Application of Recombinase-Based In Vivo Expression Technology to Bifidobacterium longum subsp. longum for Identification of Genes Induced in the Gastrointestinal Tract of Mice

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Animal Experiments
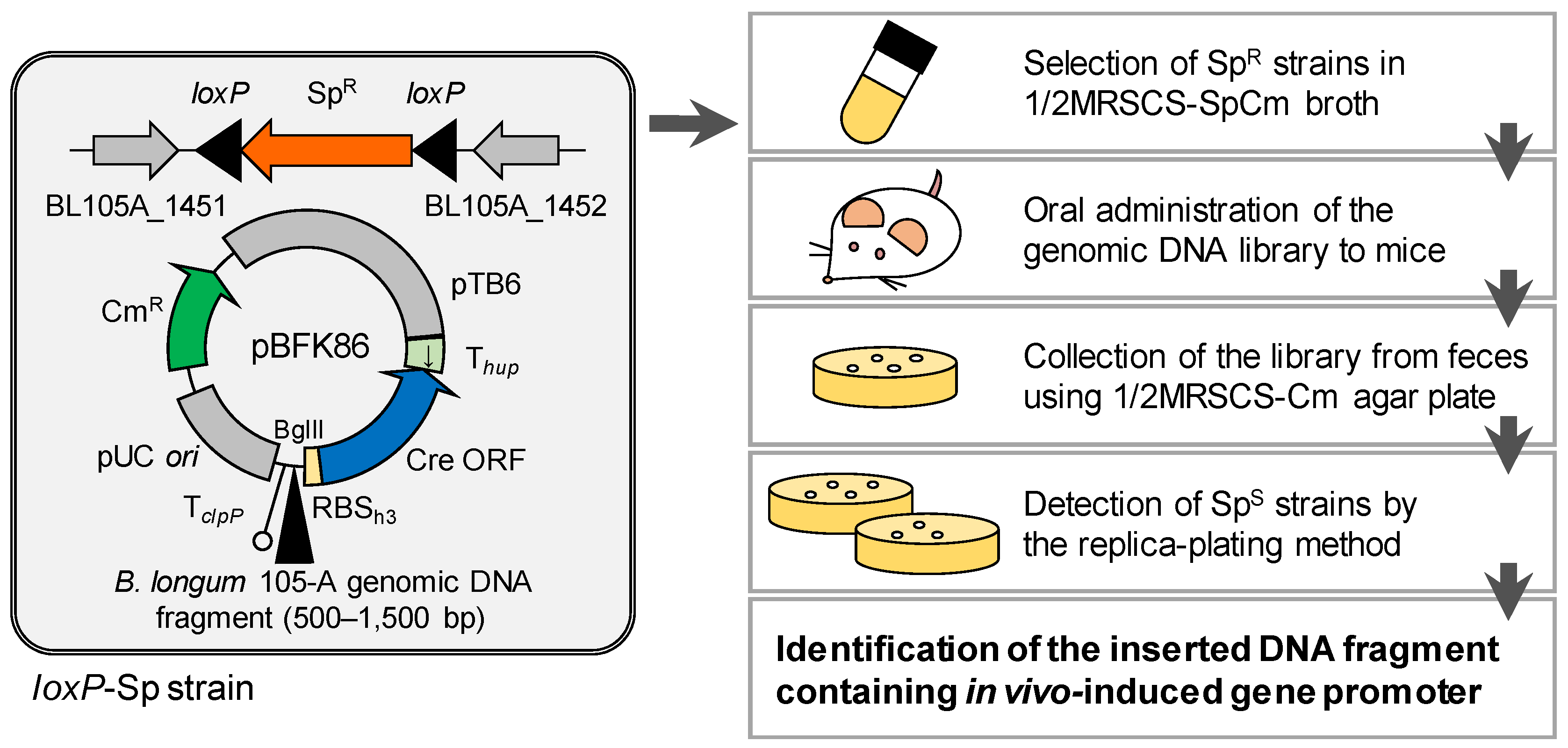
2.3. Generation of the B. longum Strain for the R-IVET System
2.4. Construction of a Plasmid Harboring the Cre Gene for the R-IVET System
2.4.1. Cloning of a Promoterless Cre Gene with an RBS
2.4.2. Insertion of a Transcriptional Terminator
2.4.3. Insertion of a Promoter
2.5. Evaluation of Basal Cre Expression Levels in Promoterless Cre Plasmids
2.6. Construction of the Genomic DNA Library
2.7. Screening for In Vivo-Induced Genes in B. longum
2.7.1. First and Second Trials
2.7.2. Third and Fourth Trials
2.8. RNA Extraction and qRT-PCR Analysis to Verify Specific In Vivo Gene Expression
2.8.1. Administration of B. longum 105-A Harboring pBFS63
2.8.2. RNA Extraction and qRT-PCR Analysis
3. Results
3.1. Development of the R-IVET System for B. longum 105-A
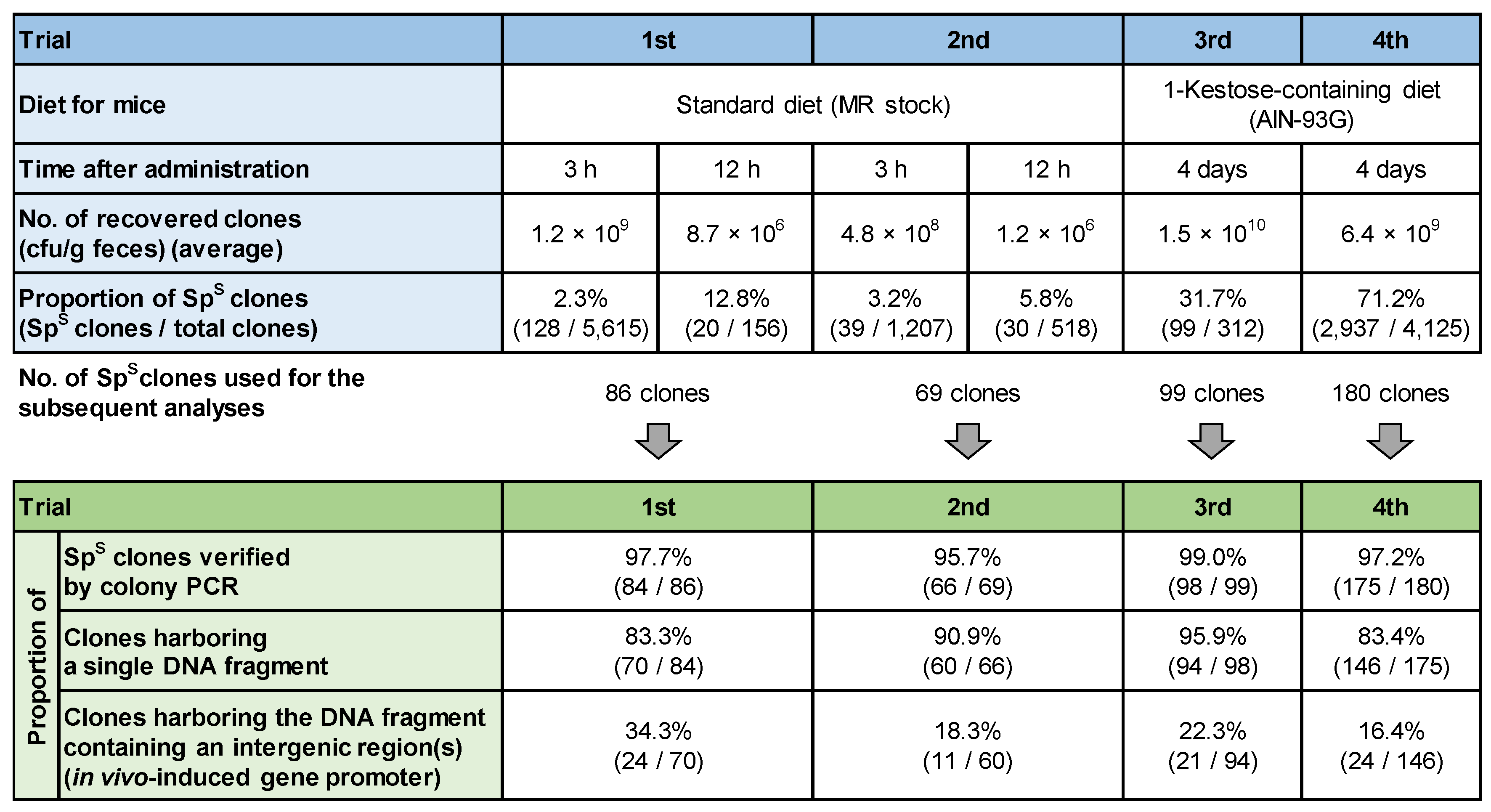
3.2. Construction of the Genomic DNA Library
3.3. Screening of In Vivo-Induced Genes
3.3.1. First and Second Trials
3.3.2. Third and Fourth Trials
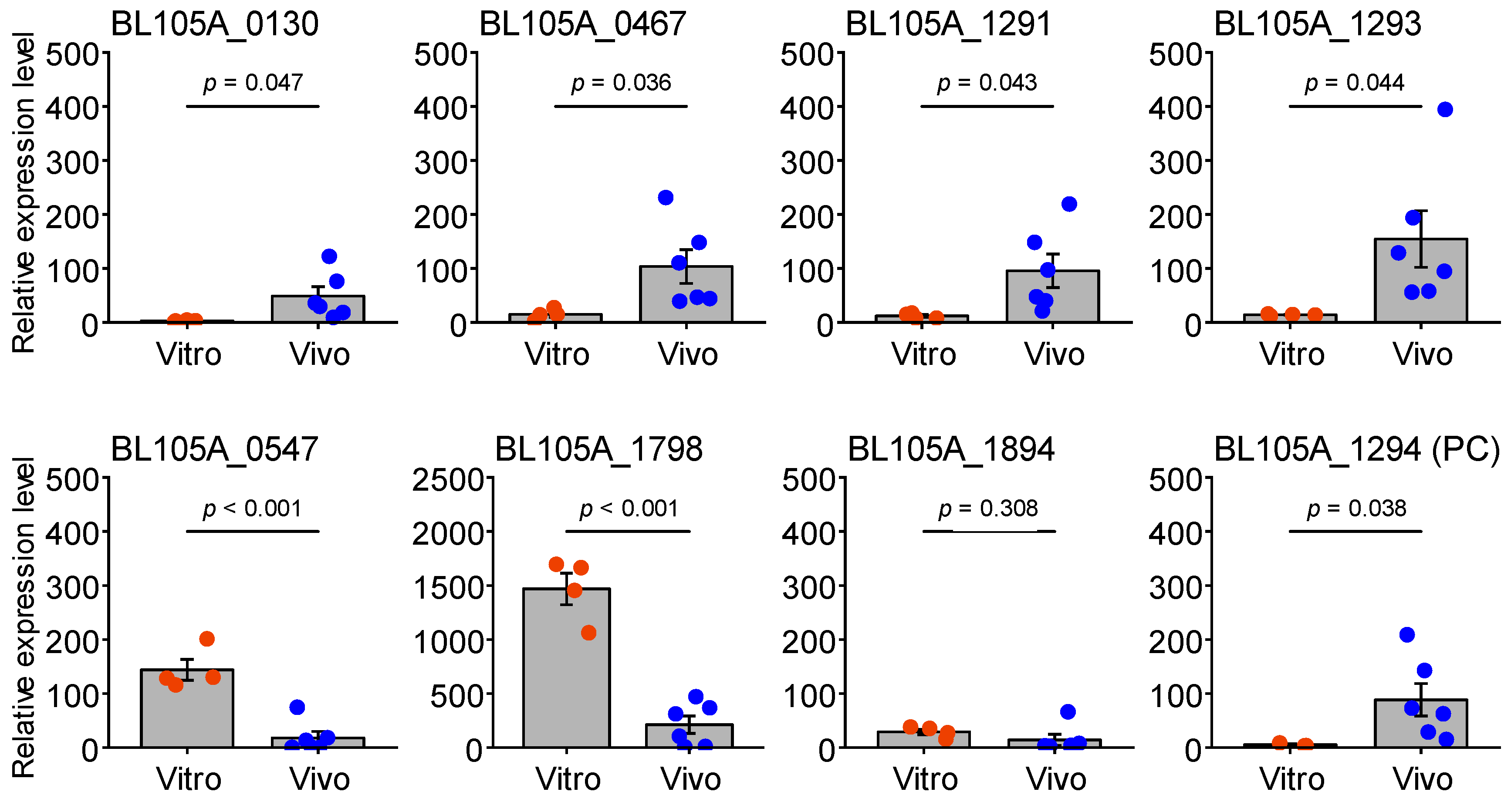
3.4. Verification of In Vivo-Induced Gene Expression in the Cecum
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nishijima, S.; Suda, W.; Oshima, K.; Kim, S.W.; Hirose, Y.; Morita, H.; Hattori, M. The gut microbiome of healthy Japanese and its microbial and functional uniqueness. DNA Res. 2016, 23, 125–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odamaki, T.; Kato, K.; Sugahara, H.; Hashikura, N.; Takahashi, S.; Xiao, J.Z.; Abe, F.; Osawa, R. Age-related changes in gut microbiota composition from newborn to centenarian: A cross-sectional study. BMC Microbiol. 2016, 16, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakanaka, M.; Hansen, M.E.; Gotoh, A.; Katoh, T.; Yoshida, K.; Odamaki, T.; Yachi, H.; Sugiyama, Y.; Kurihara, S.; Hirose, J.; et al. Evolutionary adaptation in fucosyllactose uptake systems supports bifidobacteria-infant symbiosis. Sci. Adv. 2019, 5, eaaw7696. [Google Scholar] [CrossRef] [Green Version]
- Mattarelli, P.; Biavati, B. Species in the genus Bifidobacterium. In The Bifidobacteria and Related Organisms: Biology, Taxonomy, Applications; Mattarelli, P., Biavati, B., Holzapfel, W.H., Wood, B.J.B., Eds.; Academic Press: London, UK, 2018; pp. 9–48. [Google Scholar] [CrossRef]
- Kato, K.; Odamaki, T.; Mitsuyama, E.; Sugahara, H.; Xiao, J.Z.; Osawa, R. Age-related changes in the composition of gut Bifidobacterium species. Curr. Microbiol. 2017, 74, 987–995. [Google Scholar] [CrossRef]
- Odamaki, T.; Bottacini, F.; Kato, K.; Mitsuyama, E.; Yoshida, K.; Horigome, A.; Xiao, J.Z.; van Sinderen, D. Genomic diversity and distribution of Bifidobacterium longum subsp. longum across the human lifespan. Sci. Rep. 2018, 8, 85. [Google Scholar] [CrossRef] [Green Version]
- Milani, C.; Mangifesta, M.; Mancabelli, L.; Lugli, G.A.; James, K.; Duranti, S.; Turroni, F.; Ferrario, C.; Ossiprandi, M.C.; van Sinderen, D.; et al. Unveiling bifidobacterial biogeography across the mammalian branch of the tree of life. ISME J. 2017, 11, 2834–2847. [Google Scholar] [CrossRef] [Green Version]
- Hidalgo-Cantabrana, C.; Delgado, S.; Ruiz, L.; Ruas-Madiedo, P.; Sánchez, B.; Margolles, A. Bifidobacteria and their health-promoting effects. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef]
- Leahy, S.C.; Higgins, D.G.; Fitzgerald, G.F.; van Sinderen, D. Getting better with bifidobacteria. J. Appl. Microbiol. 2005, 98, 1303–1315. [Google Scholar] [CrossRef]
- Fujita, K.; Sasaki, Y.; Kitahara, K. Degradation of plant arabinogalactan proteins by intestinal bacteria: Characteristics and functions of the enzymes involved. Appl. Microbiol. Biotechnol. 2019, 103, 7451–7457. [Google Scholar] [CrossRef]
- Katayama, T. Host-derived glycans serve as selected nutrients for the gut microbe: Human milk oligosaccharides and bifidobacteria. Biosci. Biotechnol. Biochem. 2016, 80, 621–632. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.; Nishiyama, K.; Miyajima, H.; Osawa, R.; Yamamoto, Y.; Mukai, T. Adhesion properties of a putative polymorphic fimbrial subunit protein from Bifidobacterium longum subsp. longum. Biosci. Microbiota Food Health 2016, 35, 19–27. [Google Scholar] [CrossRef]
- McCarville, J.L.; Dong, J.; Caminero, A.; Bermudez-Brito, M.; Jury, J.; Murray, J.A.; Duboux, S.; Steinmann, M.; Delley, M.; Tangyu, M.; et al. A commensal Bifidobacterium longum strain prevents gluten-related immunopathology in mice through expression of a serine protease inhibitor. Appl. Environ. Microbiol. 2017, 83, e01323–e01417. [Google Scholar] [CrossRef] [Green Version]
- Sonnenburg, J.L.; Chen, C.T.L.; Gordon, J.I. Genomic and metabolic studies of the impact of probiotics on a model gut symbiont and host. PLoS Biol. 2006, 4, e413. [Google Scholar] [CrossRef] [Green Version]
- Sugahara, H.; Odamaki, T.; Fukuda, S.; Kato, T.; Xiao, J.Z.; Abe, F.; Kikuchi, J.; Ohno, H. Probiotic Bifidobacterium longum alters gut luminal metabolism through modification of the gut microbial community. Sci. Rep. 2015, 5, 13548. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, E.; Shima, T.; Suda, K.; Shirasawa, Y.; Sato, T.; Umesaki, Y. Comparison of Bifidobacterium breve strain Yakult transcriptomes in germ-free mice with those in fecal cultures. J. Biosci. Bioeng. 2011, 112, 451–457. [Google Scholar] [CrossRef]
- O’Connell-Motherway, M.; Zomer, A.; Leahy, S.C.; Reunanen, J.; Bottacini, F.; Claesson, M.J.; O’Brien, F.; Flynn, K.; Casey, P.G.; Munoz, J.A.M.; et al. Functional genome analysis of Bifidobacterium breve UCC2003 reveals type IVb tight adherence (Tad) pili as an essential and conserved host-colonization factor. Proc. Natl. Acad. Sci. USA 2011, 108, 11217–11222. [Google Scholar] [CrossRef] [Green Version]
- O’Connell-Motherway, M.; O’Brien, F.; O’Driscoll, T.; Casey, P.G.; Shanahan, F.; van Sinderen, D. Carbohydrate syntrophy enhances the establishment of Bifidobacterium breve UCC2003 in the neonatal gut. Sci. Rep. 2018, 8, 10627. [Google Scholar] [CrossRef]
- Bron, P.A.; Grangette, C.; Mercenier, A.; de Vos, W.M.; Kleerebezem, M. Identification of Lactobacillus plantarum genes that are induced in the gastrointestinal tract of mice. J. Bacteriol. 2004, 186, 5721–5729. [Google Scholar] [CrossRef] [Green Version]
- Bachmann, H.; Kleerebezem, M.; van Hylckama Vlieg, J.E.T. High-throughput identification and validation of in situ-expressed genes of Lactococcus lactis. Appl. Environ. Microbiol. 2008, 74, 4727–4736. [Google Scholar] [CrossRef] [Green Version]
- Bachmann, H.; de Wilt, L.; Kleerebezem, M.; van Hylckama Vlieg, J.E.T. Time-resolved genetic responses of Lactococcus lactis to a dairy environment. Environ. Microbiol. 2010, 12, 1260–1270. [Google Scholar] [CrossRef]
- Hanin, A.; Sava, I.; Bao, Y.Y.; Huebner, J.; Hartke, A.; Auffray, Y.; Sauvageot, N. Screening of in vivo activated genes in Enterococcus faecalis during insect and mouse infections and growth in urine. PLoS ONE 2010, 5, e11879. [Google Scholar] [CrossRef] [Green Version]
- Junjua, M.; Galia, W.; Gaci, N.; Uriot, O.; Genay, M.; Bachmann, H.; Kleerebezem, M.; Dary, A.; Roussel, Y. Development of the recombinase-based in vivo expression technology in Streptococcus thermophilus and validation using the lactose operon promoter. J. Appl. Microbiol. 2014, 116, 620–631. [Google Scholar] [CrossRef] [PubMed]
- Uriot, O.; Galia, W.; Awussi, A.A.; Perrin, C.; Denis, S.; Chalancon, S.; Lorson, E.; Poirson, C.; Junjua, M.; Le Roux, Y.; et al. Use of the dynamic gastro-intestinal model TIM to explore the survival of the yogurt bacterium Streptococcus thermophilus and the metabolic activities induced in the simulated human gut. Food Microbiol. 2016, 53, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, H.; Takeuchi, A.; Kano, Y. Construction of Escherichia coli–Bifidobacterium longum shuttle vector transforming B. longum 105-A and 108-A. Biosci. Biotechnol. Biochem. 1997, 61, 1211–1212. [Google Scholar] [CrossRef] [PubMed]
- De Man, J.C.; Rogosa, M.; Sharpe, M.E. A medium for the cultivation of lactobacilli. J. Appl. Bacteriol. 1960, 23, 130–135. [Google Scholar] [CrossRef]
- Hirayama, Y.; Sakanaka, M.; Fukuma, H.; Murayama, H.; Kano, Y.; Fukiya, S.; Yokota, A. Development of a double-crossover markerless gene deletion system in Bifidobacterium longum: Functional analysis of the α-galactosidase gene for raffinose assimilation. Appl. Environ. Microbiol. 2012, 78, 4984–4994. [Google Scholar] [CrossRef] [Green Version]
- Kanegae, Y.; Lee, G.; Sato, Y.; Tanaka, M.; Nakal, M.; Sakaki, T.; Sugano, S.; Saito, I. Efficient gene activation in mammalian cells by using recombinant adenovirus expressing site-specific Cre recombinase. Nucleic Acids Res. 1995, 23, 3816–3821. [Google Scholar] [CrossRef] [Green Version]
- Sakanaka, M.; Tamai, S.; Hirayama, Y.; Onodera, A.; Koguchi, H.; Kano, Y.; Yokota, A.; Fukiya, S. Functional analysis of bifidobacterial promoters in Bifidobacterium longum and Escherichia coli using the α-galactosidase gene as a reporter. J. Biosci. Bioeng. 2014, 118, 489–495. [Google Scholar] [CrossRef]
- Yasui, K.; Kano, Y.; Tanaka, K.; Watanabe, K.; Shimizu-Kadota, M.; Yoshikawa, H.; Suzuki, T. Improvement of bacterial transformation efficiency using plasmid artificial modification. Nucleic Acids Res. 2009, 37, e3. [Google Scholar] [CrossRef]
- He, J.; Sakaguchi, K.; Suzuki, T. Determination of the ribosome-binding sequence and spacer length between binding site and initiation codon for efficient protein expression in Bifidobacterium longum 105-A. J. Biosci. Bioeng. 2012, 113, 442–444. [Google Scholar] [CrossRef]
- Sternberg, N.; Sauer, B.; Hoess, R.; Abremski, K. Bacteriophage P1 cre gene and its regulatory region. Evidence for multiple promoters and for regulation by DNA methylation. J. Mol. Biol. 1986, 187, 197–212. [Google Scholar] [CrossRef]
- Llanos, R.M.; Harris, C.J.; Hillier, A.J.; Davidson, B.E. Identification of a novel operon in Lactococcus lactis encoding three enzymes for lactic acid synthesis: Phosphofructokinase, pyruvate kinase, and lactate dehydrogenase. J. Bacteriol. 1993, 175, 2541–2551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pátek, M.; Hochmannová, J.; Jelínková, M.; Nešvera, J.; Eggeling, L. Analysis of the leuB gene from Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 1998, 50, 42–47. [Google Scholar] [CrossRef]
- Ruiz, L.; O’Connell-Motherway, M.; Lanigan, N.; van Sinderen, D. Transposon mutagenesis in Bifidobacterium breve: Construction and characterization of a Tn5 transposon mutant library for Bifidobacterium breve UCC2003. PLoS ONE 2013, 8, e64699. [Google Scholar] [CrossRef] [Green Version]
- Kanesaki, Y.; Masutani, H.; Sakanaka, M.; Shiwa, Y.; Fujisawa, T.; Nakamura, Y.; Yokota, A.; Fukiya, S.; Suzuki, T.; Yoshikawa, H. Complete genome sequence of Bifidobacterium longum 105-A, a strain with high transformation efficiency. Genome Announc. 2014, 2, e01311–e01314. [Google Scholar] [CrossRef] [Green Version]
- Galperin, M.Y.; Makarova, K.S.; Wolf, Y.I.; Koonin, E.V. Expanded microbial genome coverage and improved protein family annotation in the COG database. Nucleic Acids Res. 2015, 43, D261–D269. [Google Scholar] [CrossRef]
- Sakanaka, M.; Nakakawaji, S.; Nakajima, S.; Fukiya, S.; Abe, A.; Saburi, W.; Mori, H.; Yokota, A. A transposon mutagenesis system for Bifidobacterium longum subsp. longum based on an IS3 family insertion sequence, ISBlo11. Appl. Environ. Microbiol. 2018, 84, e00824–e00918. [Google Scholar] [CrossRef] [Green Version]
- Tanno, H.; Fujii, T.; Ose, R.; Hirano, K.; Tochio, T.; Endo, A. Characterization of fructooligosaccharide-degrading enzymes in human commensal Bifidobacterium longum and Anaerostipes caccae. Biochem. Biophys. Res. Commun. 2019, 518, 294–298. [Google Scholar] [CrossRef]
- Clarke, L.; Carbon, J. A colony bank containing synthetic Col El hybrid plasmids representative of the entire E. coli genome. Cell 1976, 9, 91–99. [Google Scholar] [CrossRef]
- Marco, M.L.; Bongers, R.S.; de Vos, W.M.; Kleerebezem, M. Spatial and temporal expression of Lactobacillus plantarum genes in the gastrointestinal tracts of mice. Appl. Environ. Microbiol. 2007, 73, 124–132. [Google Scholar] [CrossRef] [Green Version]
- Bottacini, F.; Zomer, A.; Milani, C.; Ferrario, C.; Lugli, G.A.; Egan, M.; Ventura, M.; van Sinderen, D. Global transcriptional landscape and promoter mapping of the gut commensal Bifidobacterium breve UCC2003. BMC Genom. 2017, 18, 991. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Description 1 | Source or Reference |
|---|---|---|
| Escherichia coli | ||
| E. coli DH5α | F−, Φ80d lacZΔM15, Δ(lacZYA-argF) U169, deoR, recA1, endA1, hsdR17(rK− mK+), phoA, supE44, λ−, thi-1, gyrA96, relA1 | National BioResource Project (NIG, Mishima, Japan) |
| Bifidobacterium | ||
| B. longum subsp. longum 105-A (JCM 31944) | Human fecal isolate | [25] |
| loxP-Sp strain | B. longum 105-A derivative strain harboring loxP-SpR-loxP cassette on the chromosome, SpR | This study |
| No. | PCR Product 1 | DNA Template | Cloning Strategy 2 | Primer | Nucleotide Sequence (5′-3′) 3 |
|---|---|---|---|---|---|
| Integration of loxP-SpR-loxP into a chromosome of B. longum 105-A (construction of loxP-Sp strain) | |||||
| 1 | SpR gene | pBS423 [27] | Blunt-end ligation | Pr-Blo0041 | GCATGCCTGCAGGTCGATTTTC |
| Pr-Blo0042 | CAAAAAAATTGAAAAAAGTGTTTCCAC | ||||
| 2 | Homologous region to BL105A_1451 locus (HR1) | B. longum 105-A genomic DNA | Restriction-ligation | Pr-Blo0100 | GCGAATTCATGACGTACGATTTCACGTCG |
| Pr-Blo0101 | TAGAATTCCGCAATCGCGGATGCATGCCGT | ||||
| 3 | Homologous region to BL105A_1452 locus (HR2) | B. longum 105-A genomic DNA | Restriction-ligation | Pr-Blo0098 | AGGGATCCGTGTCCTGGAAAGACGAATGCC |
| Pr-Blo0099 | TGGGATCCATGTCCGTTTCGCAGTCACCGG | ||||
| 4 | HR1-loxP-SpR-loxP-HR2 | pBFH35 (this study) | In-Fusion cloning | Pr-Blo0119 | TATATATGAGTACTGAGGTCGACTCTAGAGGATCC |
| Pr-Blo0120 | AAACGACGGCCAGTTAAACGACGGCCAGTGAATTG | ||||
| 5 | pBS423ΔrepA lacking SpR gene | pBFS423ΔrepA [27] | In-Fusion cloning | Pr-Blo0116 | CAGTACTCATATATACTTTAGATTGATTTA |
| Pr-Blo0117 | AAGCTTGCATGCCTGCAGATAGGCAG | ||||
| Construction of Cre expression plasmid | |||||
| 6 | CmR gene | pBFS38 [29] | In-Fusion cloning | Pr-Blo0239 | AAAGTATATATGAGTACTTGGGCGCGGCGGCCATGAAG |
| Pr-Blo0240 | GCGGCCGCGCCGGCATGCATTATGGAAGCGCTGAACTAGTC | ||||
| 7 | BglII-RBSh4-Cre ORF | Bacteriophage P1 genomic DNA | In-Fusion cloning | Pr-Blo0247 | CTTCCCGGCGAGATCCTAATCGCCATCTTCCAGC |
| Pr-Blo0249 | GATTACTTCGGCGCGAGATCTCCCAAGAAGGATGCTATGTCCAATTTACTGACCGTACAC | ||||
| 8 | BglII-RBSh3-Cre ORF | Bacteriophage P1 genomic DNA | In-Fusion cloning | Pr-Blo0247 | CTTCCCGGCGAGATCCTAATCGCCATCTTCCAGC |
| Pr-Blo0257 | GATTACTTCGGCGCGAGATCTCCCAAGAAGGATGCATGTCCAATTTACTGACCGTACAC | ||||
| 9 | Tlas | L. lactis subsp. cremoris MG1363 genomic DNA | Restriction-ligation | Pr-Blo0258 | ACGTGGATCCGGACAATATGGGGTAAGCG |
| Pr-Blo0259 | AAGAAGATCTCTAAAGCTGACGGGGTAAAC | ||||
| 10 | Trps9 | B. longum 105-A genomic DNA | Restriction-ligation | Pr-Blo0264 | TGACCGAGATCTTGTGGATGATACACCGGACACTC |
| Pr-Blo0265 | TAGGGATCCTCGTGGAGCGCAAGAAGGCTGGTCTG | ||||
| 11 | TleuB | C. glutamicum ATCC 13032 genomic DNA | Restriction-ligation | Pr-Blo0260 | GTATGCAGATCTCCAGCAAGTATTTACACCAAC |
| Pr-Blo0261 | AGTGGATCCTGCGATGCTGCTGCGTCACTTAG | ||||
| 12 | TclpP stem loop-BglII-RBSh3-Cre ORF | pBFK71 (this study) | In-Fusion cloning | Pr-Blo0277 | ATGGCTTCCCGGCGACTAATCGCCATCTTCCAGC |
| Pr-Blo0280 | ATTACTTCGGCGCGAaaaaccctcggtcggtctgaccgggggttttAGATCTCCCAAGAAGGATGCATG | ||||
| 13 | PcscBA | B. longum 105-A genomic DNA | Restriction-ligation | Pr-Blo0292 | ATTAGATCTTTGGTTGGTTATTGGTTATGTAAC |
| Pr-Blo0293 | ATTAGATCTCCGAGTCCCACACGATTTCTC | ||||
| Genotypic analysis of the loxP-Sp strain | |||||
| 14 | SpR gene | Genomic DNA of loxP-Sp strain | NA | Pr-Blo0099 | TGGGATCCATGTCCGTTTCGCAGTCACCGG |
| Pr-Blo0100 | GCGAATTCATGACGTACGATTTCACGTCG | ||||
| Determination of Inserted DNA Fragments in the R-IVET Library | |||||
| 15 | Inserted DNA fragment | pBFK86 derivative carrying a random DNA fragment (this study) | NA | Pr-Blo0277 | ATGGCTTCCCGGCGACTAATCGCCATCTTCCAGC |
| Pr-Blo0318 | GTAAGCGGCAGGGTCGGAACAGGAGAGCG | ||||
| qRT-PCR analysis | |||||
| 16 | BL105A_0130 | B. longum 105-A genomic DNA | NA | Pr-Blo0414 | AGGCGAAAGAACGGCTATGC |
| Pr-Blo0415 | GACTTCAGGATGGCGACCAG | ||||
| 17 | BL105A_0467 | B. longum 105-A genomic DNA | NA | Pr-Blo0416 | CCTTGTTGCCCAGACCCAAC |
| Pr-Blo0417 | CATAAGAGCGACGCAGCGAG | ||||
| 18 | BL105A_0547 | B. longum 105-A genomic DNA | NA | Pr-Blo0432 | TCGGCAACCATGTTGAGCAC |
| Pr-Blo0433 | GCCTACCCCGATCAGCTCTC | ||||
| 19 | BL105A_1291 | B. longum 105-A genomic DNA | NA | Pr-Blo0434 | ATGTTCAAGCCGAAGGCCAC |
| Pr-Blo0435 | GCCATCCACATCGAAGCAGG | ||||
| 20 | BL105A_1293 | B. longum 105-A genomic DNA | NA | Pr-Blo0436 | AAATCGGCAACGCCACCTAC |
| Pr-Blo0437 | CGCAGGAACATCACGGTAGC | ||||
| 21 | BL105A_1294 | B. longum 105-A genomic DNA | NA | Pr-Blo0408 | AAGGTCGACCACCACTACCG |
| Pr-Blo0409 | CTCGTATTCCCAGCGGACCA | ||||
| 22 | BL105A_1798 | B. longum 105-A genomic DNA | NA | Pr-Blo0428 | GCATCGCGGGAAGAACAGAC |
| Pr-Blo0429 | ATACGCAAACGGCTTCACCG | ||||
| 23 | BL105A_1894 | B. longum 105-A genomic DNA | NA | Pr-Blo0430 | CCACCGACGACCCACTTTTG |
| Pr-Blo0431 | AGTCGAACCAGACCATCCCG | ||||
| 24 | BL105A_1946 | B. longum 105-A genomic DNA | NA | Pr-Blo0372 | GCCTTCGCGATCTGCTGATCTAG |
| Pr-Blo0373 | ACCCGTAATACGGTGAAGCGTAG | ||||
| No. | In Vivo Induced Genes 1 | Annotation 1 | Identified Round | COG Category 2, 3 |
|---|---|---|---|---|
| 1 | BL105A_0064 | Hypothetical protein | 2nd | – |
| 2 | BL105A_0075 | Hypothetical protein | 3rd | S |
| 3 | BL105A_0117 | GrpE protein | 1st | O |
| 4 | BL105A_0130 | Presumable pilin subunit for the Tad-pili | 4th | – |
| 5 | BL105A_0136 | Recombination protein RecR | 1st | L |
| 6 | BL105A_0138 | Hypothetical protein | 4th | – |
| 7 | BL105A_0202 | ABC transporter permease component | 4th | G |
| 8 | BL105A_0204 | Glycoside hydrolase family 127 β-l-arabinofuranosidase | 4th | S |
| 9 | BL105A_0248 | Hypothetical protein | 3rd | – |
| 10 | BL105A_0262 | Hypothetical protein | 4th | – |
| 11 | BL105A_0267 | Hypothetical protein | 1st, 2nd, 4th | – |
| 12 | BL105A_0338 | Ribonuclease VapC | 4th | R |
| 13 | BL105A_0374 | Magnesium-translocating P-type ATPase | 4th | – |
| 14 | BL105A_0377 | Hypothetical protein | 1st | – |
| 15 | BL105A_0414 | Oligosaccharide repeat unit polymerase Wzy | 2nd | M |
| 16 | BL105A_0415 | Hypothetical protein | 4th | M |
| 17 | BL105A_0422 | Transposase | 4th | X |
| 18 | BL105A_0423 | Integrase catalytic region | 1st | X |
| 19 | BL105A_0467 | Putative adhesin | 3rd | X, R |
| 20 | BL105A_0490 | Putative ABC transporter ATP-binding component | 3rd | E |
| 21 | BL105A_0507 | Peptides ABC transporter ATP-binding component | 1st | P, E |
| 22 | BL105A_0534 | Hypothetical protein | 3rd | V, M |
| 23 | BL105A_0540 | Hypothetical protein | 3rd | V |
| 24 | BL105A_0547 | ATPase of the ABC transporter | 3rd, 4th | E |
| 25 | BL105A_0662 | Transcriptional regulator | 2nd | M |
| 26 | BL105A_0669 | Putative phosphoribosylpyrophosphate amidotransferase | 3rd | R |
| 27 | BL105A_0776 | Hypothetical protein | 3rd, 4th | – |
| 28 | BL105A_0812 | Shikimate kinase/3-dehydroquinate synthase | 4th | E |
| 29 | BL105A_0835 | NAD(P) transhydrogenase α-2 subunit | 2nd | C |
| 30 | BL105A_0854 | Hypothetical protein | 2nd | V |
| 31 | BL105A_0900 | Hypothetical protein | 3rd | – |
| 32 | BL105A_0929 | Hypothetical protein | 1st | – |
| 33 | BL105A_0934 | Phosphoribosyl-ATP pyrophosphatase | 2nd | E |
| 34 | BL105A_1028 | Hypothetical protein | 3rd | – |
| 35 | BL105A_1049 | Hypothetical protein | 1st | – |
| 36 | BL105A_1053 | Hypothetical protein | 4th | – |
| 37 | BL105A_1079 | tRNA N6-adenosine threonylcarbamoyltransferase | 1st | J |
| 38 | BL105A_1118 | Hypothetical protein | 1st | – |
| 39 | BL105A_1123 | RecX-like protein | 3rd | O |
| 40 | BL105A_1233 | Cell division protein FtsW | 3rd | D |
| 41 | BL105A_1250 | 16S RNA methylase | 1st | J |
| 42 | BL105A_1253 | Transporter | 2nd | G |
| 43 | BL105A_1291 | Serine protease inhibitor | 1st | O |
| 44 | BL105A_1293 | Galactoside transport protein | 1st | P |
| 45 | BL105A_1371 | ABC-type fructose transport system ATPase subunit FruK | 4th | G |
| 46 | BL105A_1419 | Hypothetical protein | 3rd | I |
| 47 | BL105A_1426 | Hypothetical protein | 4th | – |
| 48 | BL105A_1456 | Sugar kinase in PfkB family | 4th | G, F |
| 49 | BL105A_1489 | Endonuclease | 4th | L |
| 50 | BL105A_1517 | Peptide chain release factor 1 | 4th | J |
| 51 | BL105A_1556 | Hypothetical protein | 4th | N |
| 52 | BL105A_1562 | tRNA-Phe | 3rd | – |
| 53 | BL105A_1583 | Hypothetical protein | 3rd | – |
| 54 | BL105A_1603 | Sugar ABC transporter permease component | 2nd | G |
| 55 | BL105A_1605 | Hypothetical protein | 1st | – |
| 56 | BL105A_1637 | DNA-directed RNA polymerase α subunit | 1st | K |
| 57 | BL105A_1680 | Amino acid transporter | 1st | E |
| 58 | BL105A_1696 | Hypothetical protein | 4th | L |
| 59 | BL105A_1707 | Possible extracellular exo-xylanase | 4th | G |
| 60 | BL105A_1708 | endo-1,4-β-Xylanase | 2nd | G |
| 61 | BL105A_1718 | Hypothetical protein | 1st | G |
| 62 | BL105A_1733 | 16S ribosomal RNA | 1st | – |
| 63 | BL105A_1798 | Putative glycosyltransferase | 1st, 3rd | M |
| 64 | BL105A_1810 | Probable potassium uptake protein Kup | 3rd | P |
| 65 | BL105A_1828 | Hypothetical protein | 1st | – |
| 66 | BL105A_1834 | Hypothetical protein | 1st, 1st | L |
| 67 | BL105A_1857 | Hypothetical protein | 4th | R, G |
| 68 | BL105A_1883 | α-Glucosidase | 3rd | G |
| 69 | BL105A_1885 | Glycosidase | 1st | G |
| 70 | BL105A_1886 | Permease protein of ABC transporter system for sugars | 4th | G |
| 71 | BL105A_1894 | Raffinose transport system permease protein | 2nd, 3rd | G |
| 72 | BL105A_1910 | Lipopolysaccharide kinase | 3rd | T |
| 73 | BL105A_1945 | Preprotein translocase subunit YidC | 1st | M |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koguchi, H.; Ishigami, N.; Sakanaka, M.; Yoshida, K.; Hiratou, S.; Shimada, M.; Fukiya, S.; Sonoyama, K.; Yokota, A. Application of Recombinase-Based In Vivo Expression Technology to Bifidobacterium longum subsp. longum for Identification of Genes Induced in the Gastrointestinal Tract of Mice. Microorganisms 2020, 8, 410. https://doi.org/10.3390/microorganisms8030410
Koguchi H, Ishigami N, Sakanaka M, Yoshida K, Hiratou S, Shimada M, Fukiya S, Sonoyama K, Yokota A. Application of Recombinase-Based In Vivo Expression Technology to Bifidobacterium longum subsp. longum for Identification of Genes Induced in the Gastrointestinal Tract of Mice. Microorganisms. 2020; 8(3):410. https://doi.org/10.3390/microorganisms8030410
Chicago/Turabian StyleKoguchi, Hiroka, Natsumi Ishigami, Mikiyasu Sakanaka, Kako Yoshida, Sayaka Hiratou, Mina Shimada, Satoru Fukiya, Kei Sonoyama, and Atsushi Yokota. 2020. "Application of Recombinase-Based In Vivo Expression Technology to Bifidobacterium longum subsp. longum for Identification of Genes Induced in the Gastrointestinal Tract of Mice" Microorganisms 8, no. 3: 410. https://doi.org/10.3390/microorganisms8030410
APA StyleKoguchi, H., Ishigami, N., Sakanaka, M., Yoshida, K., Hiratou, S., Shimada, M., Fukiya, S., Sonoyama, K., & Yokota, A. (2020). Application of Recombinase-Based In Vivo Expression Technology to Bifidobacterium longum subsp. longum for Identification of Genes Induced in the Gastrointestinal Tract of Mice. Microorganisms, 8(3), 410. https://doi.org/10.3390/microorganisms8030410