Investigation of Heterologously Expressed Glucose-6-Phosphate Dehydrogenase Genes in a Yeast zwf1 Deletion


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Media, and Culture Conditions
2.2. Serial Drop Dilution Assays
2.3. Construction of Deletion Mutants, Cloning, and Tagging of Genes
2.4. Enzyme Assays and Western Blot Analysis
2.5. Determination of Chronological Life Span
3. Results
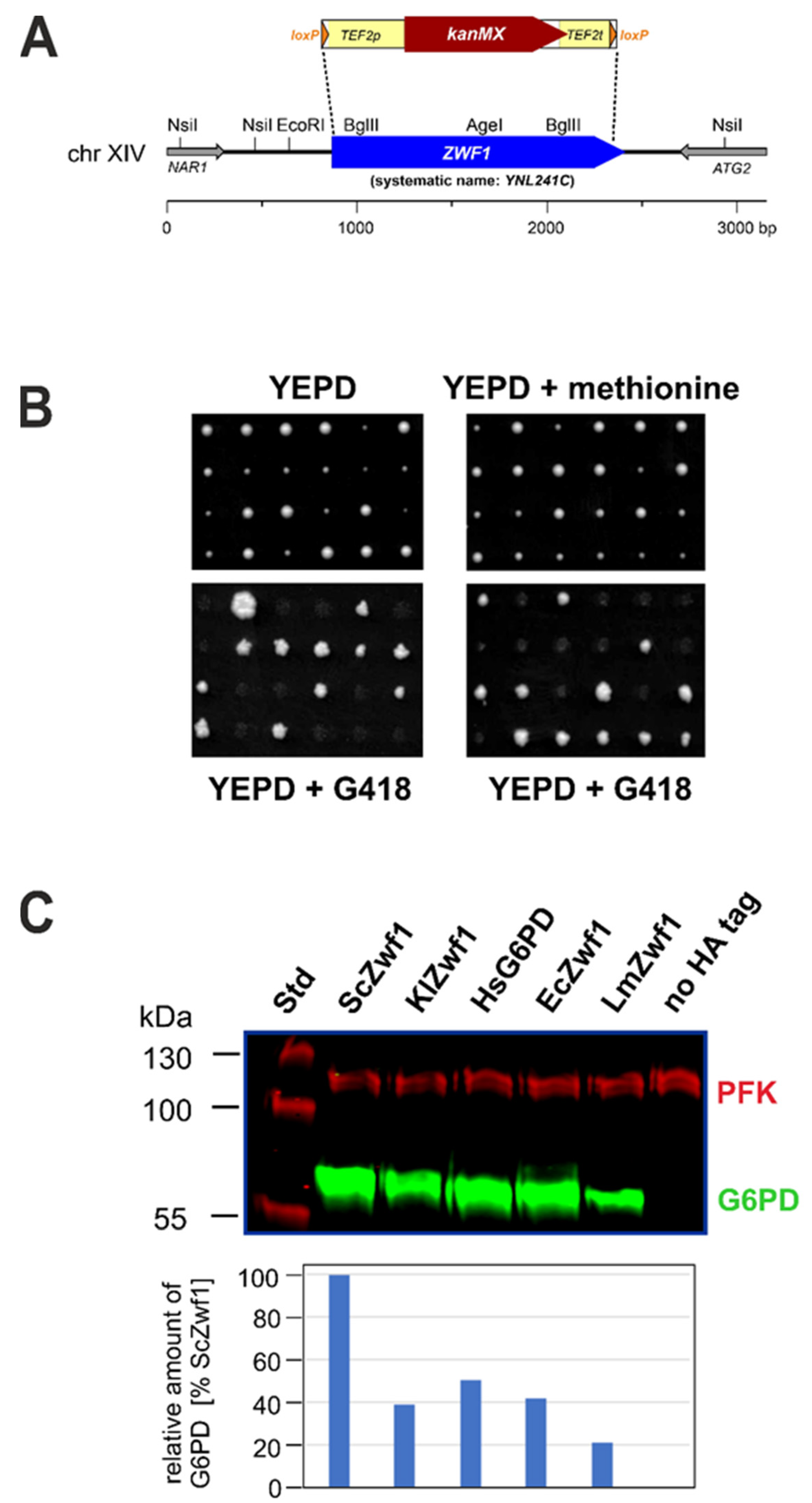
3.1. A Yeast System for Heterologous Expression of G6PD Genes
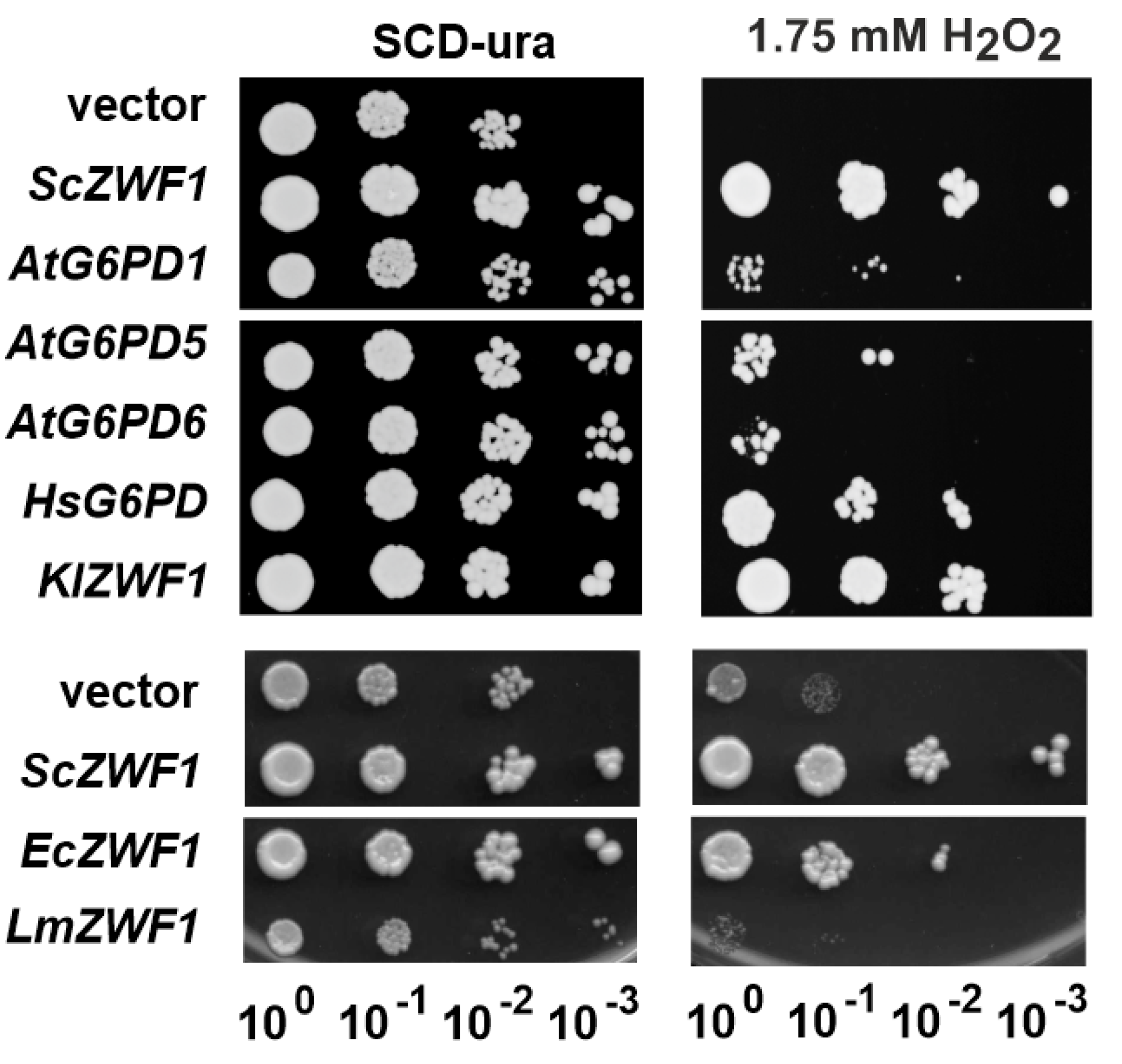
3.2. Heterologous G6PD Genes from Different Biological Kingdoms Complement a Yeast zwf1 Deletion
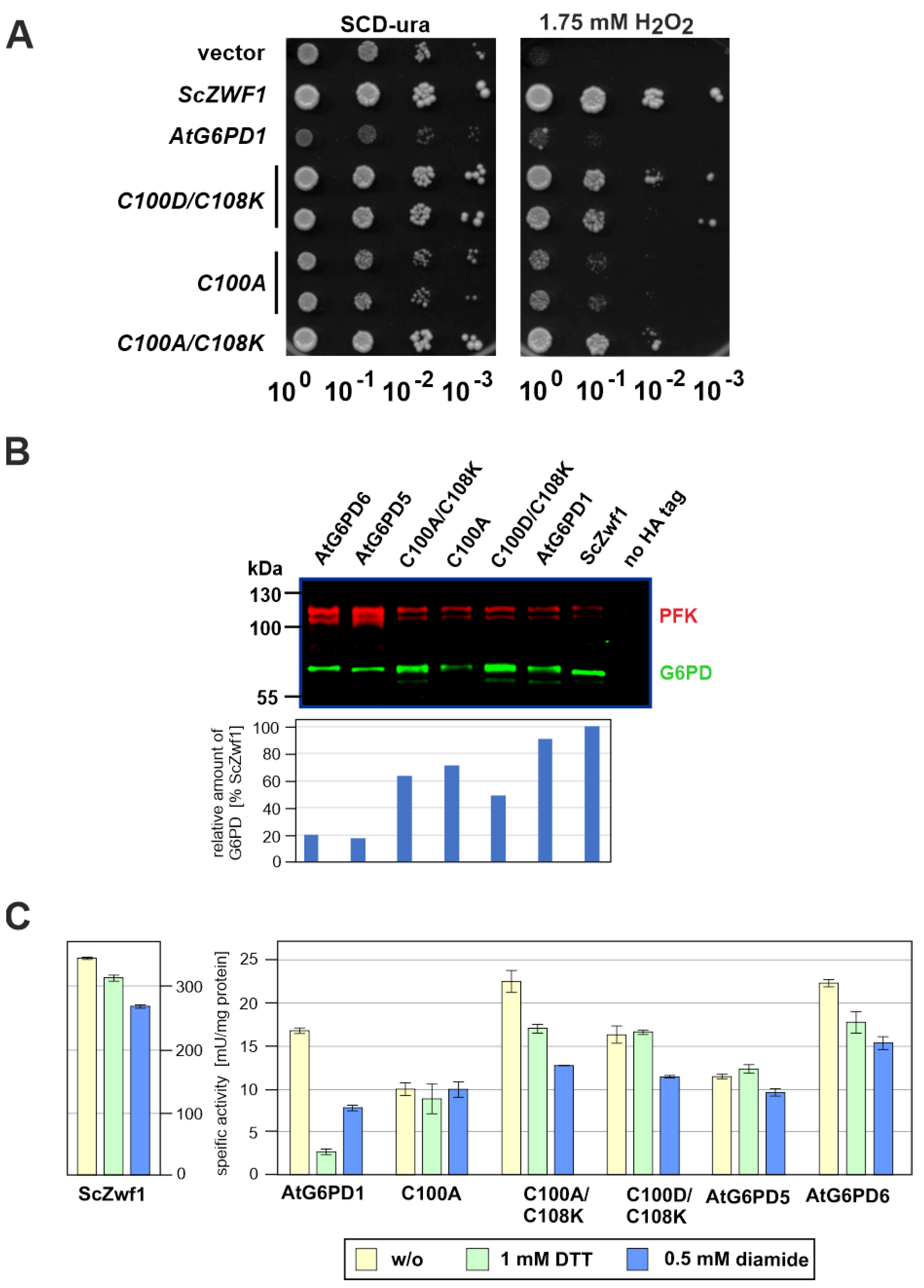
3.3. Mutational Analysis of Plant G6PD1
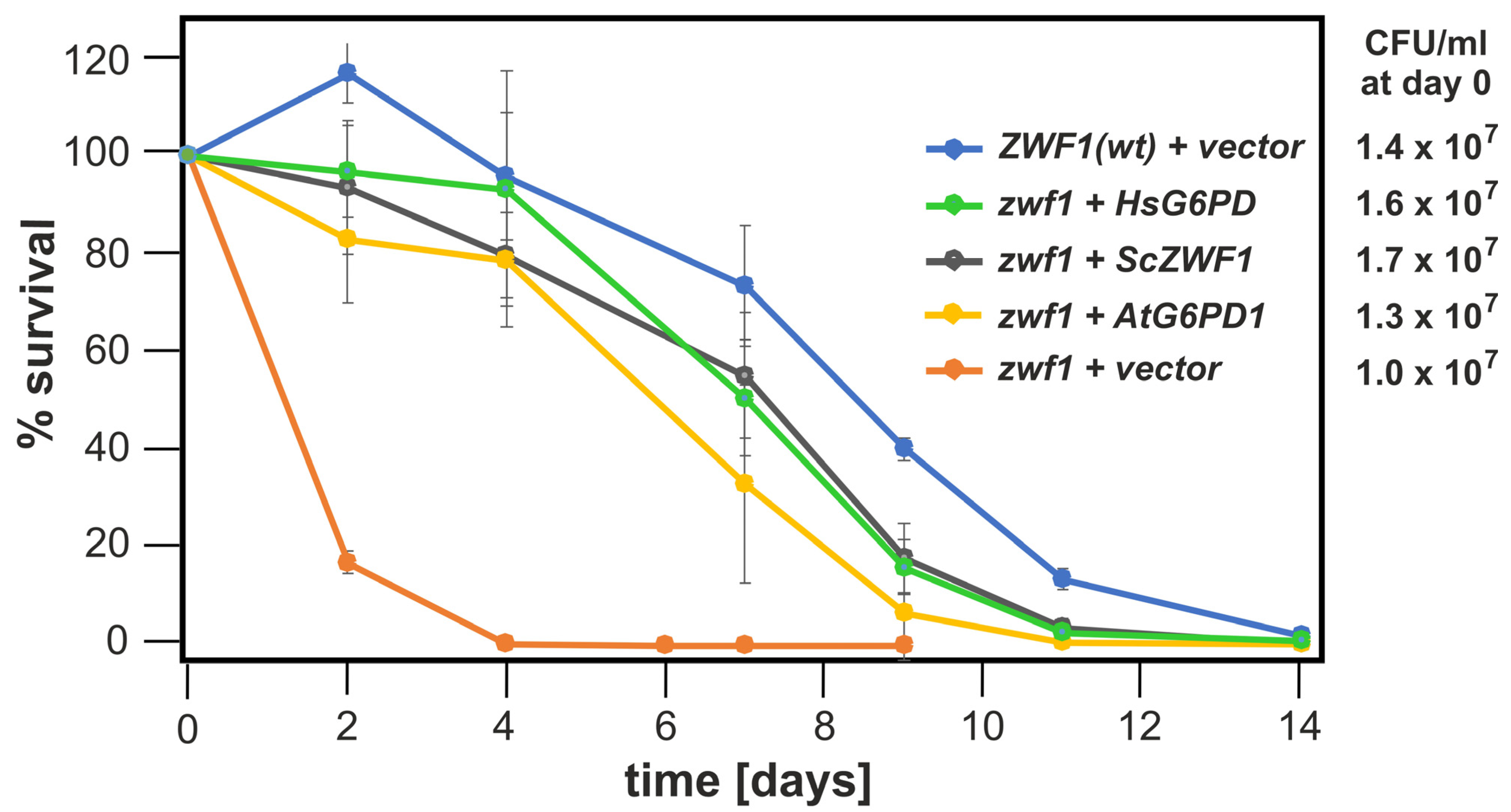
3.4. Chronological Life Span of S. cerevisiae Is Drastically Reduced by the zwf1 Deletion and Restored by Heterologous Expression of Functional ZWF1 Homologs
4. Discussion
5. Conlcusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CLS | chronological lifespan |
| G6PD | glucose-6-phosphate dehydrogenase |
| DTT | dithiothreitol |
| PPP | pentose phosphate pathway |
| ZWF1 | “Zwischenferment”; a synonym to G6PD from early metabolic analyses performed in the first half of last century |
References
- Ali, S.S.; Ahsan, H.; Zia, M.K.; Siddiqui, T.; Khan, F.H. Understanding oxidants and antioxidants: Classical team with new players. J. Food Biochem. 2020, e13145. [Google Scholar] [CrossRef] [PubMed]
- Pugh, C.W.; Ratcliffe, P.J. New horizons in hypoxia signaling pathways. Exp. Cell Res. 2017, 356, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.C.; Wu, Y.H.; Liu, H.Y.; Stern, A.; Chiu, D.T. What has passed is prolog: New cellular and physiological roles of G6PD. Free Radic. Res. 2016, 50, 1047–1064. [Google Scholar] [CrossRef] [PubMed]
- Campbell, K.; Vowinckel, J.; Keller, M.A.; Ralser, M. Methionine metabolism alters oxidative stress resistance via the pentose phosphate pathway. Antioxid. Redox. Signal 2016, 24, 543–547. [Google Scholar] [CrossRef] [Green Version]
- Grabowska, D.; Jablonska-Skwiecinska, E.; Plochocka, D.; Chelstowska, A.; Lewandowska, I.; Witos, I.; Majewska, Z.; Rokicka-Milewska, R.; Burzynska, B. A novel mutation in the glucose-6-phosphate dehydrogenase gene in a subject with chronic nonspherocytic hemolytic anemia--characterization of enzyme using yeast expression system and molecular modeling. Blood Cells Mol. Dis. 2004, 32, 124–130. [Google Scholar] [CrossRef]
- Mehta, A.; Mason, P.J.; Vulliamy, T.J. Glucose-6-phosphate dehydrogenase deficiency. Best Pract. Res. Clin. Haematol. 2000, 13, 21–38. [Google Scholar] [CrossRef] [Green Version]
- Patra, K.C.; Hay, N. The pentose phosphate pathway and cancer. Trends Biochem. Sci. 2014, 39, 347–354. [Google Scholar] [CrossRef] [Green Version]
- Riganti, C.; Gazzano, E.; Polimeni, M.; Aldieri, E.; Ghigo, D. The pentose phosphate pathway: An antioxidant defense and a crossroad in tumor cell fate. Free Radic. Biol. Med. 2012, 53, 421–436. [Google Scholar] [CrossRef]
- Ho, H.Y.; Cheng, M.L.; Chiu, D.T. Glucose-6-phosphate dehydrogenase—Beyond the realm of red cell biology. Free Radic. Res. 2014, 48. [Google Scholar] [CrossRef]
- Jain, M.; Brenner, D.A.; Cui, L.; Lim, C.C.; Wang, B.; Pimentel, D.R.; Koh, S.; Sawyer, D.B.; Leopold, J.A.; Handy, D.E.; et al. Glucose-6-phosphate dehydrogenase modulates cytosolic redox status and contractile phenotype in adult cardiomyocytes. Circ. Res. 2003, 93, e9–e16. [Google Scholar] [CrossRef] [Green Version]
- Ruwende, C.; Khoo, S.C.; Snow, R.W.; Yates, S.N.; Kwiatkowski, D.; Gupta, S.; Warn, P.; Allsopp, C.E.; Gilbert, S.C.; Peschu, N.; et al. Natural selection of hemi- and heterozygotes for G6PD deficiency in Africa by resistance to severe malaria. Nature 1995, 376, 246–249. [Google Scholar] [CrossRef] [PubMed]
- Preuss, J.; Jortzik, E.; Becker, K. Glucose-6-phosphate metabolism in Plasmodium falciparum. IUBMB Life 2012, 64, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Wakao, S.; Benning, C. Genome-wide analysis of glucose-6-phosphate dehydrogenases in Arabidopsis. Plant J. 2005, 41, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Meyer, T.; Hölscher, C.; Schwöppe, C.; von Schaewen, A. Alternative targeting of Arabidopsis plastidic glucose-6-phosphate dehydrogenase G6PD1 involves cysteine-dependent interaction with G6PD4 in the cytosol. Plant J. 2011, 66, 745–758. [Google Scholar] [CrossRef]
- Graeve, K.; von Schaewen, A.; Scheibe, R. Purification, characterization, and cDNA sequence of glucose-6-phosphate dehydrogenase from potato (Solanum tuberosum L). Plant J. 1994, 5, 353–361. [Google Scholar] [CrossRef]
- Neé, G.; Aumont-Nicaise, M.; Zaffagnini, M.; Nessler, S.; Valerio-Lepiniec, M.; Issakidis-Bourguet, E. Redox regulation of chloroplastic G6PDH activity by thioredoxin occurs through structural changes modifying substrate accessibility and cofactor binding. Biochem. J. 2014, 457. [Google Scholar] [CrossRef]
- Wenderoth, I.; Scheibe, R.; von Schaewen, A. Identification of the cysteine residues involved in redox modification of plant plastidic glucose-6-phosphate dehydrogenase. J. Biol. Chem. 1997, 272, 26985–26990. [Google Scholar] [CrossRef] [Green Version]
- Scheibe, R.; Geissler, A.; Fickenscher, K. Chloroplast glucose-6-phosphate dehydrogenase: Km shift upon light modulation and reduction. Arch. Biochem. Biophys. 1989, 274, 290–297. [Google Scholar] [CrossRef]
- Knüsting, J.; Scheibe, R. Small molecules govern thiol redox switches. Trends Plant. Sci. 2018, 23, 769–782. [Google Scholar] [CrossRef]
- Von Schaewen, A.; Langenkamper, G.; Graeve, K.; Wenderoth, I.; Scheibe, R. Molecular characterization of the plastidic glucose-6-phosphate dehydrogenase from potato in comparison to its cytosolic counterpart. Plant Physiol. 1995, 109, 1327–1335. [Google Scholar] [CrossRef] [Green Version]
- Cardi, M.; Zaffagnini, M.; De Lillo, A.; Castiglia, D.; Chibani, K.; Gualberto, J.M.; Rouhier, N.; Jacquot, J.P.; Esposito, S. Plastidic P2 glucose-6P dehydrogenase from poplar is modulated by thioredoxin m-type: Distinct roles of cysteine residues in redox regulation and NADPH inhibition. Plant Sci. 2016, 252, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Landi, S.; Nurcato, R.; De Lillo, A.; Lentini, M.; Grillo, S.; Esposito, S. Glucose-6-phosphate dehydrogenase plays a central role in the response of tomato (Solanum lycopersicum) plants to short and long-term drought. Plant Physiol. Biochem. 2016, 105, 79–89. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Ma, Y.; Huang, C.; Li, J.; Wan, Q.; Bi, Y. Involvement of glucose-6-phosphate dehydrogenase in reduced glutathione maintenance and hydrogen peroxide signal under salt stress. Plant Signal Behav. 2008, 3, 394–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Fu, Z.; Su, Y.; Zhang, X.; Li, G.; Guo, J.; Que, Y.; Xu, L. A cytosolic glucose-6-phosphate dehydrogenase gene, ScG6PDH, plays a positive role in response to various abiotic stresses in sugarcane. Sci. Rep. 2014, 4, 7090. [Google Scholar] [CrossRef] [PubMed]
- Debnam, P.M.; Fernie, A.R.; Leisse, A.; Golding, A.; Bowsher, C.G.; Grimshaw, C.; Knight, J.S.; Emes, M.J. Altered activity of the P2 isoform of plastidic glucose 6-phosphate dehydrogenase in tobacco (Nicotiana tabacum cv. Samsun) causes changes in carbohydrate metabolism and response to oxidative stress in leaves. Plant J. 2004, 38, 49–59. [Google Scholar] [CrossRef]
- Scharte, J.; Schon, H.; Tjaden, Z.; Weis, E.; von Schaewen, A. Isoenzyme replacement of glucose-6-phosphate dehydrogenase in the cytosol improves stress tolerance in plants. Proc. Natl. Acad. Sci. USA 2009, 106, 8061–8066. [Google Scholar] [CrossRef] [Green Version]
- Bussell, J.D.; Keech, O.; Fenske, R.; Smith, S.M. Requirement for the plastidial oxidative pentose phosphate pathway for nitrate assimilation in Arabidopsis. Plant J. 2013, 75, 578–591. [Google Scholar] [CrossRef]
- Esposito, S. Nitrogen assimilation, abiotic stress and glucose 6-phosphate dehydrogenase: The full circle of reductants. Plants 2016, 5. [Google Scholar] [CrossRef]
- Lobo, Z.; Maitra, P.K. Pentose phosphate pathway mutants of yeast. Mol. Gen. Genet. 1982, 185, 367–368. [Google Scholar] [CrossRef]
- Nogae, I.; Johnston, M. Isolation and characterization of the ZWF1 gene of Saccharomyces cerevisiae, encoding glucose-6-phosphate dehydrogenase. Gene 1990, 96, 161–169. [Google Scholar] [CrossRef]
- Thomas, D.; Cherest, H.; Surdin-Kerjan, Y. Identification of the structural gene for glucose-6-phosphate dehydrogenase in yeast. Inactivation leads to a nutritional requirement for organic sulfur. EMBO J. 1991, 10, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Krüger, A.; Gruning, N.M.; Wamelink, M.M.; Kerick, M.; Kirpy, A.; Parkhomchuk, D.; Bluemlein, K.; Schweiger, M.R.; Soldatov, A.; Lehrach, H.; et al. The pentose phosphate pathway is a metabolic redox sensor and regulates transcription during the antioxidant response. Antioxid. Redox. Signal 2011, 15, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Stincone, A.; Prigione, A.; Cramer, T.; Wamelink, M.M.; Campbell, K.; Cheung, E.; Olin-Sandoval, V.; Gruning, N.M.; Kruger, A.; Tauqeer Alam, M.; et al. The return of metabolism: Biochemistry and physiology of the pentose phosphate pathway. Biol. Rev. Camb. Philos. Soc. 2015, 90, 927–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagunas, R.; Gancedo, J.M. Reduced pyridine-nucleotides balance in glucose-growing Saccharomyces cerevisiae. Eur. J. Biochem. 1973, 37, 90–94. [Google Scholar] [CrossRef]
- Jacoby, J.; Hollenberg, C.P.; Heinisch, J.J. Transaldolase mutants in the yeast Kluyveromyces lactis provide evidence that glucose can be metabolized through the pentose phosphate pathway. Mol. Microbiol. 1993, 10, 867–876. [Google Scholar] [CrossRef]
- Rodicio, R.; Heinisch, J.J. Yeast on the milky way: Genetics, physiology and biotechnology of Kluyveromyces lactis. Yeast 2013, 30, 165–177. [Google Scholar] [CrossRef]
- Ralser, M.; Wamelink, M.M.; Kowald, A.; Gerisch, B.; Heeren, G.; Struys, E.A.; Klipp, E.; Jakobs, C.; Breitenbach, M.; Lehrach, H.; et al. Dynamic rerouting of the carbohydrate flux is key to counteracting oxidative stress. J. Biol. 2007, 6. [Google Scholar] [CrossRef] [Green Version]
- Dick, T.P.; Ralser, M. Metabolic Remodeling in Times of Stress: Who Shoots Faster than His Shadow? Mol. Cell 2015, 59, 519–521. [Google Scholar] [CrossRef] [Green Version]
- Kuehne, A.; Emmert, H.; Soehle, J.; Winnefeld, M.; Fischer, F.; Wenck, H.; Gallinat, S.; Terstegen, L.; Lucius, R.; Hildebrand, J.; et al. Acute activation of oxidative pentose phosphate pathway as first-line response to oxidative stress in human skin cells. Mol. Cell 2015, 59, 359–371. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, A.; Lin, M. Regulation of glucose-6-phosphate dehydrogenase activity in red blood cells from hemolytic and nonhemolytic variant subjects. Blood 1973, 41, 877–891. [Google Scholar] [CrossRef]
- Hildebrandt, T.; Knuesting, J.; Berndt, C.; Morgan, B.; Scheibe, R. Cytosolic thiol switches regulating basic cellular functions: GAPDH as an information hub? Biol. Chem. 2015, 396, 523–537. [Google Scholar] [CrossRef] [PubMed]
- Peralta, D.; Bronowska, A.K.; Morgan, B.; Doka, E.; Van Laer, K.; Nagy, P.; Grater, F.; Dick, T.P. A proton relay enhances H2O2 sensitivity of GAPDH to facilitate metabolic adaptation. Nat. Chem. Biol. 2015, 11, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Arvanitidis, A.; Heinisch, J.J. Studies on the function of yeast phosphofructokinase subunits by in vitro mutagenesis. J. Biol. Chem. 1994, 269, 8911–8918. [Google Scholar] [PubMed]
- Kirchrath, L.; Lorberg, A.; Schmitz, H.P.; Gengenbacher, U.; Heinisch, J.J. Comparative genetic and physiological studies of the MAP kinase Mpk1p from Kluyveromyces lactis and Saccharomyces cerevisiae. J. Mol. Biol. 2000, 300, 743–758. [Google Scholar] [CrossRef]
- Schacherer, J.; Ruderfer, D.M.; Gresham, D.; Dolinski, K.; Botstein, D.; Kruglyak, L. Genome-wide analysis of nucleotide-level variation in commonly used Saccharomyces cerevisiae strains. PLoS ONE 2007, 2, e322. [Google Scholar] [CrossRef] [Green Version]
- Rose, M.D.; Winston, F.; Hieter, P. Methods in Yeast Genetics; Cold Harbor Spring Laboratory: New York, NY, USA, 1990. [Google Scholar]
- Schmitz, H.P.; Jendretzki, A.; Wittland, J.; Wiechert, J.; Heinisch, J.J. Identification of Dck1 and Lmo1 as upstream regulators of the small GTPase Rho5 in Saccharomyces cerevisiae. Mol. Microbiol. 2015, 96, 306–324. [Google Scholar] [CrossRef] [Green Version]
- Rothstein, R. Targeting, disruption, replacement, and allele rescue: Integrative DNA transformation in yeast. Methods Enzymol. 1991, 194, 281–301. [Google Scholar]
- Brachmann, C.B.; Davies, A.; Cost, G.J.; Caputo, E.; Li, J.; Hieter, P.; Boeke, J.D. Designer deletion strains derived from Saccharomyces cerevisiae S288C: A useful set of strains and plasmids for PCR-mediated gene disruption and other applications. Yeast 1998, 14, 115–132. [Google Scholar] [CrossRef]
- Sherman, F.; Fink, G.R.; Hicks, J.B. Laboratory Course Manual for Methods in Yeast Genetics; Cold Spring Harbour Laboratory: New York, NY, USA, 1986. [Google Scholar]
- Schneider, M.; Knuesting, J.; Birkholz, O.; Heinisch, J.J.; Scheibe, R. Cytosolic GAPDH as a redox-dependent regulator of energy metabolism. BMC Plant Biol. 2018, 18, 184. [Google Scholar] [CrossRef] [Green Version]
- Longtine, M.S.; McKenzie, A., 3rd; Demarini, D.J.; Shah, N.G.; Wach, A.; Brachat, A.; Philippsen, P.; Pringle, J.R. Additional modules for versatile and economical PCR-based gene deletion and modification in Saccharomyces cerevisiae. Yeast 1998, 14, 953–961. [Google Scholar] [CrossRef]
- Rodicio, R.; Koch, S.; Schmitz, H.P.; Heinisch, J.J. KlRHO1 and KlPKC1 are essential for cell integrity signalling in Kluyveromyces lactis. Microbiology 2006, 152, 2635–2649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kock, C.; Arlt, H.; Ungermann, C.; Heinisch, J.J. Yeast cell wall integrity sensors form specific plasma membrane microdomains important for signalling. Cell Microbiol. 2016, 18, 1251–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinisch, J. Construction and physiological characterization of mutants disrupted in the phosphofructokinase genes of Saccharomyces cerevisiae. Curr. Genet. 1986, 11, 227–234. [Google Scholar] [CrossRef]
- Schaaff-Gerstenschlager, I.; Zimmermann, F.K. Pentose-phosphate pathway in Saccharomyces cerevisiae: Analysis of deletion mutants for transketolase, transaldolase, and glucose 6-phosphate dehydrogenase. Curr. Genet. 1993, 24, 373–376. [Google Scholar] [CrossRef]
- Dowell, R.D.; Ryan, O.; Jansen, A.; Cheung, D.; Agarwala, S.; Danford, T.; Bernstein, D.A.; Rolfe, P.A.; Heisler, L.E.; Chin, B.; et al. Genotype to phenotype: A complex problem. Science 2010, 328, 469. [Google Scholar] [CrossRef] [Green Version]
- Miosga, T.; Zimmermann, F.K. Cloning and characterization of the first two genes of the non-oxidative part of the Saccharomyces cerevisiae pentose-phosphate pathway. Curr. Genet. 1996, 30, 404–409. [Google Scholar] [CrossRef]
- Zimmermann, A.; Hofer, S.; Pendl, T.; Kainz, K.; Madeo, F.; Carmona-Gutierrez, D. Yeast as a tool to identify anti-aging compounds. FEMS Yeast Res. 2018, 18. [Google Scholar] [CrossRef]
- Levy, H.R.; Vought, V.E.; Yin, X.; Adams, M.J. Identification of an arginine residue in the dual coenzyme-specific glucose-6-phosphate dehydrogenase from Leuconostoc mesenteroides that plays a key role in binding NADP+ but not NAD+. Arch. Biochem. Biophys. 1996, 326, 145–151. [Google Scholar] [CrossRef]
- Vought, V.; Ciccone, T.; Davino, M.H.; Fairbairn, L.; Lin, Y.; Cosgrove, M.S.; Adams, M.J.; Levy, H.R. Delineation of the roles of amino acids involved in the catalytic functions of Leuconostoc mesenteroides glucose 6-phosphate dehydrogenase. Biochemistry 2000, 39, 15012–15021. [Google Scholar] [CrossRef]
- Nissen, T.L.; Anderlund, M.; Nielsen, J.; Villadsen, J.; Kielland-Brandt, M.C. Expression of a cytoplasmic transhydrogenase in Saccharomyces cerevisiae results in formation of 2-oxoglutarate due to depletion of the NADPH pool. Yeast 2001, 18, 19–32. [Google Scholar] [CrossRef]
- Jeppsson, M.; Johansson, B.; Hahn-Hagerdal, B.; Gorwa-Grauslund, M.F. Reduced oxidative pentose phosphate pathway flux in recombinant xylose-utilizing Saccharomyces cerevisiae strains improves the ethanol yield from xylose. Appl. Environ. Microbiol. 2002, 68, 1604–1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorsich, S.W.; Dien, B.S.; Nichols, N.N.; Slininger, P.J.; Liu, Z.L.; Skory, C.D. Tolerance to furfural-induced stress is associated with pentose phosphate pathway genes ZWF1, GND1, RPE1, and TKL1 in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2006, 71, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, E.; Gao, K.; Mercurio, K.; Ali, M.; Baetz, K. Yeast chemogenomic screen identifies distinct metabolic pathways required to tolerate exposure to phenolic fermentation inhibitors ferulic acid, 4-hydroxybenzoic acid and coniferyl aldehyde. Metab. Eng. 2019, 52, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Kwak, S.; Yun, E.J.; Lane, S.; Oh, E.J.; Kim, K.H.; Jin, Y.S. Redirection of the glycolytic flux enhances isoprenoid production in Saccharomyces cerevisiae. Biotechnol. J. 2020, 15, e1900173. [Google Scholar] [CrossRef]
- Zhao, X.; Shi, F.; Zhan, W. Overexpression of ZWF1 and POS5 improves carotenoid biosynthesis in recombinant Saccharomyces cerevisiae. Lett. Appl. Microbiol. 2015, 61, 354–360. [Google Scholar] [CrossRef]
- Cardenas, J.; Da Silva, N.A. Engineering cofactor and transport mechanisms in Saccharomyces cerevisiae for enhanced acetyl-CoA and polyketide biosynthesis. Metab. Eng. 2016, 36, 80–89. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name (Number) | Sequence (5′ → 3′) |
|---|---|
| ZWF1atgforBam (16.234) | ggcgggatccATGAGTGAAGGCCCCGTCAAATTCG |
| ZWF1revHind (17.036) | gcgtgaaagcttGATAAGTACAAGTCCAATCGGACTG |
| KlZWF1ATGforBam (17.309) | ctacaggatccATGGCTACTCAGTTTGACGAGAAC |
| KlZWF1revwoBam (18.002) | acacaggaaacagctatgaccatgattacgccaagcttTTACATTTTAGGAGTGGTGACAGGCCATTGGTAGctTCCTGGTTTGGAAAAGG |
| 2064rev3HA (17.046) | acaatttcacacaggaaacagctatgaccatgattacgccaagcttAGGGAGACCGGCAGATCCGCGG |
| ScZWF1for3HA (18.200) | ttacgcttggcccgtgactaagccagaagatacgaaggataatCGGATCCCCGGGTTAATTAA |
| KlZWF1for3HA (18.202) | aaaccaggaagctaccaatggcctgtcaccactcctaaaatgCGGATCCCCGGGTTAATTAA |
| HsG6PD1for3HA (18.205) | ttcaatacgagggtacttacaaatgggttaatccacacaagctgCGGATCCCCGGGTTAATTAA |
| EcZWF1for3HA (18.201) | gatgattacccgtgatggtcgttcctggaatgagtttgagCGGATCCCCGGGTTAATTAA |
| AtG6PD1for3HA (18.199) | ttctaagtataacgttagatggggtgacttgggtgaagcaCGGATCCCCGGGTTAATTAA |
| AtG6PD5for3HA (18.203) | tacatgcaaacccatggttacatttggattccaccaactttgCGGATCCCCGGGTTAATTAA |
| AtG6PD6for3HA (18.204) | ttacttgcaaacccatggttatatttggattccaccaaccttgCGGATCCCCGGGTTAATTAA |
| Sczwf1del5 (16.235) | atgagtgaaggccccgtcaaattcgaaaaaaataccgtcatatCTTCGTACGCTGCAGGTCGAC |
| Sczwf1del3 (16.236) | ctaattatccttcgtatcttctggcttagtcacgggccaaGCATAGGCCACTAGTGGATCTG |
| ScZWF1forSac (16.232) | gcgtgagctCCTGGTAAGTAAGGTGTAGTTTTG |
| ScZWF1revSal (16.233) | gtgagtcgacGATAAGTACAAGTCCAATCGGACTG |
| Scrpe1del5 (19.139) | aagaaggccatttgctaattccaagagcgaggtaaacacacaagaaaaaCTTCGTACGCTGCAGGTCGAC |
| Scrpe1del3 (19.140) | tatcgtatagtatagagagtataaatataagaaatgccgcatatgtacaaGCATAGGCCACTAGTGGATCTG |
| ScRPE1forBam (19.137) | CTCGTGgatCCAATAATGAAACTGAAAAGCATG |
| ScRPE1revHind (19.138) | AAAGAAGcTTCTTTGACTTTGGTTAAGG |
| Scnqm1del5 (19.143) | cgtaagtcataaaaaataggaaataatcacatatatacaagaaattaaatCTTCGTACGCTGCAGGTCGAC |
| Scnqm1del3 (19.144) | tggtatatatatatttatatatataagtaggtacctctactcttaatgaGCATAGGCCACTAGTGGATCTG |
| ScNQM1forXho (19.141) | GCAATCTCGAGAACAATTGCAGGACAGG |
| ScNQM1revSac (19.142) | gtacggagcTCGGAATTTGATTATACGTCAG |
| Strain/Plasmid | Gene | Source Organism | Accession Number 1 | Specific Activity 2 [mU/mg Protein] |
|---|---|---|---|---|
| HD56-5A/pJJH2064 | ZWF1 (wt) | Saccharomyces cerevisiae | CP046094.1 | 99.33 ± 2.47 |
| GI:1789112053 | ||||
| HOD269-1C/pJJH2064 | vector | - | - | < 0.5 |
| HOD269-1C/pJJH2111 | ScZWF1 | Saccharomyces cerevisiae (strain CEN.PK113-7D) | CP046094.1 | 312.92 ± 26.60 |
| GI:1789112053 | ||||
| HOD269-1C/pJJH2292 | KlZWF1 | Kluyveromyces lactis | NC_006040.1 | 134.43 ± 14.67 |
| GI: 50313009 | ||||
| HOD269-1C/pJJH2223 | HsG6PD1 | Homo sapiens | BC000337.2 | 89.03 ± 5.86 |
| GI: 33991065 | ||||
| HOD269-1C/pJJH2125 | AtG6PD13 | Arabidopsis thaliana | NM_122970.6 GI:1063734559 | 12.78 ± 1.43 |
| HOD269-1C/pJJH2224 | AtG6PD5 | Arabidopsis thaliana | NM_113644.5 | 19.38 ± 4.88 |
| GI: 1063714071 | ||||
| HOD269-1C/pJJH2249 | AtG6PD6 | Arabidopsis thaliana | NM_113644.5 | 24.38 ± 4.96 |
| GI: 1063714071 | ||||
| HOD269-1C/pJJH2494 | EcZWF1 | Escherichia coli K12 | CP047127.1 | 20.28 ± 1.05 |
| GI: 1789840096 | ||||
| HOD269-1C/pJJH2495 | LmZWF1 | Leuconostoc mesenteroides | M64446.1 | 52.01 ± 1.07 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heinisch, J.J.; Knuesting, J.; Scheibe, R. Investigation of Heterologously Expressed Glucose-6-Phosphate Dehydrogenase Genes in a Yeast zwf1 Deletion. Microorganisms 2020, 8, 546. https://doi.org/10.3390/microorganisms8040546
Heinisch JJ, Knuesting J, Scheibe R. Investigation of Heterologously Expressed Glucose-6-Phosphate Dehydrogenase Genes in a Yeast zwf1 Deletion. Microorganisms. 2020; 8(4):546. https://doi.org/10.3390/microorganisms8040546
Chicago/Turabian StyleHeinisch, Jürgen J., Johannes Knuesting, and Renate Scheibe. 2020. "Investigation of Heterologously Expressed Glucose-6-Phosphate Dehydrogenase Genes in a Yeast zwf1 Deletion" Microorganisms 8, no. 4: 546. https://doi.org/10.3390/microorganisms8040546
APA StyleHeinisch, J. J., Knuesting, J., & Scheibe, R. (2020). Investigation of Heterologously Expressed Glucose-6-Phosphate Dehydrogenase Genes in a Yeast zwf1 Deletion. Microorganisms, 8(4), 546. https://doi.org/10.3390/microorganisms8040546