pediococcus acidilactici kp10 at different extreme conditions. BioMed. Res. Int. 2018, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Sarika, A.R.; Lipton, A.P.; Aishwarya, M.S. Bacteriocin production by a new isolate of Lactobacillus rhamnosus GP1 under different culture conditions. Adv. J. Food Sci. Technol. 2010, 2, 291–297. [Google Scholar]
- Li, J.; Song, D.; Gu, Q. Optimization of plantaricin production by Lactobacillus plantarum ZJ316. Wei Sheng Wu Xue Bao 2008, 48, 818–823. [Google Scholar] [PubMed]
- Parente, E.; Ricciardi, A. Influence of pH on the production of enterocin 1146 during batch fermentation. Lett. Appl. Microbiol. 1994, 19, 12–15. [Google Scholar] [CrossRef] [PubMed]
- Fisher, K.; Phillips, C. The ecology, epidemiology and virulence of Enterococcus. Microbiology 2009, 155, 1749–1757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parente, E.; Brienza, C.; Ricciardi, A.; Addario, G. Growth and bacteriocin production by Enterococcus faecium DPC1146 in batch and continuous culture. J. Ind. Microbiol. Biotechnol. 1997, 18, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Liu, S. Bioprocess Engineering: Kinetics, Sustainability, and Reactor Design, 2nd ed.; Elsevier: Cambridge, MA, USA, 2017; pp. 629–697. [Google Scholar]
- Pongtharangkul, T.; Demirci, A. Evaluation of culture medium for nisin production in a repeated-batch biofilm reactor. Biotechnol. Prog. 2006, 22, 217–224. [Google Scholar] [CrossRef]
- Abbasiliasi, S.; Tan, J.S.; Tengku Ibrahim, T.A.; Bashokouh, F.; Ramakrishnan, N.R.; Mustafa, S.; Ariff, A.B. Fermentation factors influencing the production of bacteriocins by lactic acid bacteria: A review. RSC Adv. 2017, 7, 29395–29420. [Google Scholar] [CrossRef]
- Pingitore, E.V.; Todorov, S.D.; Sesma, F.; Franco, B.D.G.M. Application of bacteriocinogenic Enterococcus mundtii CRL35 and Enterococcus faecium ST88Ch in the control of Listeria monocytogenes in fresh Minas cheese. Food Microbiol. 2012, 32, 38–47. [Google Scholar] [CrossRef]
- Ficoseco, C.A.; Mansilla, F.I.; Maldonado, N.C.; Miranda, H.; Nader-Macias, M.E.F.; Vignolo, G.M. Safety and growth optimization of lactic acid bacteria isolated from feedlot cattle for probiotic formula design. Front. Microbiol. 2018, 9, 2220. [Google Scholar] [CrossRef]
- Murray, B.E. The life and times of the Enterococcus. Clin. Microbiol. Rev. 1990, 3, 46–65. [Google Scholar] [CrossRef]
- Hegstad, K.; Mikalsen, T.; Coque, T.; Werner, G.; Sundsfjord, A. Mobile genetic elements and their contribution to the emergence of antimicrobial resistant Enterococcus faecalis and Enterococcus faecium. Clin. Microbiol. Infec. 2010, 16, 541–554. [Google Scholar] [CrossRef]
- Mühlberg, E.; Umstätter, F.; Kleist, C.; Domhan, C.; Mier, W.; Uhl, P. Renaissance of vancomycin: Approaches for breaking antibiotic resistance in multidrug-resistant bacteria. Can. J. Microbiol. 2019, 66, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Donnenberg, M.S. Escherichia coli: Virulence Mechanisms of a Versatile Pathogen; Academic Press: London, UK, 2002. [Google Scholar]
- Barbieri, F.; Montanari, C.; Gardini, F.; Tabanelli, G. Biogenic amine production by lactic acid bacteria: A Review. Foods 2019, 8, 17. [Google Scholar] [CrossRef] [Green Version]
- Bargossi, E.; Gardini, F.; Gatto, V.; Montanari, C.; Torriani, S.; Tabanelli, G. The capability of tyramine production and correlation between phenotypic and genetic characteristics of Enterococcus faecium and Enterococcus faecalis strains. Front. Microbiol. 2015, 6, 1371. [Google Scholar] [CrossRef] [Green Version]
- Chakchouk-Mtibaa, A.; Sellem, I.; Kamoun, Y.; Smaoui, S.; Karray-Rebai, I.; Mellouli, L. Safety aspect of Enterococcus faecium FL31 strain and antibacterial mechanism of its hydroxylated bacteriocin BacFL31 against Listeria monocytogenes. BioMed Res. Int. 2018, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Todorov, S.D.; Rachman, C.; Fourrier, A.; Dicks, L.M.; van Reenen, C.A.; Prévost, H.; Dousset, X. Characterization of a bacteriocin produced by Lactobacillus sakei R1333 isolated from smoked salmon. Anaerobe 2011, 17, 23–31. [Google Scholar] [CrossRef]
- Stockwell, V.O.; Moore, L.W.; Loper, J.E. Fate of Agrobacterium radiobacter K84 in the environment. Appl. Environ. Microbiol. 1993, 59, 2112–2120. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Lee, M.; Ji, Y.; Todorov, S.D.; Holzapfel, W.H. Safety evaluation and in vivo strain-specific functionality of Bacillus strains isolated from Korean traditional fermented foods. Probiotics Antimicrob. Proteins 2020, in press. [Google Scholar] [CrossRef]
- Montel, M.C.; Masson, F.; Talon, R. Bacterial role in flavour development. Meat Sci. 1998, 49, 111–123. [Google Scholar] [CrossRef]
- Gagne, F. Biochemical Ecotoxicology: Principles and Method; Elsevier Academic Press: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Tian, J.; Lu, Y.; Zhang, H.; Chau, C.H.; Dang, H.N.; Kaufman, D.L. Gamma-aminobutyric acid inhibits T cell autoimmunity and the development of inflammatory responses in a mouse type 1 diabetes model. J. Immunol. 2004, 173, 5298–5304. [Google Scholar] [CrossRef]
- Bhat, R.; Axtell, R.; Mitra, A.; Miranda, M.; Lock, C.; Tsien, R.W.; Steinman, L. Inhibitory role for GABA in autoimmune inflammation. Proc. Nat. Acad. Sci. USA 2010, 107, 2580–2585. [Google Scholar] [CrossRef] [Green Version]
- Kelley, J.M.; Hughes, L.B.; Bridges, S.L., Jr. Does gamma-aminobutyric acid (GABA) influence the development of chronic inflammation in rheumatoid arthritis? J. Neuroinflamm. 2008, 5. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Yong, J.; Dang, H.; Kaufman, D.L. Oral GABA treatment downregulates inflammatory responses in a mouse model of rheumatoid arthritis. Autoimmunity 2011, 44, 465–470. [Google Scholar] [CrossRef] [Green Version]
- Nigam, R.; El-Nour, H.; Amatya, B.; Nordlind, K. GABA and GABA(A) receptor expression on immune cells in psoriasis: A pathophysiological role. Arch. Dermatol. Res. 2010, 302, 507–515. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| µg/mL | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ampicillin | Vancomycin | Gentamicin | Kanamycin | Streptomycin | Erythromycin | Clindamycin | Tylosin | Tetracycline | Chloramphenicol | |
| Breakpoints for E. faecium according to EFSA | 2 | 4 | 32 | 1024 | 128 | 4 | 4 | 4 | 4 | 16 |
| Experimental values for E. faecium ST10Bz | 0.5 | 0.5 | 32 | 128 | 32 | 8 | 0.5 | 1 | 0.5 | 8 |
| Quality control strain range for E. faecalis ATCC®29212TM according CLSI | 0.5–2 | 1–4 | 4–16 | 16–64 | − | 1–4 | 4–16 | - | 8–32 | 4–16 |
| Experimental values for E. faecalis ATCC®29212TM | 1 | 2 | 8 | 16 | 32 | 2 | 16 | 1 | 16 | 8 |
| Sequences | |
|---|---|
| Enterocin B | |
| E. faecium ST10BZ | CHINLFPTPVITFVCCFEN DHRMPNELNR PNNLSKGGAK CGAAIAGGLF GIPKGPLAWA AGLANVYS |
| GenBank: AAD28234.1 | MQNVKELSTKEMKQIIGGEN DHRMPNELNR PNNLSKGGAK CGAAIAGGLF GIPKGPLAWA AGLANVYSKCN |
| Enterocin P | |
| E. faecium ST10BZ | MRKKLFSLTL IGKFGLVVTN FGTKVDAATR SYDNGIYCNN SKCWVNWGEA KENIAGIVIS GWASGLAGM |
| GenBank: ACU28817.1 | MRKKLFSLTL IGKFGLVVTN FGTKVDAATR SYDNGIYCNN SKCWVNWGEA KENIAGIVIS GWASGLAGM |
| Species | Total |
|---|---|
| Enterococcus avium | 1/3 |
| Enterococcus faecium | 6/6 |
| Enterococcus faecalis | 2/2 |
| Enterococcus gallinarum | 0/1 |
| Enterococcus thailandicus | 1/1 |
| Enterococcus durans | 1/1 |
| Vacomycin-resistant Enterococcus strains (VRE) | 30/30 |
| Listeria monocytogenes | 1/1 |
| Listeria innocua | 2/2 |
| Bacillus cereus | 0/2 |
| Bacillus subtilis | 0/1 |
| Bacillus licheniformis | 0/1 |
| Bacillus pumilus | 0/1 |
| Clostridium butyricum | 0/1 |
| Pediococcus acidilactici | 1/1 |
| Pediococcus pentosaceus | 1/4 |
| Weissella cibaria | 0/1 |
| Leuconostoc mesenteroides | 0/6 |
| Staphylococcus aureus | 0/1 |
| Lactobacillus rhamnosus | 0/5 |
| Lactobacillus paracasei | 0/5 |
| Lactobacillus plantarum | 0/9 |
| Lactobacillus sanikiri | 0/4 |
| Lactobacillus brevis | 0/3 |
| Lactobacillus salivarius | 0/3 |
| Lactobacillus sakei | 1/4 |
| Lactobacillus curvatus | 0/2 |
| Lactobacillus fermentum | 0/2 |
| Staphylococcus epidermidis | 0/1 |
| Staphylococcus cohnii subsp. cohnii | 0/1 |
| Staphylococcus warneri | 0/1 |
| Staphylococcus simulans | 0/1 |
| Staphylococcus capitis subsp. capitis | 0/1 |
| Staphylococcus lentus | 0/1 |
| Staphylococcus carnosus subsp. carnosus | 0/1 |
| Staphylococcus auricularis | 0/1 |
| Staphylococcus arlettae | 0/1 |
| Sthaphylococcus delphini | 0/1 |
| Streptococcus sanguinis | 0/1 |
| Streptococcus mitis | 0/1 |
| Streptococcus gordonii | 0/1 |
| Streptococcus mutans | 0/1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valledor, S.J.D.; Bucheli, J.E.V.; Holzapfel, W.H.; Todorov, S.D. Exploring Beneficial Properties of the Bacteriocinogenic Enterococcus faecium ST10Bz Strain Isolated from Boza, a Bulgarian Cereal-Based Beverage. Microorganisms 2020, 8, 1474. https://doi.org/10.3390/microorganisms8101474
Valledor SJD, Bucheli JEV, Holzapfel WH, Todorov SD. Exploring Beneficial Properties of the Bacteriocinogenic Enterococcus faecium ST10Bz Strain Isolated from Boza, a Bulgarian Cereal-Based Beverage. Microorganisms. 2020; 8(10):1474. https://doi.org/10.3390/microorganisms8101474
Chicago/Turabian StyleValledor, Samantha Joy D., Jorge Enrique Vazquez Bucheli, Wilhelm H. Holzapfel, and Svetoslav Dimitrov Todorov. 2020. "Exploring Beneficial Properties of the Bacteriocinogenic Enterococcus faecium ST10Bz Strain Isolated from Boza, a Bulgarian Cereal-Based Beverage" Microorganisms 8, no. 10: 1474. https://doi.org/10.3390/microorganisms8101474
APA StyleValledor, S. J. D., Bucheli, J. E. V., Holzapfel, W. H., & Todorov, S. D. (2020). Exploring Beneficial Properties of the Bacteriocinogenic Enterococcus faecium ST10Bz Strain Isolated from Boza, a Bulgarian Cereal-Based Beverage. Microorganisms, 8(10), 1474. https://doi.org/10.3390/microorganisms8101474