Eradication of Saccharomyces cerevisiae by Pulsed Electric Field Treatments


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Conditions
2.1.1. PBS Solution
2.1.2. Preservative Solutions
2.1.3. The Solutions’ Conductivity and pH
2.2. Design and Construction of the Electroporator
2.2.1. A High-Voltage Generator Was Used for Applying an Electric Field to the S. cerevisiae Suspension
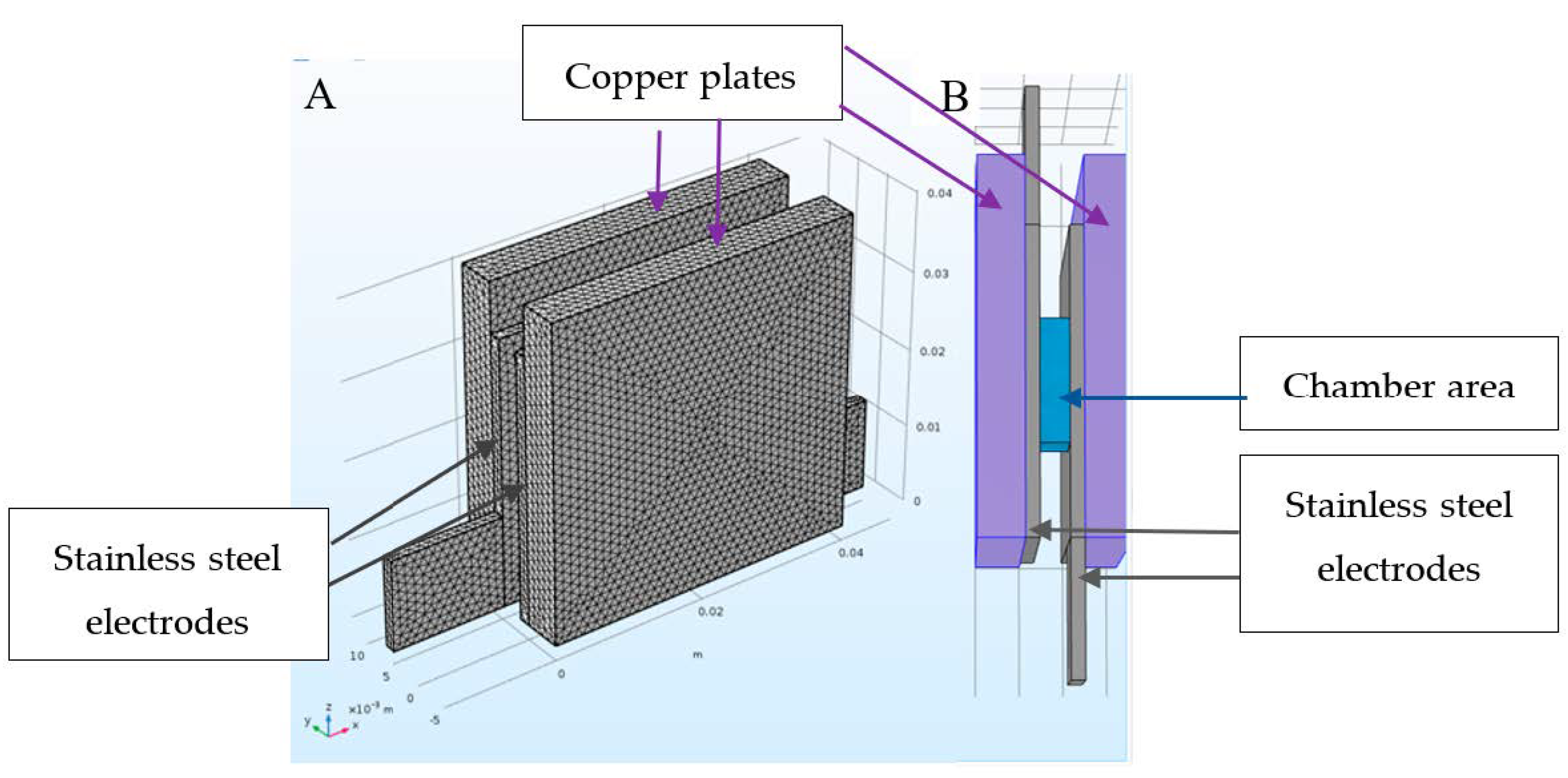
2.2.2. Construction of the Electroporator Chamber
2.3. Characterization of One Cycle of PEF Treatment
2.4. Electro-Pulsation Procedure for Determining the Conditions of Yeast Eradication
2.5. Total Specific Energy
2.6. Electro-Pulsation Procedure for Determination of Membrane Permeability and Viability as a Function of Dilution in PBS and YMB Media
2.7. Examination of Yeast Membrane Permeability Using FCM Analysis
2.8. Statistics
3. Results and Discussion
3.1. The Effect of the Current Density on S. cerevisiae Eradication
3.2. Viability of PEF-Treated S. cerevisiae as a Function of Suspension in YMB Medium vs. PBS
3.3. Membrane Permeability of PEF-Treated S. cerevisiae When Suspended in YMB vs. PBS
3.4. Yeast Eradication as a Function of PEF Treatment-Cycle Mode
3.5. S. cerevisiae Eradication as a Function of Combined PEF Treatment and Preservative
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Schottroff, F.; Fröhling, A.; Zunabovic-Pichler, M.; Krottenthaler, A.; Schlüter, O.; Jäger, H. Sublethal injury and Viable but Non-culturable (VBNC) state in microorganisms during preservation of food and biological materials by non-thermal processes. Front. Microbiol. 2018, 9, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Barba, F.J.; Parniakov, O.; Pereira, S.A.; Wiktor, A.; Grimi, N.; Boussetta, N.; Saraiva, J.A.; Raso, J.; Martin-Belloso, O.; Witrowa-Rajchert, D.; et al. Current applications and new opportunities for the use of pulsed electric fields in food science and industry. Food Res. Int. 2015, 77, 773–798. [Google Scholar] [CrossRef]
- Tylewicz, U.; Tappi, S.; Mannozzi, C.; Romani, S.; Dellarosa, N.; Laghi, L.; Ragni, L.; Rocculi, P.; Dalla Rosa, M. Effect of pulsed electric field (PEF) pre-treatment coupled with osmotic dehydration on physico-chemical characteristics of organic strawberries. J. Food Eng. 2017, 213, 2–9. [Google Scholar] [CrossRef]
- González-arenzana, L.; Reinares, J.P.; López, N.; Santamaría, P.; Garde-cerdán, T.; Gutiérrez, A.R.; López-alfaro, I.; López, R. Impact of Pulsed Electric Field Treatment on Must and Wine Quality. In Handbook of Electroporation; Miklavčič, D., Ed.; Springer: Berlin, Germany, 2017; pp. 2391–2406. ISBN 9783319328867. [Google Scholar]
- Mathys, A.; Reineke, K.; Jäger, H. Editorial: Microbial decontamination by novel technologies -mechanisms and application concepts. Front. Microbiol. 2019, 10, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, O.; Qin, B.L.; Chang, F.J.; Barbosa-Canovas, G.V.; Swanson, B.G. Inactivation of Escherichia Coli in skim milk by high intensity pulsed electric fields. J. Food Process Eng. 1997, 20, 317–336. [Google Scholar] [CrossRef]
- Evrendilek, G.A.; Altuntas, J.; Sangun, M.K.; Zhang, H.Q. Apricot nectar processing by pulsed electric fields. Int. J. Food Prop. 2013, 16, 216–227. [Google Scholar] [CrossRef]
- Sobrino-López, A.; Martin-Belloso, O. Review: Potential of high-intensity pulsed electric field technology for milk processing. Food Eng. Rev. 2009, 2, 17–27. [Google Scholar] [CrossRef]
- Amiali, M.; Ngadi, M.O.; Smith, J.P.; Raghavan, V.G.S. Inactivation of Escherichia coli O157:H7 and Salmonella enteritidis in liquid egg white using pulsed electric field. J. Food Sci. 2006, 71, M88–M94. [Google Scholar] [CrossRef]
- Puertolas, E.; Lopez, N.; Condon, S.; Raso, J.; Alvarez, I. Pulsed electric fields inactivation of wine spoilage yeast and bacteria. Int. J. Food Microbiol. 2009, 130, 49–55. [Google Scholar] [CrossRef]
- Guionet, A.; David, F.; Zaepffel, C.; Coustets, M.; Helmi, K.; Cheype, C.; Packan, D.; Garnier, J.P.; Blanckaert, V.; Teissie, J. E. coli electroeradication on a closed loop circuit by using milli-, micro- and nanosecond pulsed electric fields: Comparison between energy costs. Bioelectrochemistry 2015, 103, 65–73. [Google Scholar] [CrossRef]
- Coustets, M.; Ganeva, V.; Galutzov, B.; Teissie, J. Millisecond duration pulses for flow-through electro-induced protein extraction from E. coli and associated eradication. Bioelectrochemistry 2015, 103, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Sale, A.J.H.; Hamilton, W.A. Effects of high electric fields on microorganisms: I. Killing of bacteria and yeasts. Biochim. Biophys. Acta Gen. Subj. 1967, 148, 781–788. [Google Scholar] [CrossRef]
- Jayaram, S.; Castle, G.S.P.; Margaritis, A. The effects of high field DC pulse and liquid medium conductivity on survivability of Lactobacillus brevis. Appl. Microbiol. Biotechnol. 1993, 40, 117–122. [Google Scholar] [CrossRef]
- Emanuel, E.; Roman, P.; Cahan, R. Influence of the current density in moderate pulsed electric fields on P. putida F1 eradication. Bioelectrochemistry 2019, 126, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Canatella, P.J.; Karr, J.F.; Petros, J.A.; Prausnitz, M.R. Quantitative study of electroporation-mediated molecular uptake and cell viability. Biophys. J. 2001, 80, 755–764. [Google Scholar] [CrossRef] [Green Version]
- French, D.M.; Uhler, M.D.; Gilgenbach, R.M.; Lau, Y.Y. Conductive versus capacitive coupling for cell electroporation with nanosecond pulses. J. Appl. Phys. 2009, 106. [Google Scholar] [CrossRef]
- Khan, S.I.; Blumrosen, G.; Vecchio, D.; Golberg, A.; Mccormack, M.C.; Yarmush, M.L.; Hamblin, M.R.; Austen, W.G. Eradication of multidrug-resistant pseudomonas biofilm with pulsed electric fields. Biotechnol. Bioeng. 2016, 113, 643–650. [Google Scholar] [CrossRef] [Green Version]
- Neumann, E.; Schaefer-Ridder, M.; Wang, Y.; Hofschneider, P.H. Gene transfer into mouse lyoma cells by electroporation in high electric fields. EMBO J. 1982, 1, 841–845. [Google Scholar] [CrossRef]
- Reineke, K.; Schottroff, F.; Meneses, N.; Knorr, D. Sterilization of liquid foods by pulsed electric fields-an innovative ultra-high temperature process. Front. Microbiol. 2015, 6, 400. [Google Scholar] [CrossRef] [Green Version]
- Alkhafaji, S.R.; Farid, M. An investigation on pulsed electric fields technology using new treatment chamber design. Innov. Food Sci. Emerg. Technol. 2007, 8, 205–212. [Google Scholar] [CrossRef]
- Pucihar, G.; Kotnik, T.; Kandušer, M.; Miklavčič, D. The influence of medium conductivity on electropermeabilization and survival of cells in vitro. Bioelectrochemistry 2001, 54, 107–115. [Google Scholar] [CrossRef]
- Baldwin, W.H.; Gregory, B.W.; Osgood, C.J.; Schoenbach, K.H.; Kolb, J.F. Membrane permeability and cell survival after nanosecond pulsed-electric-field exposure-significance of exposure-media composition. IEEE Trans. Plasma Sci. 2010, 38, 2948–2953. [Google Scholar] [CrossRef]
- Kotnik, T.; Frey, W.; Sack, M.; Haberl Meglič, S.; Peterka, M.; Miklavčič, D. Electroporation-based applications in biotechnology. Trends Biotechnol. 2015, 33, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Yarmush, M.L.; Golberg, A.; Serša, G.; Kotnik, T.; Miklavčič, D. Electroporation-based technologies for medicine: Principles, applications, and challenges. Ann. Rev. Biomed. Eng. 2014, 16, 295–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinosita, K.; Tsong, T.Y. Voltage-induced pore formation and hemolysis of human erythrocytes. BBA Biomembr. 1977, 471, 227–242. [Google Scholar] [CrossRef]
- Rols, M.P.; Teissié, J. Electropermeabilization of mammalian cells. Quantitative analysis of the phenomenon. Biophys. J. 1990, 58, 1089–1098. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.; River, L.P.; Pan, F.-S.; Ji, L.; Wollmann, R.L. Surfactant-induced sealing of electropermeabilized skeletal muscle membranes in vivo. Proc. Natl. Acad. Sci. USA 1992, 89, 4524–4528. [Google Scholar] [CrossRef] [Green Version]
- Kramar, P.; Miklavcic, D.; Lebar, A.M. Determination of the lipid bilayer breakdown voltage by means of linear rising signal. Bioelectrochemistry 2007, 70, 23–27. [Google Scholar] [CrossRef]
- Pucihar, G.; Krmelj, J.; Reberšek, M.; Napotnik, T.B.; Miklavčič, D. Equivalent pulse parameters for electroporation. IEEE Trans. Biomed. Eng. 2011, 58, 3279–3288. [Google Scholar] [CrossRef] [Green Version]
- Emerson, J.B.; Adams, R.I.; Román, C.M.B.; Brooks, B.; Coil, D.A.; Dahlhausen, K.; Ganz, H.H.; Hartmann, E.M.; Hsu, T.; Justice, N.B.; et al. Schrödinger’s microbes: Tools for distinguishing the living from the dead in microbial ecosystems. Microbiome 2017, 5. [Google Scholar] [CrossRef]
- Davis, C. Enumeration of probiotic strains: Review of culture-dependent and alternative techniques to quantify viable bacteria. J. Microbiol. Methods 2014, 103, 9–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davey, H.M. Life, death, and in-between: Meanings and methods in microbiology. Appl. Environ. Microbiol. 2011, 77, 5571–5576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somolinos, M.; Mañas, P.; Condón, S.; Pagán, R.; García, D. Recovery of Saccharomyces cerevisiae sublethally injured cells after Pulsed Electric Fields. Int. J. Food Microbiol. 2008, 125, 352–356. [Google Scholar] [CrossRef] [PubMed]
- Marchante, L.; Loarce, L.; Izquierdo-Cañas, P.M.; Alañón, M.E.; García-Romero, E.; Pérez-Coello, M.S.; Díaz-Maroto, M.C. Natural extracts from grape seed and stem by-products in combination with colloidal silver as alternative preservatives to SO2 for white wines: Effects on chemical composition and sensorial properties. Food Res. Int. 2019, 125, 108594. [Google Scholar] [CrossRef]
- Ribéreau-Gayon, P.; Dubourdieu, D.; Donèche, B.; Lonvaud, A. Handbook of Enology, Volume 1: The Microbiology of Wine and Vinifications; John Wiley & Sons: Hoboken, NJ, USA, 2006; Volume 1, ISBN 0470010355. [Google Scholar]
- Santos, M.C.; Nunes, C.; Saraiva, J.A.; Coimbra, M.A. Chemical and physical methodologies for the replacement/reduction of sulfur dioxide use during winemaking: Review of their potentialities and limitations. Eur. Food Res. Technol. 2012, 234, 1–12. [Google Scholar] [CrossRef]
- Sofos, J.N.; Pierson, M.D.; Blocher, J.C.; Busta, F.F. Mode of action of sorbic acid on bacterial cells and spores. Int. J. Food Microbiol. 1986, 3, 1–17. [Google Scholar] [CrossRef]
- Nemes, D.; Kovács, R.; Nagy, F.; Tóth, Z.; Herczegh, P.; Borbás, A.; Kelemen, V.; Pfliegler, W.P.; Rebenku, I.; Hajdu, P.B.; et al. Comparative biocompatibility and antimicrobial studies of sorbic acid derivates. Eur. J. Pharm. Sci. 2020, 143. [Google Scholar] [CrossRef]
- Montanari, C.; Tylewicz, U.; Tabanelli, G.; Berardinelli, A.; Rocculi, P.; Ragni, L.; Gardini, F. Heat-assisted pulsed electric field treatment for the inactivation of Saccharomyces cerevisiae: Effects of the presence of citral. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Sperber, W.H.; Doyle, M.P. Compendium of the Microbiological Spoilage of Foods and Beverages; Springer: New York, NY, USA, 2009. [Google Scholar]
- da Cruz Almeida, E.T.; de Medeiros Barbosa, I.; Tavares, J.F.; Barbosa-Filho, J.M.; Magnani, M.; de Souza, E.L. Inactivation of spoilage yeasts by Mentha spicata L. and M. × villosa Huds. essential oils in cashew, guava, mango, and pineapple juices. Front. Microbiol. 2018, 9, 1111. [Google Scholar] [CrossRef]
- Tabanelli, G.; Montanari, C.; Arioli, S.; Magnani, M.; Patrignani, F.; Lanciotti, R.; Mora, D.; Gardini, F. Physiological response of Saccharomyces cerevisiae to citral combined with thermal treatment. LWT 2019, 101, 827–834. [Google Scholar] [CrossRef]
- Berovic, M.; Berlot, M.; Kralj, S.; Makovec, D. A new method for the rapid separation of magnetized yeast in sparkling wine. Biochem. Eng. J. 2014, 77–84. [Google Scholar] [CrossRef]
- Pretorius, I.S. Tailoring wine yeast for the new millennium: Novel approaches to the ancient art of winemaking. Yeast 2000, 16, 675–729. [Google Scholar] [CrossRef]
- Raso, J.; Frey, W.; Ferrari, G.; Pataro, G.; Knorr, D.; Teissie, J.; Miklavčič, D. Recommendations guidelines on the key information to be reported in studies of application of PEF technology in food and biotechnological processes. Innov. Food Sci. Emerg. Technol. 2016, 37, 312–321. [Google Scholar] [CrossRef] [Green Version]
- Tylewicz, U. How does pulsed electric field work? In Pulsed Electric Fields to Obtain Healthier and Sustainable Food for Tomorrow; Academic Press: Cambridge, MA, USA, 2020; ISBN 9780128164020. [Google Scholar]
- Ou, Q.; Nikolic-Jaric, M. Mechanisms of inactivation of Candida humilis and Saccharomyces cerevisiae by pulsed electric fields. Bioelectrochemistry 2016, 115. [Google Scholar] [CrossRef]
- Timmermans, R.A.H.; Mastwijk, H.C.; Berendsen, L.B.J.M.; Nederhoff, A.L.; Matser, A.M.; Van Boekel, M.A.J.S.; Nierop Groot, M.N. Moderate intensity Pulsed Electric Fields (PEF) as alternative mild preservation technology for fruit juice. Int. J. Food Microbiol. 2019, 298, 63–73. [Google Scholar] [CrossRef]
- Böckmann, R.A.; Grubmüller, H. Multistep binding of divalent cations to phospholipid bilayers: A molecular dynamics study. Angew. Chemie Int. Ed. 2004, 43, 1021–1024. [Google Scholar] [CrossRef]
- Sachs, J.N.; Nanda, H.; Petrache, H.I.; Woolf, T.B. Changes in phosphatidylcholine headgroup tilt and water order induced by monovalent salts: Molecular dynamics simulations. Biophys. J. 2004, 86, 3772–3782. [Google Scholar] [CrossRef] [Green Version]
- Muraji, M.; Tatebe, W.; Berg, H. The influence of extracellular alkali and alkaline-earth ions on electropermeation of Saccharomyces cerevisiae. Bioelectrochem. Bioenerg. 1998, 46, 293–295. [Google Scholar] [CrossRef]
- Paniagua-Martínez, I.; Ramírez-Martínez, A.; Serment-Moreno, V.; Rodrigues, S.; Ozuna, C. Non-thermal technologies as alternative methods for Saccharomyces cerevisiae inactivation in liquid media: A Review. Food Bioprocess Technol. 2018, 11, 487–510. [Google Scholar] [CrossRef]
- Davey, H.M.; Hexley, P. Red but not dead? Membranes of stressed Saccharomyces cerevisiae are permeable to propidium iodide. Environ. Microbiol. 2011, 13, 163–171. [Google Scholar] [CrossRef]
- El Zakhem, H.; Lanoisellé, J.L.; Lebovka, N.I.; Nonus, M.; Vorobiev, E. The early stages of Saccharomyces cerevisiae yeast suspensions damage in moderate pulsed electric fields. Colloids Surfaces B Biointerfaces 2006, 47, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Rems, L.; Viano, M.; Kasimova, M.A.; Miklavčič, D.; Tarek, M. The contribution of lipid peroxidation to membrane permeability in electropermeabilization: A molecular dynamics study. Bioelectrochemistry 2019, 125, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Breton, M.; Mir, L.M. Investigation of the chemical mechanisms involved in the electropulsation of membranes at the molecular level. Bioelectrochemistry 2018, 119, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Stirke, A.; Celiesiute-Germaniene, R.; Zimkus, A.; Zurauskiene, N.; Simonis, P.; Dervinis, A.; Ramanavicius, A.; Balevicius, S. The link between yeast cell wall porosity and plasma membrane permeability after PEF treatment. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Somolinos, M.; García, D.; Condón, S.; Mañas, P.; Pagán, R. Relationship between sublethal injury and inactivation of yeast cells by the combination of sorbic acid and pulsed electric fields. Appl. Environ. Microbiol. 2007, 73, 3814–3821. [Google Scholar] [CrossRef] [Green Version]



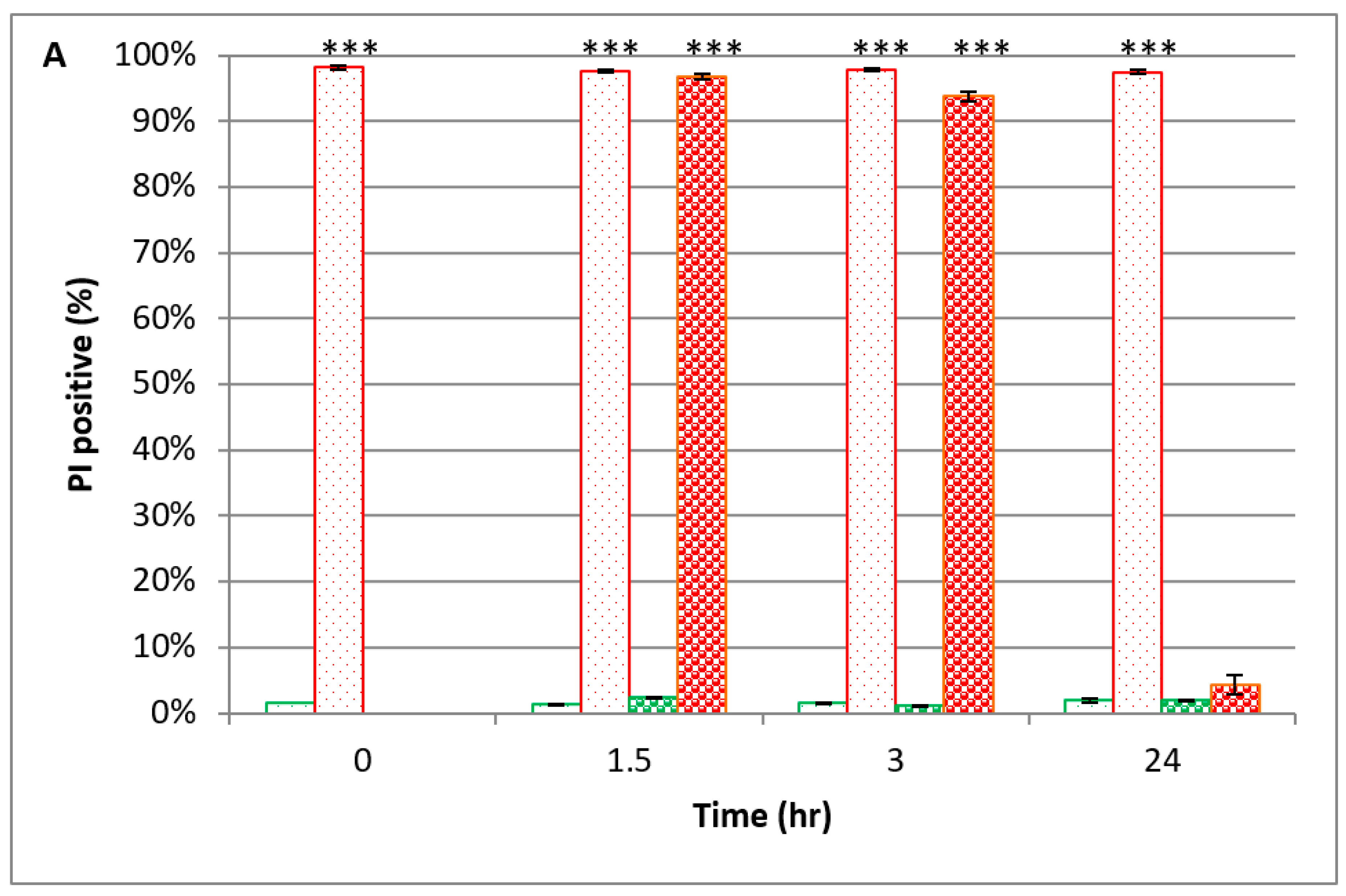
 ); nontreated yeast in time ‘0’ (
); nontreated yeast in time ‘0’ (  ); PEF-treated yeast in YMB (1–24 h) (
); PEF-treated yeast in YMB (1–24 h) (  ); PEF-treated yeast in PBS (1–24 h) (
); PEF-treated yeast in PBS (1–24 h) (  ); nontreated yeast in YMB (1–24 h) (
); nontreated yeast in YMB (1–24 h) (  ); nontreated yeast in PBS (1–24 h) (
); nontreated yeast in PBS (1–24 h) (  ). P value (t test): significance of the CFU count in each examined time related to its control *** p < 0.001; significance of the CFU of the treated yeast in PBS related to YMB, in each examined time ### p < 0.001.
); nontreated yeast in time ‘0’ ( ); PEF-treated yeast in YMB (1–24 h) ( ); PEF-treated yeast in PBS (1–24 h) ( ); nontreated yeast in YMB (1–24 h) ( ); nontreated yeast in PBS (1–24 h) ( ). P value (t test): significance of the CFU count in each examined time related to its control *** p < 0.001; significance of the CFU of the treated yeast in PBS related to YMB, in each examined time ### p < 0.001.
). P value (t test): significance of the CFU count in each examined time related to its control *** p < 0.001; significance of the CFU of the treated yeast in PBS related to YMB, in each examined time ### p < 0.001.
); nontreated yeast in time ‘0’ ( ); PEF-treated yeast in YMB (1–24 h) ( ); PEF-treated yeast in PBS (1–24 h) ( ); nontreated yeast in YMB (1–24 h) ( ); nontreated yeast in PBS (1–24 h) ( ). P value (t test): significance of the CFU count in each examined time related to its control *** p < 0.001; significance of the CFU of the treated yeast in PBS related to YMB, in each examined time ### p < 0.001.
 ); PEF-treated yeasts, which were suspended in PBS (0–24 h) (
); PEF-treated yeasts, which were suspended in PBS (0–24 h) (  ); nontreated yeasts, which were suspended in YMB (1.5–24 h) (
); nontreated yeasts, which were suspended in YMB (1.5–24 h) (  ); nontreated yeasts, which were suspended in PBS (0–24 h) (
); nontreated yeasts, which were suspended in PBS (0–24 h) (  ). P value (t test): *** p < 0.001.
); PEF-treated yeasts, which were suspended in PBS (0–24 h) ( ); nontreated yeasts, which were suspended in YMB (1.5–24 h) ( ); nontreated yeasts, which were suspended in PBS (0–24 h) ( ). P value (t test): *** p < 0.001.
). P value (t test): *** p < 0.001.
); PEF-treated yeasts, which were suspended in PBS (0–24 h) ( ); nontreated yeasts, which were suspended in YMB (1.5–24 h) ( ); nontreated yeasts, which were suspended in PBS (0–24 h) ( ). P value (t test): *** p < 0.001.

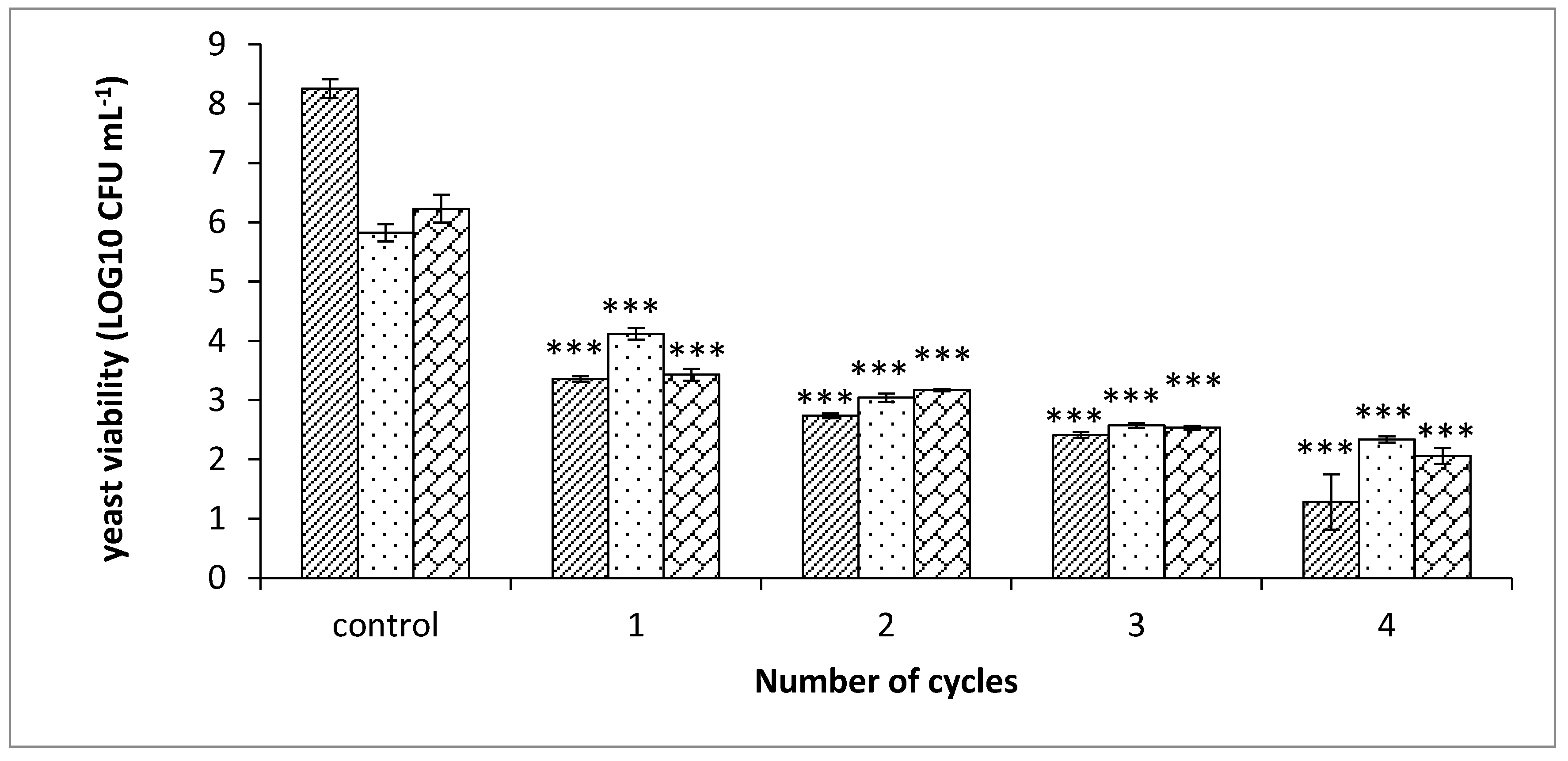
 ); added 25 ppm free SO2 and 25 ppm sorbic acid (
); added 25 ppm free SO2 and 25 ppm sorbic acid (  ); added 50 ppm free SO2 and 125 ppm sorbic acid (
); added 50 ppm free SO2 and 125 ppm sorbic acid (  ). The conditions of each PEF cycle: electric field intensity 2.9 kV cm−1, frequency 100 Hz, pulse duration 10 µs, conductivity 1200 µS cm−1. p value (t test): *** p < 0.001.
); added 25 ppm free SO2 and 25 ppm sorbic acid ( ); added 50 ppm free SO2 and 125 ppm sorbic acid ( ). The conditions of each PEF cycle: electric field intensity 2.9 kV cm−1, frequency 100 Hz, pulse duration 10 µs, conductivity 1200 µS cm−1. p value (t test): *** p < 0.001.
). The conditions of each PEF cycle: electric field intensity 2.9 kV cm−1, frequency 100 Hz, pulse duration 10 µs, conductivity 1200 µS cm−1. p value (t test): *** p < 0.001.
); added 25 ppm free SO2 and 25 ppm sorbic acid ( ); added 50 ppm free SO2 and 125 ppm sorbic acid ( ). The conditions of each PEF cycle: electric field intensity 2.9 kV cm−1, frequency 100 Hz, pulse duration 10 µs, conductivity 1200 µS cm−1. p value (t test): *** p < 0.001.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| j (A cm−2) | λ0 (µS cm−1) | WT (kJ kg−1) | ∆Tav (°C) |
|---|---|---|---|
| control | 1 | -------- | -------- |
| 0.02 | 1 | 22.04 | 0 ± 0.1 |
| 0.6 | 155 | 135.06 | ND |
| 0.88 | 330 | 136.38 | ND |
| 1.6 | 540 | 275.74 | 6 ± 0.1 |
| 3.3 | 1030 | 614.59 | 9 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Emanuel, E.; Dubrovin, I.; Hanya, E.; Pinhasi, G.A.; Pogreb, R.; Cahan, R. Eradication of Saccharomyces cerevisiae by Pulsed Electric Field Treatments. Microorganisms 2020, 8, 1684. https://doi.org/10.3390/microorganisms8111684
Emanuel E, Dubrovin I, Hanya E, Pinhasi GA, Pogreb R, Cahan R. Eradication of Saccharomyces cerevisiae by Pulsed Electric Field Treatments. Microorganisms. 2020; 8(11):1684. https://doi.org/10.3390/microorganisms8111684
Chicago/Turabian StyleEmanuel, Efrat, Irina Dubrovin, Ester Hanya, Gad A. Pinhasi, Roman Pogreb, and Rivka Cahan. 2020. "Eradication of Saccharomyces cerevisiae by Pulsed Electric Field Treatments" Microorganisms 8, no. 11: 1684. https://doi.org/10.3390/microorganisms8111684
APA StyleEmanuel, E., Dubrovin, I., Hanya, E., Pinhasi, G. A., Pogreb, R., & Cahan, R. (2020). Eradication of Saccharomyces cerevisiae by Pulsed Electric Field Treatments. Microorganisms, 8(11), 1684. https://doi.org/10.3390/microorganisms8111684