Seasonal Changes in Microbial Dissolved Organic Sulfur Transformations in Coastal Waters


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Organic Sulfur Concentrations
2.2. Stable Isotope Tracer Rate Experiments
2.3. Radiotracer Rate Experiments
2.3.1. Carbon Assimilation from DMSOd
2.3.2. Carbon Dissimilation to CO2 from DMSOd
3. Results
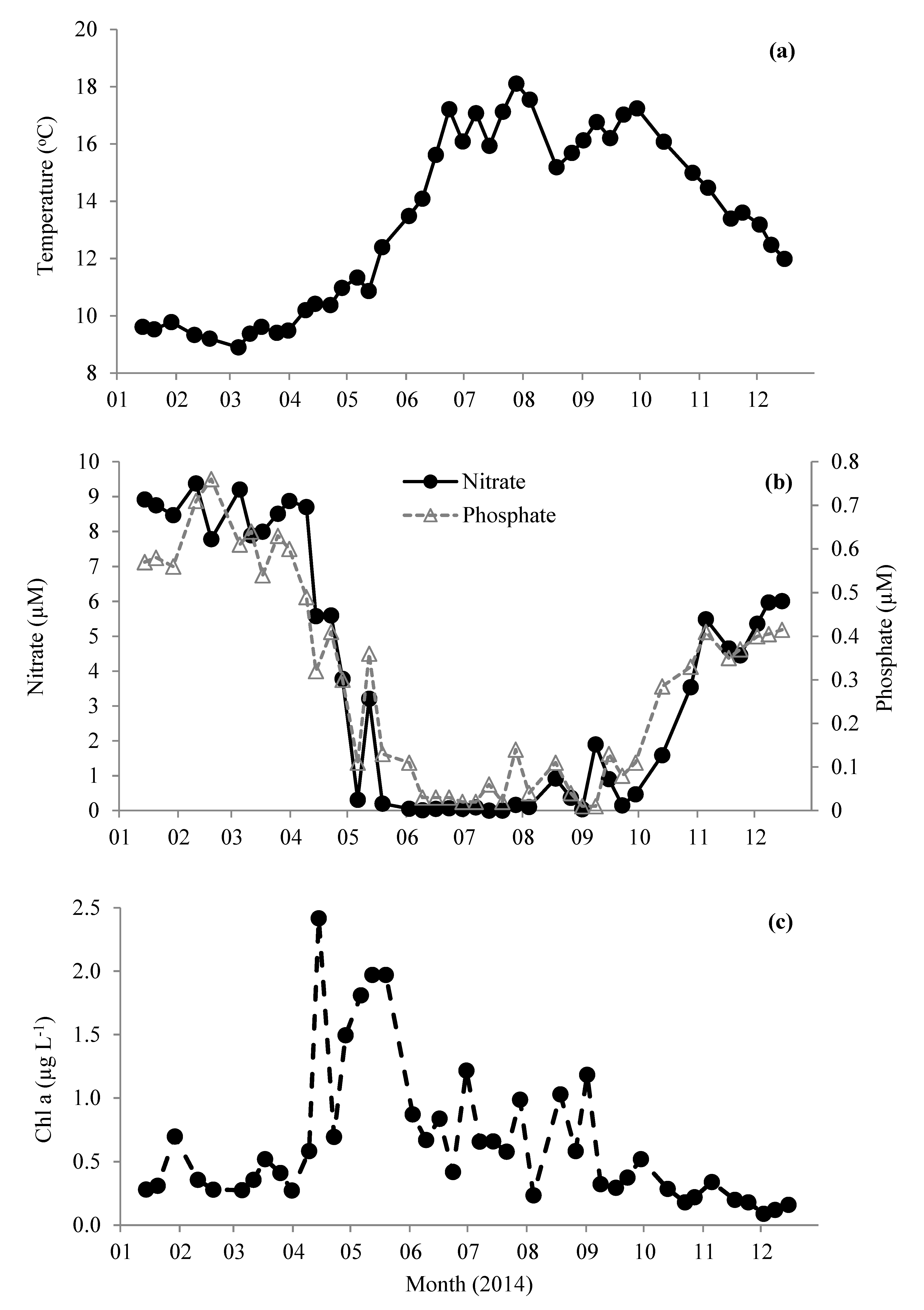
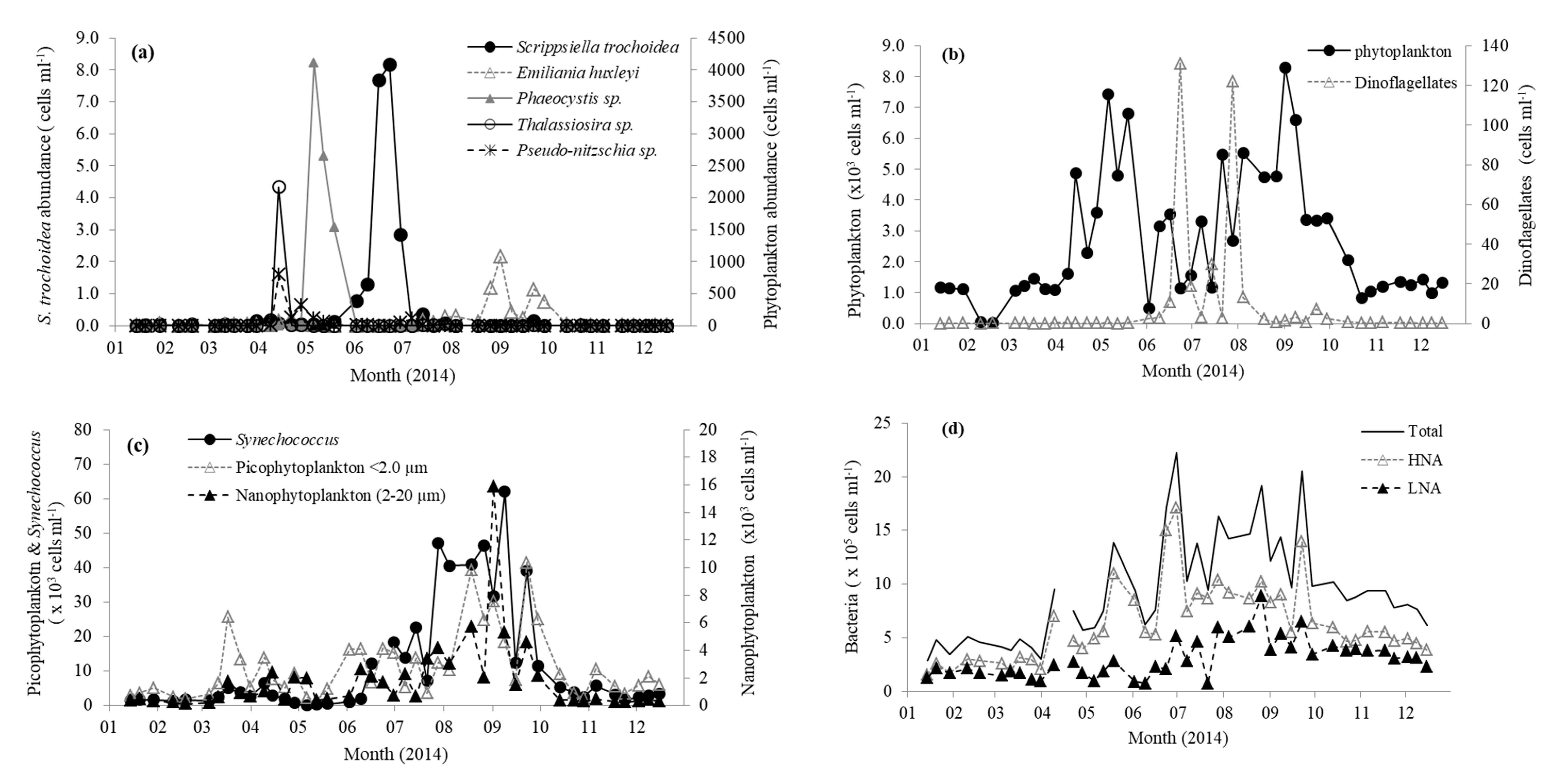
3.1. Environmental and Biological Variables at Station L4
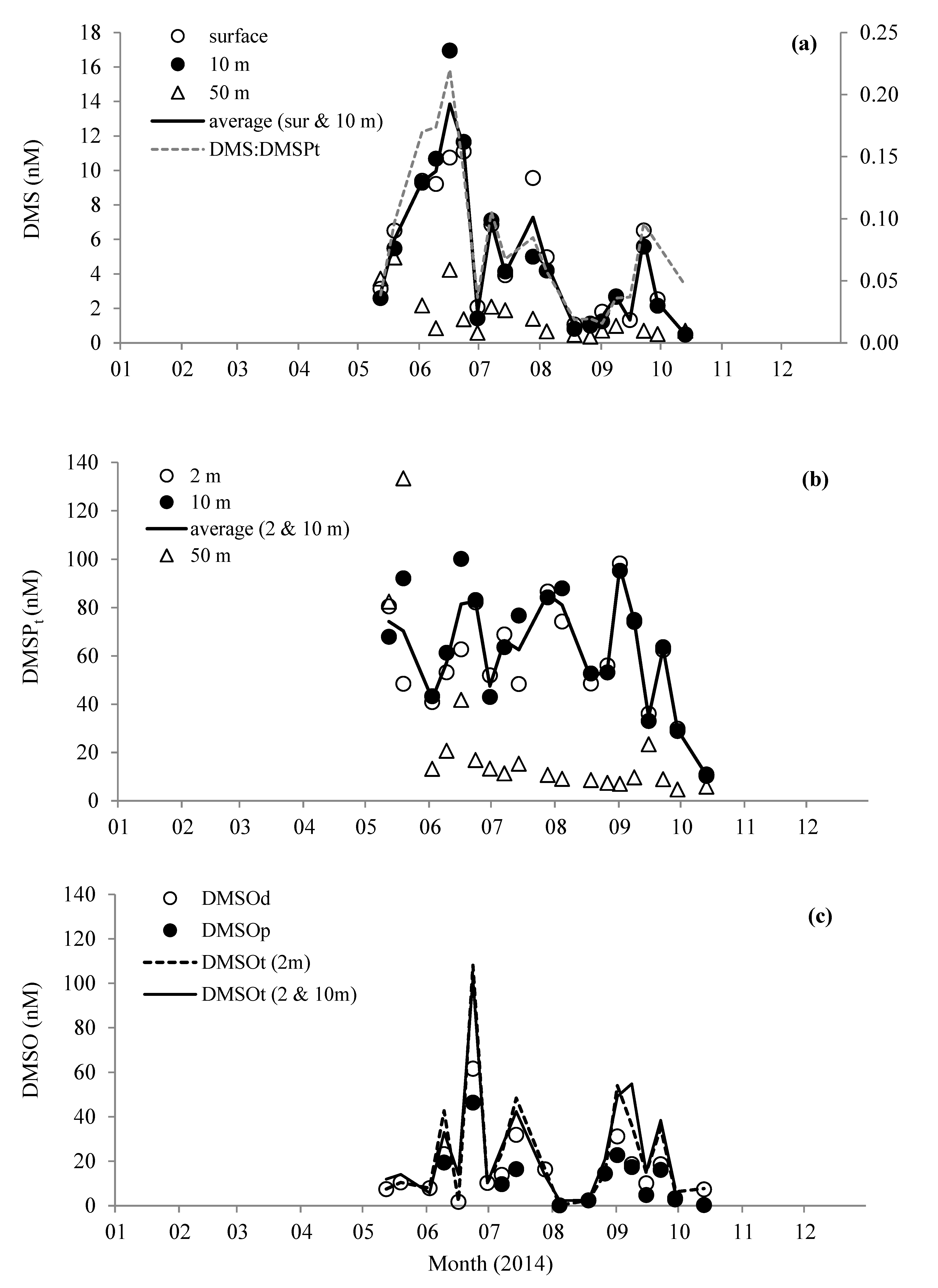
3.2. DMS, DMSP, and DMSO Concentrations
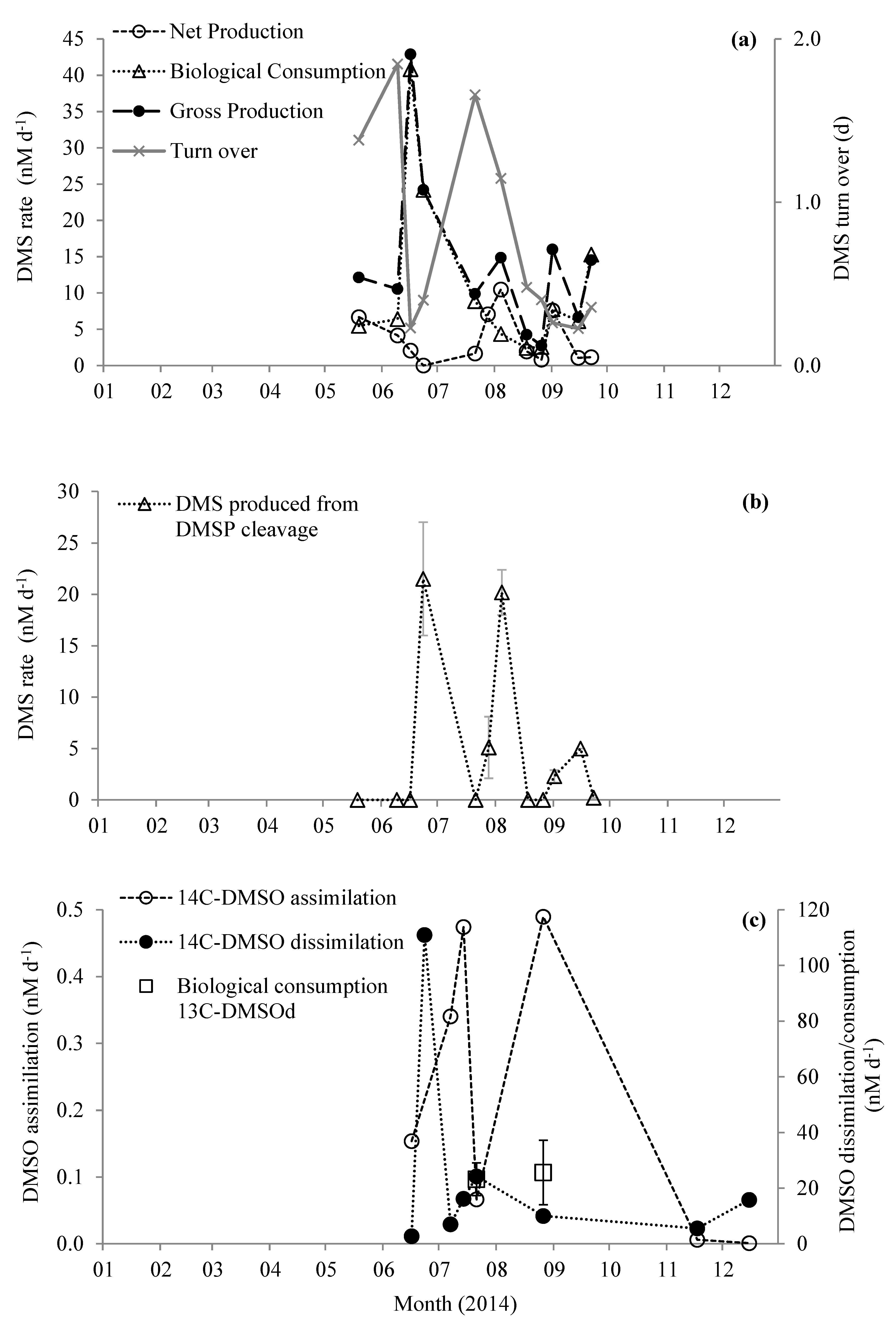
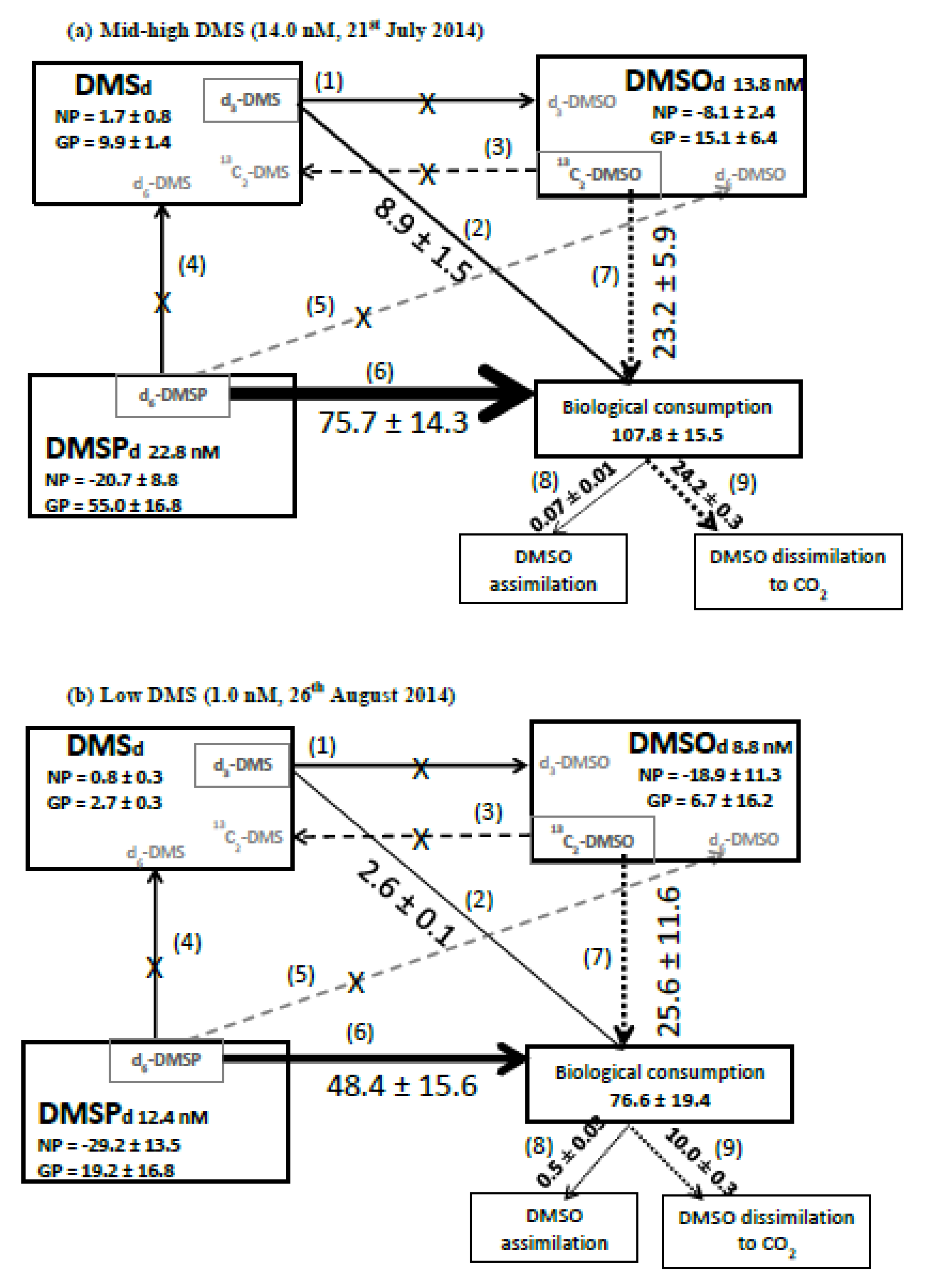
3.3. Stable-Isotope Tracer Experiments
3.4. Radiotracer Experiments
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Levasseur, M. Impact of Arctic meltdown on the microbial cycling of sulfur. Nat. Geosci. 2013, 6, 691–700. [Google Scholar] [CrossRef]
- Lovelock, J.E.; Maggs, R.J.; Rasmussen, R.A. Atmospheric dimethyl sulphide and the natural sulphur cycle. Nature 1972, 237, 452–453. [Google Scholar] [CrossRef]
- Kettle, A.J.; Andreae, M.O. Flux of dimethylsulfide from the oceans: A comparison of updated datasets and flux models. J. Geophys. Res. Atmos. 2000, 105, 26793–26808. [Google Scholar] [CrossRef]
- Curson, A.R.; Todd, J.D.; Sullivan, M.; Johnston, A.W. Catabolism of dimethylsulphoniopropionate: Microorganisms, enzymes and genes. Nat. Rev. Microbiol. 2011, 9, 849–859. [Google Scholar] [CrossRef] [PubMed]
- Galí, M.; Simó, R. A meta-analysis of oceanic DMS and DMSP cycling processes: Disentangling the summer paradox. Glob. Biogeochem. Cycles 2015, 29, 496–515. [Google Scholar] [CrossRef]
- Kiene, R.P.; Bates, T.S. Biological removal of dimethyl sulphide from sea water. Nature 1990, 345, 702–705. [Google Scholar] [CrossRef]
- Herrmann, M.; Najjar, R.G.; Neeley, A.R.; Vila-Costa, M.; Dacey, J.W.; DiTullio, G.R.; Kieber, D.J.; Kiene, R.P.; Matrai, P.A.; Simo, R.; et al. Diagnostic modeling of dimethylsulfide production in coastal water west of the Antarctic Peninsula. Cont. Shelf Res. 2012, 32, 96–109. [Google Scholar] [CrossRef] [Green Version]
- Andreae, M.O. Ocean-atmosphere interactions in the global biogeochemical sulfur cycle. Mar. Chem. 1990, 30, 1–29. [Google Scholar] [CrossRef]
- Von Glasow, R.V.; Crutzen, P.J. Model study of multiphase DMS oxidation with a focus on halogens. Atmos. Chem. Phys. 2004, 4, 589–608. [Google Scholar] [CrossRef] [Green Version]
- Kiene, R.P.; Linn, L.J. The fate of dissolved dimethylsulfoniopropionate (DMSP) in seawater: Tracer studies using 35S-DMSP. Geochim. Cosmochim. Acta 2000, 64, 2797–2810. [Google Scholar] [CrossRef]
- Simó, R.; Archer, S.D.; Pedrós-Alió, C.; Gilpin, L.; Stelfox-Widdicombe, C.E. Coupled dynamics of dimethylsulfoniopropionate and dimethylsulfide cycling and the microbial food web in surface waters of the North Atlantic. Limnol. Oceanogr. 2002, 47, 3–61. [Google Scholar] [CrossRef] [Green Version]
- Vila-Costa, M.; Kiene, R.P.; Simó, R. Seasonal variability of the dynamics of dimethylated sulfur compounds in a coastal northwest Mediterranean site. Limnol. Oceanogr. 2008, 53, 198–211. [Google Scholar] [CrossRef] [Green Version]
- del Valle, D.A.; Kieber, D.J.; Toole, D.A.; Brinkley, J.; Kienea, R.P. Biological consumption of dimethylsulfide (DMS) and its importance in DMS dynamics in the Ross Sea, Antarctica. Limnol. Oceanogr. 2009, 54, 785–798. [Google Scholar] [CrossRef]
- Del Valle, D.A.; Kieber, D.J.; Kiene, R.P. Depth-dependent fate of biologically consumed dimethylsulfide in the Sargasso Sea. Mar. Chem. 2007, 103, 197–208. [Google Scholar] [CrossRef]
- Asher, E.C.; Dacey, J.W.H.; Mills, M.M.; Arrigo, K.R.; Tortell, P.D. High concentrations and turnover rates of DMS, DMSP and DMSO in Antarctic sea ice. Geophys. Res. Letts. 2011, 38, L23609. [Google Scholar] [CrossRef] [Green Version]
- Steinke, M.; Daniel, C.; Kirst, G.O. DMSP Lyase in Marine Macro- and Microalgae. In Biological and Environmental Chemistry of DMSP and Related Sulfonium Compounds; Kiene, R.P., Visscher, P.T., Keller, M.D., Kirst, G.O., Eds.; Springer: Boston, MA, USA, 1996; pp. 317–324. [Google Scholar]
- Stefels, J.; van Boekel, W.H.M. Production of DMS from dissolved DMSP in axenic cultures of the marine phytoplankton species Phaeocystis sp. Mar. Ecol. Prog. Ser. 1993, 97, 11–18. [Google Scholar] [CrossRef]
- Spiese, C.E. Cellular Production and Losses of Dimethylsulfide in Marine Phytoplankton. Ph.D. Thesis, State University of New York College of Environmental Science and Forestry, Syracuse, NY, USA, 2010. [Google Scholar]
- González, J.M.; Kiene, R.P.; Moran, M.A. Transformation of sulphur compounds by an abundant lineage of the marine bacteria in the α-subclass of the class Proteobacteria. Appl. Environ. Microbiol. 1999, 65, 3810–3819. [Google Scholar] [CrossRef] [Green Version]
- Asher, E.C.; Dacey, J.W.H.; Stukel, M.; Long, M.C.; Tortell, P.D. Processes driving seasonal variability in DMS, DMSP, and DMSO concentrations and turnover in coastal Antarctic waters. Limnol. Oceanogr. 2017, 62, 104–124. [Google Scholar] [CrossRef]
- Stefels, J. Physiological aspects of the production and conversion of DMSP in marine algae and higher plants. J. Sea Res. 2000, 43, 183–197. [Google Scholar] [CrossRef]
- Simó, R. Production of atmospheric sulfur by oceanic plankton: Biogeochemical, ecological and evolutionary links. Trends Ecol. Evol. 2001, 16, 287–294. [Google Scholar] [CrossRef]
- Simό, R.; Grimalt, J.O.; Albaigés, J. Dissolved dimethylsulphide, dimethylsulphoniopropionate and dimethylsulphoxide in western Mediterranean waters. Deep Sea Res. II 1997, 44, 929–950. [Google Scholar] [CrossRef]
- Zindler, C.; Bracher, A.; Marandino, C.A.; Taylor, B.; Torrecilla, E.; Kock, A.; Bange, H.W. Sulphur compounds, methane, and phytoplankton: Interactions along a north-south transit in the western Pacific Ocean. Biogeosciences 2013, 10, 3297–3311. [Google Scholar] [CrossRef] [Green Version]
- Lee, P.A.; de Mora, S.J. Intracellular dimethylsulphoxide (DMSO) in unicellular marine algae: Speculations on its origin and possible biological role. J. Phycol. 1999, 35, 8–18. [Google Scholar]
- Chen, Y.; Patel, N.A.; Crombie, A.; Scrivens, J.H.; Murrell, J.C. Bacterial flavin-containing monooxygenase is trimethylamine monooxygenase. Proc. Natl. Acad. Sci. USA 2011, 108, 17791–17796. [Google Scholar] [CrossRef] [Green Version]
- Lidbury, I.; Kröber, E.; Zhang, Z.; Zhu, Y.; Murrell, J.C.; Chen, Y.; Schäfer, H. A mechanism for bacterial transformation of dimethylsulfide to dimethylsulfoxide: A missing link in the marine organic sulfur cycle. Environ. Microb. 2016, 18, 2754–2766. [Google Scholar] [CrossRef] [Green Version]
- Andreae, M.O. Dimethylsulfoxide in marine and freshwaters. Limnol. Oceanogr. 1980, 25, 1054–1063. [Google Scholar] [CrossRef]
- Sunda, W.; Kieber, D.J.; Kiene, R.P.; Huntsman, S. An antioxidant function for DMSP and DMS in marine algae. Nature 2002, 418, 317–320. [Google Scholar] [CrossRef]
- Simό, R.; Pedrόs-Aliό, C.; Malin, G.; Grimalt, J.O. Biological turnover of DMS, DMSP and DMSO in contrasting open-sea waters. Mar. Ecol. Prog. Ser. 2000, 203, 1–11. [Google Scholar] [CrossRef]
- Tyssebotn, I.M.B.; Kinsey, J.D.; Kieber, D.J.; Kiene, R.P.; Rellinger, A.N.; Motard-Côté, J. Concentrations, biological uptake, and respiration of dissolved acrylate and dimethylsulfoxide in the northern Gulf of Mexico. Limnol. Oceanogr. 2017, 62, 1198–1218. [Google Scholar] [CrossRef]
- Hatton, A.D.; Darroch, L.; Malin, G. The role of dimethylsulphoxide in the marine biogeochemical cycle of dimethylsuphide. Oceanogr. Mar. Biol. Annu. Rev. 2004, 42, 29–56. [Google Scholar]
- Spiese, C.E.; Kieber, D.J.; Nomura, C.T.; Kiene, R.P. Reduction of dimethylsulfoxide to dimethylsulfide by marine phytoplankton. Limnol. Oceanogr. 2009, 54, 560–570. [Google Scholar] [CrossRef]
- McCrindle, S.L.; Kappler, U.; McEwan, A.G. Microbial dimethylsulfoxide and trimethylamine-N-oxide respiration. Adv. Microb. Physiol. 2005, 50, 147–198. [Google Scholar] [PubMed]
- Vogt, C.; Rabenstein, A.; Rethmeier, J.; Fischer, U. Dimethyl sulphoxide reduction with reduced sulphur compounds as electron donors by anoxygenic phototrophic bacteria. Microbiology 1997, 143, 767–773. [Google Scholar] [CrossRef] [Green Version]
- deBont, J.A.M.; van Dijken, J.P.; Harder, W. Dimethyl sulphoxide and dimethyl sulfide as a carbon, sulphur and energy source for growth of Hyphomicrobium. S. J. Gen. Microbiol. 1981, 127, 315–323. [Google Scholar]
- Borodina, E.; Kelly, D.P.; Rainey, F.A.; Ward-Rainey, N.L.; Wood, A.P. Dimethylsulfone as a growth substrate for novel methylotrophic species of Hyphomicrobium and Arthrobacter. Arch. Microbiol. 2000, 173, 425–437. [Google Scholar] [CrossRef]
- Kim, H.G.; Doronina, N.V.; Trostsenko, Y.A.; Kim, S.W. Methylophaga aminisulfidivorans sp. nov., a restricted facultatively methylotrophic marine bacterium. Int. J. Evol. Syst. Microbiol. 2007, 57, 2096–2101. [Google Scholar] [CrossRef]
- Smyth, T.J.; Fishwick, J.R.; Al-Moosawi, L.; Cummings, D.G.; Harris, C.; Kitidis, V.; Rees, A.; Martinez-Vicente, V.; Woodward, E.M.S.W. A broad spatio-temporal view of the Western English Channel observatory. J. Plankton Res. 2010, 32, 585–601. [Google Scholar] [CrossRef] [Green Version]
- Tilstone, G.; Smyth, T.A.P.; Hutson, R. Measured and remotely sensed estimates of primary production in the Atlantic Ocean from 1998 to 2005. Deep Sea Res. II 2009, 56, 918–930. [Google Scholar] [CrossRef]
- Brewer, P.G.; Riley, J.P. The automatic determination of nitrate in sea water. Deep Sea Res. 1965, 12, 765–772. [Google Scholar] [CrossRef]
- Grasshoff, H. Methods of Seawater Analysis; Chemie: Weinheim, Germany; New York, NY, USA, 1976; p. 317. [Google Scholar]
- Zhang, J.Z.; Chi, J. Automated analysis of nanomolar concentrations of phosphate in natural waters with liquid waveguides. Environ. Sci. Technol. 2002, 36, 1048–1053. [Google Scholar] [CrossRef]
- Tarran, G.A.; Bruun, J.T. Nanoplankton and picoplankton in the Western English Channel: Abundance and seasonality from 2007–2013. Prog. Oceanogr. 2015, 137, 446–455. [Google Scholar] [CrossRef]
- Widdicombe, C.E.; Eloire, D.; Harbour, D.; Harris, R.P.; Somerfield, P.J. Long-term phytoplankton community dynamics in the Western English Channel. J. Plankton Res. 2010, 32, 643–655. [Google Scholar] [CrossRef] [Green Version]
- Simό, R.; Grimalt, J.O.; Albaigés, J. Sequential method for the determination of nanomolar concentrations of dimethyl sulfoxide in natural waters. Anal. Chem. 1996, 68, 1493–1498. [Google Scholar] [CrossRef]
- Simό, R.; Vila-Costa, M. Ubiquity of algal dimethylsulfoxide in the surface ocean: Geographic and temporal distribution patterns. Mar. Chem. 2006, 100, 136–146. [Google Scholar] [CrossRef]
- Archer, S.D.; Kimmance, S.A.; Stephens, J.A.; Hopkins, F.E.; Bellerby, R.G.J.; Schulz, K.G.; Piontek, J.; Engel, A. Contrasting responses of DMS and DMSP to ocean acidification in Arctic waters. Biogeosciences 2013, 10, 1893–1908. [Google Scholar] [CrossRef] [Green Version]
- Townsend, D.W.; Keller, M.D. Dimethylsulfide (DMS) and dimethylsulfoniopropionate (DMSP) in relation to phytoplankton in the Gulf of Maine. Mar. Ecol. Prog. Ser. 1996, 137, 229–241. [Google Scholar] [CrossRef]
- Simό, R.; Malin, G.; Liss, P. Refinement of the borohydride reduction method for trace analysis of dissolved and particulate dimethyl sulfoxide in marine water samples. Anal. Chem. 1998, 70, 4864–4867. [Google Scholar] [CrossRef]
- Kinsey, J.D.; Kieber, D.J. Microwave preservation method for DMSP, DMSO and acrylate in unfiltered seawater and phytoplankton culture samples. Limnol. Oceanogr. Methods 2016, 14, 196–209. [Google Scholar] [CrossRef]
- Kiene, R.P.; Hoffmann Williams, L.P. Glycine betaine uptake, retention, and degradation by microorganisms in seawater. Limnol. Oceanogr. 1998, 43, 1592–1603. [Google Scholar] [CrossRef]
- Kiene, R.P.; Slezak, D. Low dissolved DMSP concentrations in seawater revealed by small-volume gravity filtration and dialysis sampling. Limnol. Oceanogr: Methods 2006, 4, 80–95. [Google Scholar] [CrossRef]
- Goodwin, K.D.; Schaefer, J.K.; Oremland, R.S. Bacterial oxidation of dibromomethane and methyl bromide in natural waters and enrichement cultures. Appl. Environ. Microbiol. 1998, 64, 4629–4636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archer, S.D.; Cummings, D.G.; Llewellyn, C.A.; Fishwick, J.R. Phytoplankton taxa, irradiance and nutrient availability determine the seasonal cycle of DMSP in temperate shelf seas. Mar. Ecol. Prog. Ser. 2009, 394, 111–124. [Google Scholar] [CrossRef]
- Zindler, C.; Peeken, I.; Marandino, C.A.; Bange, H.W. Environmental control on the variability of DMS and DMSP in the Mauritanian upwelling region. Biogosciences 2012, 9, 1041–1051. [Google Scholar] [CrossRef] [Green Version]
- Niki, T.; Kunugi, M.; Otsuki, A. DMSP-lyase activity in five marine phytoplankton species: Its potential importance in DMS production. Mar. Biol. 2000, 136, 759–764. [Google Scholar] [CrossRef]
- Caruana, A.M.N.; Steinke, M.; Turner, S.M.; Malin, G. Concentrations of dimethylsulphoniopropionate and activities of dimethylsulphide-producing enzymes in batch cultures of nine dinoflagellate species. Biogeochemistry 2012, 110, 87–107. [Google Scholar] [CrossRef]
- Steinke, M.; Malin, G.; Archer, S.D.; Burkill, P.H.; Liss, P.S. DMS production in a coccolithophorid bloom: Evidence for the importance of dinoflagellate DMSP lyases. Aquat. Microb. Ecol. 2002, 26, 259–270. [Google Scholar] [CrossRef] [Green Version]
- Franklin, D.J.; Poulton, A.J.; Steinke, M.; Young, J.; Peeken, I.; Malin, G. Dimethylsulphide, DMSP-lyase activity and microplankton community structure inside and outside of the Mauritanian upwelling. Prog. Oceanogr. 2009, 83, 134–142. [Google Scholar] [CrossRef]
- Michaud, S.; Levasseur, M.; Cantin, G. Seasonal variations in dimethylsulfoniopropionate and dimethylsulfide concentrations in relation to the plankton community in the St. Lawrence Estuary. Estuar. Coast. Shelf Sci. 2007, 71, 741–750. [Google Scholar] [CrossRef]
- Speeckaert, G.; Borges, A.V.; Champenois, W.; Royer, C.; Gypens, N. Annual cycle of dimethylsulfoniopropionate (DMSP) and dimethylsulfoxide (DMSO) related to phytoplankton succession in the Southern North Sea. Sci. Total Environ. 2018, 622, 362–372. [Google Scholar] [CrossRef] [Green Version]
- Boden, R.; Murrell, J.C.; Schäfer, H. Dimethylsulfide is an energy source for the heterotrophic marine bacterium Sagittula stellata. FEMS Microbiol. Lett. 2011, 322, 188–193. [Google Scholar] [CrossRef]
- Cui, Y.; Suzuki, S.; Omori, Y.; Wong, A.-K.; Ijichi, M.; Kaneko, R.; Kameyama, S.; Tanimoto, H.; Hamasaki, K. Abundance and distribution of dimethylsulfoniopropionate degradation genes and the corresponding bacterial community structure are dimethyl sulfide hot spots in the tropical and subtropical Pacific Ocean. Appl. Environ. Microbiol. 2015, 81, 4184–4194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newton, R.J.; Griffin, L.E.; Bowles, K.M.; Meile, C.; Gifford, S.; Givens, C.E.; Howard, E.C.; King, E.; Oakley, C.A.; Reisch, C.Y.; et al. Genome characteristics of a generalist marine bacterial lineage. ISME J. 2010, 4, 784–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanukollu, S.; Wemheuer, B.; Herber, J.; Billerbeck, S.; Lucas, J.; Daniel, R.; Simon, M.; Cypionka, H.; Engelen, B. Distinct compositions of free-living, particle-associated and benthic communities of the Roseobacter group in the North Sea. FEMS Microbiol. Ecol. 2016, 92, fiv145. [Google Scholar] [CrossRef] [PubMed]
- Thume, K.; Gebser, B.; Chen, L.; Meyer, N.; Kieber, D.J.; Pohnert, G. The metabolite dimethylsulfoxonium propionate extends the marine organosulfur cycle. Nature 2018, 563, 412–415. [Google Scholar] [CrossRef] [PubMed]
- Asher, E.; Dacey, J.W.; Ianson, D.; Peña, A.; Tortell, P.D. Concentrations and cycling of DMS, DMSP, and DMSO in coastal and offshore waters of the Subarctic Pacific during summer, 2010–2011. J. Geophys. Res. Oceans 2017, 122, 3269–3286. [Google Scholar] [CrossRef]
- Yang, M.; Archer, S.D.; Blomquist, B.W.; Ho, D.T.; Lance, V.P.; Torres, R.J. Lagrangian evolution of DMS during the Southern Ocean gas exchange experiment: The effects of vertical mixing and biological community shift. J. Geophys. Res. Oceans 2013, 118, 6774–6790. [Google Scholar] [CrossRef]
- Hatton, A.D.; Wilson, S.T. Particulate dimethylsulphoxide and dimethylsulphoniopropionate in phytoplankton cultures and Scottish coastal waters. Aquat. Sci. 2007, 69, 330–340. [Google Scholar] [CrossRef]
- Keller, M.D.; Bellows, W.K.; Guillard, R.R.L. Dimethylsulfide production in marine phytoplankton. ACS Symp. Ser. 1989, 393, 167–182. [Google Scholar]
- Hatton, A.D.; Shenoy, D.M.; Hart, M.C.; Mogg, A.; Green, D.H. Metabolism of DMSP, DMS and DMSO by the cultivable bacterial community associated with the DMSP-producing dinoflagellate Scrippsiella trochoidea. Biogeochemistry 2012, 110, 131–146. [Google Scholar] [CrossRef]
- Strom, S.; Wolfe, G.; Holmes, J.; Stecher, H.; Shimeneck, C.; Lambert, S. Chemical defense in the microplankton I: Feeding and growth rates of heterotrophic protists on the DMS-producing phytoplankter Emiliania huxleyi. Limnol. Oceanogr. 2003, 48, 217–229. [Google Scholar] [CrossRef]
- Evans, C.; Kadner, S.V.; Darroch, L.J.; Wilson, W.H.; Liss, P.S.; Malin, G. The relative significance of viral lysis and microzooplankton grazing as pathways of dimethylsulfoniopropionate (DMSP) cleavage: An Emiliania huxleyi culture study. Limnol. Oceanogr. 2007, 52, 1036–1045. [Google Scholar] [CrossRef] [Green Version]
- Lee, P.A.; de Mora, S.J.; Levasseur, M. A review of dimethylsulfoxide in aquatic environments. Atmos. Ocean 1999, 37, 439–456. [Google Scholar] [CrossRef]
- Kiene, R.P.; Gerard, G. Determination of trace levels of dimethylsulphoxide (DMSO) in seawater and rainwater. Mar. Chem. 1994, 47, 1–12. [Google Scholar] [CrossRef]
- Dixon, J.L.; Beale, R.; Nightingale, P.D. Microbial methanol uptake in northeast Atlantic waters. ISME J. 2011, 5, 704–716. [Google Scholar] [CrossRef] [Green Version]
- Dixon, J.L.; Beale, R.; Nightingale, P.D. Rapid biological oxidation of methanol in the tropical Atlantic: Significance as a microbial carbon source. Biogeosciences 2011, 8, 2707–2716. [Google Scholar] [CrossRef] [Green Version]
- Dixon, J.L.; Sargeant, S.; Nightingale, P.D.; Murrell, J.C. Gradients in microbial methanol uptake: Productive coastal upwelling waters to oligotrophic gyres in the Atlantic Ocean. ISME J. 2013, 7, 568–680. [Google Scholar] [CrossRef]
- Sargeant, S.L.; Murrell, J.C.; Nightingale, P.D.; Dixon, J.L. Seasonal variability in microbial methanol utilisation in coastal waters of the western English Channel. Mar. Ecol. Prog. Ser. 2016, 550, 53–64. [Google Scholar] [CrossRef]
- Sun, J.; Steindler, L.; Thrash, J.C.; Halsey, K.H.; Smith, D.P.; Carter, A.E.; Landry, Z.C.; Giovannoni, S.J. One carbon metabolism in SAR11 pelagic marine bacteria. PLoS ONE 2011, 6, e23973. [Google Scholar] [CrossRef] [Green Version]
- Halsey, K.H.; Giovannoni, S.J.; Graus, M.; Zhao, Y.; Landry, Z.; Thrash, J.C.; Vergin, K.L.; de Gouw, J. Biological cycling of volatile organic carbon by phytoplankton and bacterioplankton. Limnol. Oceanogr. 2017, 62, 2650–2661. [Google Scholar] [CrossRef]
- Kiene, R.P.; Linn, L.J.; Bruton, J.A. New and important roles for DMSP in marine microbial communities. J. Sea Res. 2000, 43, 209–224. [Google Scholar] [CrossRef]
- Vila-Costa, M.; Simó, R.; Harada, H.; Gasol, J.M.; Slezak, D.; Kiene, R.P. Dimethylsulfoniopropionate uptake by marine phytoplankton. Science 2006, 314, 652–654. [Google Scholar] [CrossRef]
- Simon, M.; Azam, F. Protein content and protein synthesis rates of planktonic marine bacteria. Mar. Ecol. Prog. Ser. 1989, 51, 201–213. [Google Scholar] [CrossRef]
- Robinson, C. Heterotrophic bacterial respiration. In Microbial Ecology of the Oceans; Kirchman, D.L., Ed.; Wiley: Hoboken, NJ, USA, 2008; pp. 299–334. [Google Scholar]
- Fagerbakke, K.M.; Heldal, M.; Norland, S. Content of carbon, nitrogen, oxygen, sulfur and phosphorus in native aquatic and cultured bacteria. Aquat. Microb. Ecol. 1996, 10, 15–27. [Google Scholar] [CrossRef]
- Kiene, R.P.; Linn, L.J.; González, J.; Moran, M.A.; Bruton, J.A. Dimethylsulfoniopropionate and methanethiol are important precursors of methionine and protein-sulfur in marine bacterioplankton. Appl. Environ. Microbiol. 1999, 65, 4549–4558. [Google Scholar] [CrossRef] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dixon, J.L.; Hopkins, F.E.; Stephens, J.A.; Schäfer, H. Seasonal Changes in Microbial Dissolved Organic Sulfur Transformations in Coastal Waters. Microorganisms 2020, 8, 337. https://doi.org/10.3390/microorganisms8030337
Dixon JL, Hopkins FE, Stephens JA, Schäfer H. Seasonal Changes in Microbial Dissolved Organic Sulfur Transformations in Coastal Waters. Microorganisms. 2020; 8(3):337. https://doi.org/10.3390/microorganisms8030337
Chicago/Turabian StyleDixon, Joanna L, Frances E Hopkins, John A Stephens, and Hendrik Schäfer. 2020. "Seasonal Changes in Microbial Dissolved Organic Sulfur Transformations in Coastal Waters" Microorganisms 8, no. 3: 337. https://doi.org/10.3390/microorganisms8030337
APA StyleDixon, J. L., Hopkins, F. E., Stephens, J. A., & Schäfer, H. (2020). Seasonal Changes in Microbial Dissolved Organic Sulfur Transformations in Coastal Waters. Microorganisms, 8(3), 337. https://doi.org/10.3390/microorganisms8030337