Advances in Molecular Tools and In Vivo Models for the Study of Human Fungal Pathogenesis


Abstract
:1. Introduction
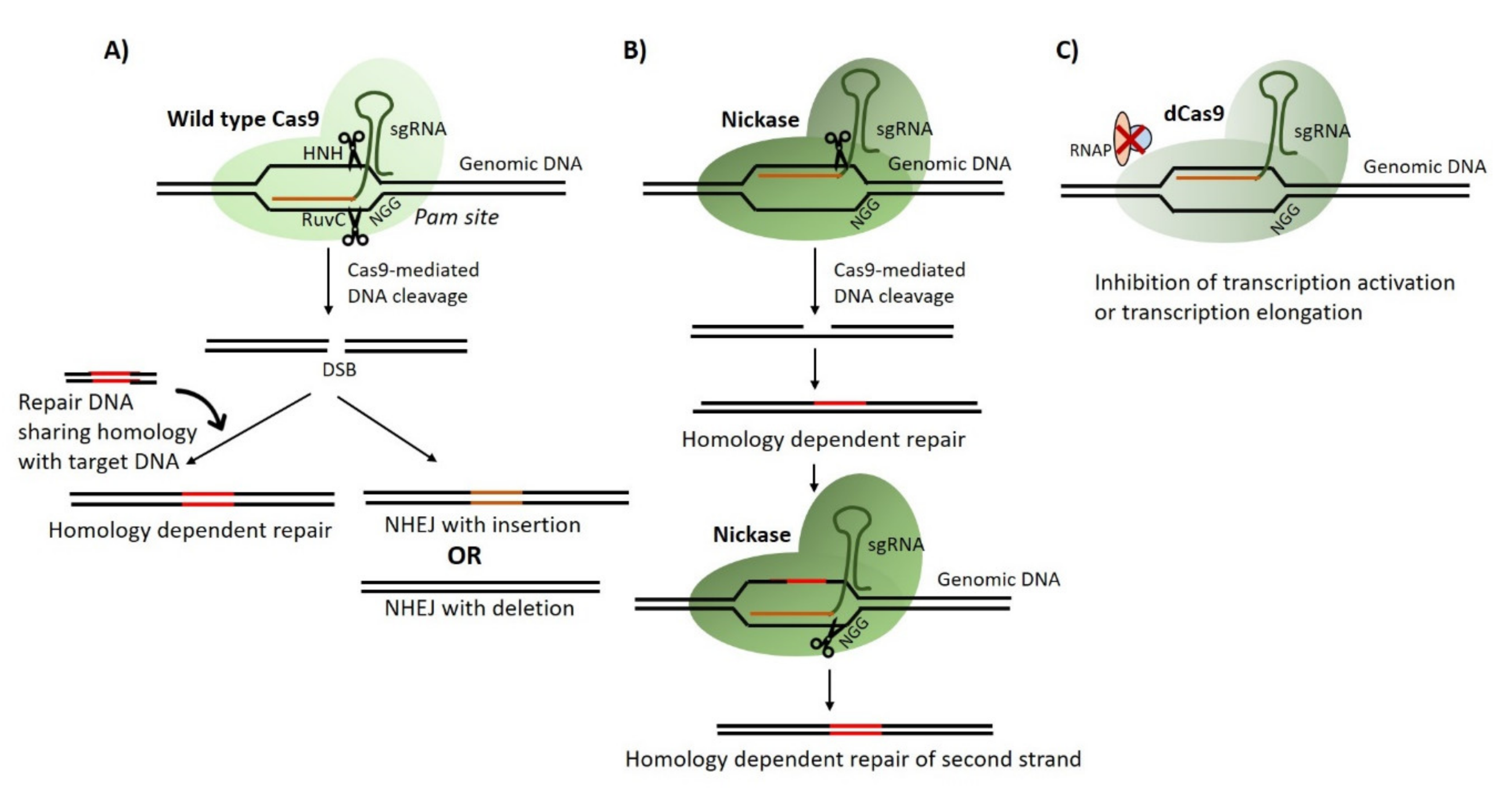
2. CRISPR/Cas Gene Editing
3. RNAi
4. Transposable Elements
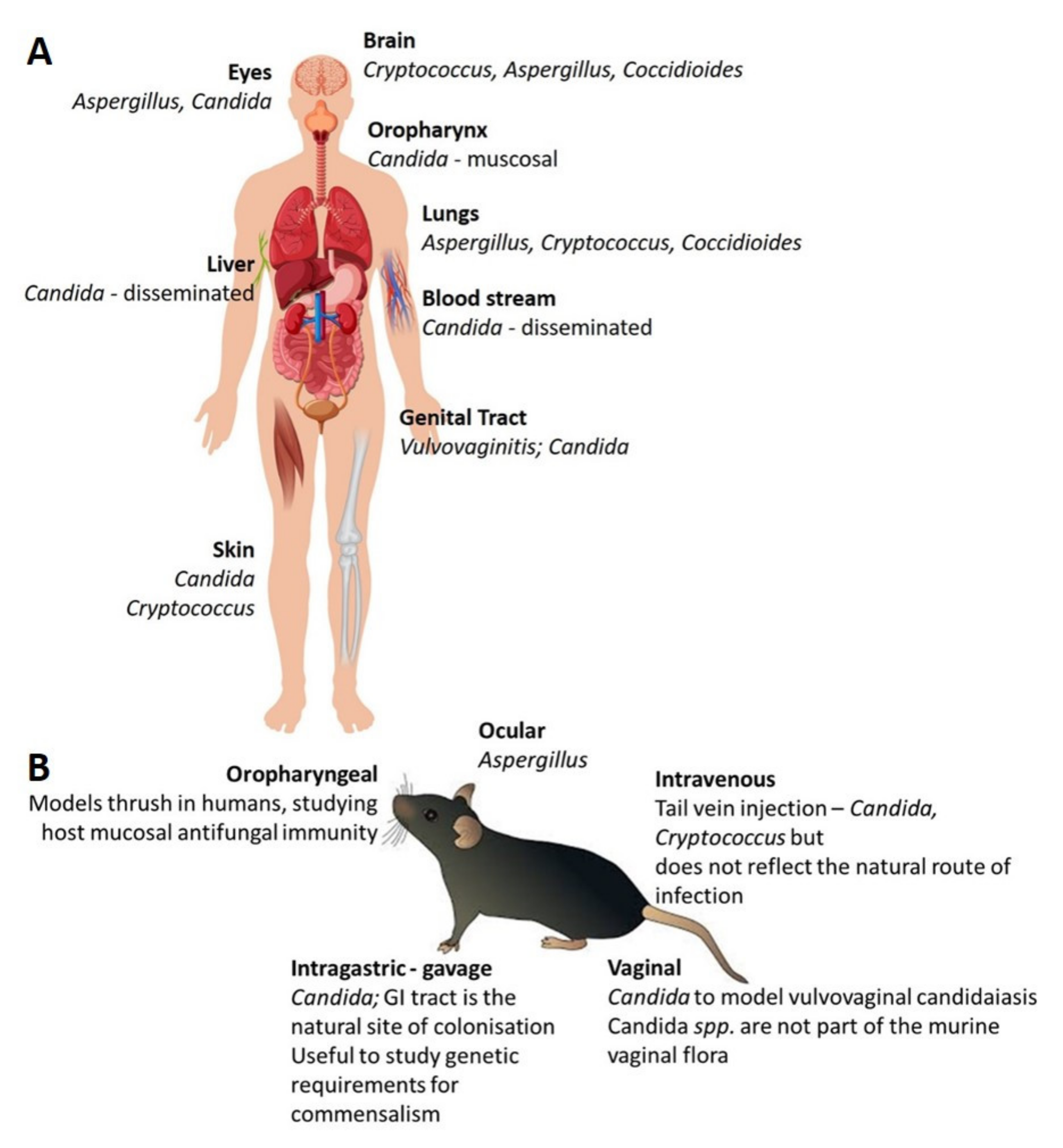
5. Animal Models for Studying Human Fungal Pathogens
5.1. Zebrafish
5.2. Bombyx mori—The Silkworm
5.3. Galleria mellonella—The Wax Moth
5.4. Murine Model—Mus musculus
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bongomin, F.; Gago, S.; Oladele, R.; Denning, D.W. Global and multi-national prevalence of fungal disease- estimate precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef] [PubMed]
- Kohler, J.R.; Casadevall, A.; Perfect, J. The spectrum of fungi that infects humans. Cold Spring Harb. Perspect. Med. 2015, 5, a019273. [Google Scholar] [CrossRef] [Green Version]
- Brown, G.D.; Denning, D.W.; Gow, N.A.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden Killer: Human Fungal infections. Sci. Transl. Med. 2012, 4, 165rv13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denning, D.W. Global Fungal Burden. Mycoses 2013, 56, 13. [Google Scholar]
- Jain, A.; Jain, S.; Rawat, S. Emerging fungal infections among children: A review on its clinical manifestations, diagnosis and preventation. J. Pharm. Bioallied. Sci. 2010, 2, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Doudna, J.A.; Charpentier, E. The new frontier of genome engineering with CRISPR-Cas9. Science (80-) 2014, 346, 1258096. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA – Guided. Science 2012, 337, 816–822. [Google Scholar]
- Zetsche, B.; Gootenberg, J.S.; Abudayyeh, O.O.; Slaymaker, I.M.; Makarova, K.S.; Essletzbichler, P.; Volz, S.E.; Joung, J.; Van Der Oost, J.; Regev, A.; et al. Cpf1 Is a Single RNA-Guided Endonuclease of a Class 2 CRISPR-Cas System. Cell 2015, 163, 759–771. [Google Scholar] [CrossRef] [Green Version]
- Hryhorowicz, M.; Lipin, D. CRISPR/Cas9 Immune System as a Tool for Genome Engineering. Arch. Immunol. Et Ther. Exp. 2017, 65, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Nishida, K.; Arazoe, T.; Yachie, N.; Banno, S.; Kakimoto, M.; Tabata, M.; Mochizuki, M.; Miyabe, A.; Araki, M.; Hara, K.Y.; et al. Targeted nucleotide editing using hybrid prokaryotic and vertebrate adaptive immune systems. Science (80-) 2016, 353, aaf8729. [Google Scholar] [CrossRef]
- Umeyama, T.; Hayashi, Y.; Shimosaka, H.; Inukai, T.; Yamagoe, S.; Takatsuka, S.; Hoshino, Y.; Nagi, M.; Nakamura, S.; Kamei, K.; et al. crossm CRISPR/Cas9 Genome Editing to Demonstrate the Contribution of Cyp51A Gly138Ser to Azole Resistance in Aspergillus fumigatus. Antimicrob. Agents Chemother. 2018, 62, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahim, K.; Huo, L.; Li, C.; Zhang, P.; Basit, A.; Xiang, B.; Ting, B.; Hao, X.; Zhu, X. Identification of a basidiomycete-specific Vilse-like GTPase activating proteins (GAPs) and its roles in the production of virulence factors in Cryptococcus neoformans. FEMS Yeast Res. 2017, 17, 1–10. [Google Scholar] [CrossRef]
- Arras, S.D.; Chua, S.M.; Wizrah, M.S.; Faint, J.A.; Yap, A.S.; Fraser, J.A. Targeted genome editing via CRISPR in the pathogen Cryptococcus neoformans. PLoS ONE 2016, 11, e0164322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cen, Y.; Timmermans, B.; Souffriau, B.; Thevelein, J.M.; Van Dijck, P. Comparison of genome engineering using the CRISPR-Cas9 system in C. glabrata wild-type and lig4 strains. Fungal Genet. Biol. 2017, 107, 44–50. [Google Scholar] [CrossRef]
- Zoppo, M.; Lombardi, L.; Rizzato, C.; Lupetti, A.; Bottai, D.; Papp, C.; Gacser, A.; Tavanti, A. CORT0C04210 is required for Candida orthopsilosis adhesion to human buccal cells. Fungal Genet. Biol. 2018, 120, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, L.; Turner, S.A.; Zhao, F.; Butler, G. Gene editing in clinical isolates of Candida parapsilosis using CRISPR/Cas9. Sci Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Meng, X.; Wei, X.; Lu, L. Highly efficient CRISPR mutagenesis by microhomology-mediated end joining in Aspergillus fumigatus. Fungal Genet. Biol. 2016, 86, 47–57. [Google Scholar] [CrossRef]
- Poyntner, C.; Mirastschijski, U.; Sterflinger, K.; Tafer, H. Transcriptome Study of an Exophiala dermatitidis PKS1 Mutant on an ex Vivo Skin Model: Is Melanin Important for Infection? Front. Microbiol. 2018, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Wang, P. Two distinct approaches for CRISPR-Cas9-Mediated gene editing in Cryptococcus neoformans and related species. mSphere 2018, 3, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Kildegaard, K.R.; Tramontin, L.R.; Chekina, K.; Li, M.; Goedecke, T.J.; Kristensen, M.; Borodina, I. CRISPR/Cas9—RNA interference system for combinatorial metabolic engineering of Saccharomyces cerevisiae. Yeast 2019, 35, 237–247. [Google Scholar] [CrossRef] [Green Version]
- Mans, R.; van Rossum, H.M.; Wijsman, M.; Backx, A.; Kuijpers, N.G.; van den Broek, M.; Daran-Lapujade, P.; Pronk, J.T.; van Maris, A.J.; Daran, J.M. CRISPR/Cas9: A molecular Swiss army knife for simultaneous introduction of multiple genetic modifications in Saccharomyces cerevisiae. FEMS Yeast Res. 2015, 15, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Lin, X. Multiple applications of a transient CRISPR-Cas9 coupled with electroporation (TRACE) system in the Ccryptococcus neoformans species complex. Genetics 2018, 208, 1357–1372. [Google Scholar] [CrossRef] [Green Version]
- Nødvig, C.S.; Hoof, J.B.; Kogle, M.E.; Jarczynska, Z.D.; Lehmbeck, J.; Klitgaard, D.K.; Mortensen, U.H. Efficient oligo nucleotide mediated CRISPR-Cas9 gene editing in Aspergilli. Fungal Genet. Biol. 2018, 115, 78–89. [Google Scholar] [CrossRef] [Green Version]
- Sharon, E.; Chen, S.A.; Khosla, N.M.; Smith, J.D.; Pritchard, J.K.; Fraser, H.B. Functional genetic variants revealed by massively parallel precise genome editing. Cell 2019, 175, 544–557. [Google Scholar] [CrossRef] [Green Version]
- Bao, Z.; HamediRad, M.; Xue, P.; Xiao, H.; Tasan, I.; Chao, R.; Liang, J.; Zhao, H. Genome-Scale engineering of Saccharomyces cerevisiae with single-nucleotide precision. Nat. Biotechnol. 2018, 36, 505–508. [Google Scholar] [CrossRef]
- Roy, K.R.; Smith, J.D.; Vonesch, S.C.; Lin, G.; Tu, C.S.; Lederer, A.R.; Chu, A.; Suresh, S.; Nguyen, M.; Horecka, J.; et al. Multiplexed precision genome editing with trackable genomic barcodes in yeast. Nat. Biotechnol. 2018, 36, 512–520. [Google Scholar] [CrossRef]
- Garst, A.D.; Bassalo, M.C.; Pines, G.; A Lynch, S.; Halweg-Edwards, A.L.; Liu, R.; Liang, L.; Wang, Z.; Zeitoun, R.; Alexander, W.G.; et al. Genome-wide mapping of mutations at single-nucleotide resolution for protein, metabolic and genome engineering. Nat. Biotechnol. 2017, 35, 48–55. [Google Scholar] [CrossRef]
- Huang, L.; Dong, H.; Zheng, J.; Wang, B.; Pan, L. Highly efficient single base editing in Aspergillus niger with CRISPR/Cas9 cytidine deaminase fusion. Microbiol Res. 2019, 223–225, 44–50. [Google Scholar] [CrossRef]
- Després, P.C.; Dubé, A.K.; Nielly-Thibault, L.; Yachie, N.; Landry, C.R. Double Selection Enhances the Ef fi ciency of Target-AID and Cas9-Based Genome Editing in Yeast. G3 Genes Genomes Genet. 2018, 8, 3163–3171. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.; Zhang, F.; Karcher, D.; Bock, R. Engineering of high-precision base editors for site-specific single nucleotide replacement. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef]
- Satomura, A.; Nishioka, R.; Mori, H.; Sato, K.; Kuroda, K.; Ueda, M. Precise genome-wide base editing by the CRISPR Nickase system in yeast. Sci. Rep. 2017, 7, 12354. [Google Scholar] [CrossRef] [PubMed]
- Fu, B.X.H.; Smith, J.D.; Fuchs, R.T.; Mabuchi, M.; Curcuru, J.; Robb, G.B.; Fire, A.Z. Target-dependent nickase activities of the CRISPR-Cas nucleases Cpf1 and Cas9. Nat. Microbiol. 2019, 4, 888–897. [Google Scholar] [CrossRef] [PubMed]
- Román, E.; Coman, I.; Prieto, D.; Alonso-Monge, R.; Pla, J. Implementation of a CRISPR-Based System for Gene Regulation in Candida albicans. mSphere 2019, 4, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wensing, L.; Sharma, J.; Uthayakumar, D.; Proteau, Y.; Chavez, A.; Shapiro, R.S. A CRISPR Interference Platform for Efficient Genetic Repression in Candida albicans. mSphere 2019, 4, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Godfray, H.C.J.; North, A.; Burt, A. How driving endonuclease genes can be used to combat pests and disease vectors. BMC Biol. 2017, 15, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gantz, V.M.; Jasinskiene, N.; Tatarenkova, O.; Fazekas, A.; Macias, V.M.; Bier, E.; James, A.A. Highly efficient Cas9-mediated gene drive for population modification of the malaria vector mosquito Anopheles stephensi. Proc. Natl. Acad. Sci. USA 2015, 112, E6736–E6743. [Google Scholar] [CrossRef] [Green Version]
- Champer, J.; Reeves, R.; Oh, S.Y.; Liu, C.; Liu, J.; Clark, A.G.; Messer, P.W. Novel CRISPR/Cas9 gene drive constructs reveal insights into mechanisms of resistance allele formation and drive efficiency in genetically diverse populations. PLoS Genet. 2017, 13, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Grunwald, H.A.; Gantz, V.M.; Poplawski, G.H.D.; Xu, X.-R.S.; Bier, E.; Cooper, K.L. Super-Mendelian inheritance mediated by CRISPR–Cas9 in the female mouse germline. Nature 2019, 566, 105–109. [Google Scholar] [CrossRef]
- Yan, Y.; Finnigan, G.C. Development of a multi-locus CRISPR gene drive system in budding yeast. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Bull, J.J.; Barrick, J.E. Arresting Evolution. Trends Genet. 2017, 33, 910–920. [Google Scholar] [CrossRef]
- Roggenkamp, E.; Giersch, R.M.; Wedeman, E.; Eaton, M.; Turnquist, E.; Schrock, M.N.; Alkotami, L.; Jirakittisonthon, T.; Schluter-Pascua, S.E.; Bayne, G.H.; et al. CRISPR-UnLOCK: Multipurpose Cas9-based strategies for conversion of yeast libraries and strains. Front. Microbiol. 2017, 8, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Roggenkamp, E.; Giersch, R.M.; Schrock, M.N.; Turnquist, E.; Halloran, M.; Finnigan, G.C. Tuning CRISPR-Cas9 gene drives in Saccharomyces cerevisiae. G3 Genes Genomes Genet. 2018, 8, 999–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dicarlo, J.E.; Chavez, A.; Dietz, S.L.; Esvelt, K.M.; Church, G.M. Safeguarding CRISPR-Cas9 gene drives in yeast. Nat. Biotechnol. 2015, 33, 1250–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapiro, R.S.; Chavez, A.; Porter, C.B.M.; Hamblin, M.; Kaas, C.S.; Dicarlo, J.E.; Zeng, G.; Xu, X.; Revtovich, A.V.; Kirienko, N.V.; et al. A CRISPR-Cas9-based gene drive platform for genetic interaction analysis in Candida albicans. Nat. Microbiol. 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halder, V.; Porter, C.B.M.; Chavez, A. Design, execution, and analysis of CRISPR—Cas9-based deletions and genetic interaction networks in the fungal pathogen Candida albicans. Nat. Protoc. 2019, 14, 955–975. [Google Scholar] [CrossRef] [PubMed]
- Goeckel, M.E.; Basgall, E.M.; Lewis, I.C.; Goetting, S.C.; Yan, Y.; Halloran, M.; Finnigan, G.C. Modulating CRISPR gene drive activity through nucleocytoplasmic localization of Cas9 in S. cerevisiae. Fungal Biol. Biotechnol. 2019, 6, 1–11. [Google Scholar] [CrossRef]
- Longmuir, S.; Akhtar, N.; Macneill, S.A. Unexpected insertion of carrier DNA sequences into the fission yeast genome during CRISPR—Cas9 mediated gene deletion. BMC Res. Notes 2019, 12, 1–5. [Google Scholar] [CrossRef]
- Bruni, G.O.; Zhong, K.; Chan, S.; Wang, P. CRISPR-Cas9 induces point mutation in the mucormycosis fungus Rhizopus delemar. Fungal Genet. Biol. 2019, 124, 1–7. [Google Scholar] [CrossRef]
- Ng, H.; Dean, N. Dramatic Improement of CRISPR/Cas9 Editing in Candida albicans by Increased Single Guide RNA Expression. mSphere 2017, 2, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Song, L.; Ouedraogo, J.-P.; Kolbusz, M.; Nguyen, T.T.M.; Tsang, A. Efficient genome editing using tRNA promoter-driven CRISPR/Cas9 gRNA in Aspergillus niger. PLoS ONE 2018, 13, e0202868. [Google Scholar] [CrossRef]
- Verwaal, R.; Buiting-Wiessenhaan, N.; Dalhuijsen, S.; Roubos, J. CRISPR/Cpf1 enables fast and simple genome editing of Saccharomyces cerevisiae. Yeast 2018, 35, 201–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laughery, M.F.; Hunter, T.; Brown, A.; Hoopes, J.; Ostbye, T.; Shumaker, T.; Wyrick, J. New vectors for simple and streamlined CRISPR-Cas9 genome editing in Saccharomyces cerevisiae. Yeast 2015, 32, 711–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dicarlo, J.E.; Norville, J.; Mali, P.; Rios, X.; Aach, J.; Church, G.M. Genome engineering in Saccharomyces cerevisiae using CRISPR-Cas systems. Nucleic Acids Res. 2013, 41, 4336–4343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombardi, L.; Oliveira-Pacheco, J.; Bulter, G. Plasmid-Based CRISPR-Cas9 Gene Editing in Multiple Candida Speices. mSphere 2019, 4, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Vyas, V.K.; Bushkin, G.G.; Bernstein, D.A.; Getz, M.A.; Sewastianik, M.; Barrasa, M.I.; Bartel, D.P.; Fink, G. New CRISPR Mutagenesis Strategies Reveal Variation in Repair Mechanisms among Fungi. mSphere 2018, 3, e00154-18. [Google Scholar] [CrossRef] [Green Version]
- She, W.; Ni, J.; Shui, K.; Wang, F.; He, R.; Xue, J.; Reetz, M.T.; Li, A.; Ma, L. Rapid and Error-Free Site-Directed Mutagenesis by a PCR-Free in Vitro CRISPR/Cas9-Mediated Mutagenic System. ACS Synth. Biol. 2018, 7, 2236–2244. [Google Scholar] [CrossRef]
- Jakociunas, T.; Rajkumar, A.S.; Zhang, J.; Arsovska, D.; Rodriguez, A.; Jendresen, C.B.; Skjødt, M.L.; Nielsen, A.T.; Borodina, I.; Jensen, M.K.; et al. CasEMBLR: Cas9-Facilitated Multiloci Genomic Integration of in Vivo Assembled DNA Parts in Saccharomyces cerevisiae. ACS Synth. Biol. 2015, 4, 1126–1134. [Google Scholar] [CrossRef]
- Apel, A.R.; D’Espaux, L.; Wehrs, M.; Sachs, D.; Li, R.A.; Tong, G.J.; Garber, M.; Nnadi, O.; Zhuang, W.; Hillson, N.J.; et al. A Cas9-based toolkit to program gene expression in Saccharomyces cerevisiae. Nucleic Acids Res. 2017, 45, 496–508. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; He, J.; Wang, Y.; Du, L. A Cloning-Free Method for CRISPR/Cas9-Mediated Genome Editing in Fission Yeast. G3 Genes Genomes Genet. 2018, 8, 2067–2077. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Chavez, A.; Tung, A.; Chan, Y.; Kaas, C.; Yin, Y.; Cecchi, R.; Lopez-Garnier, S.; Kelsic, E.D.; Schubert, M.; et al. High-throughput creation and functional profiling of DNA sequence variant libraries using CRISPR/Cas9 in yeast. Nat. Biotechnol. 2018, 36, 540–546. [Google Scholar] [CrossRef]
- Anzalone, A.V.; Randolph, P.B.; Davis, J.R.; Sousa, A.A.; Koblan, L.W.; Levy, J.M.; Chen, P.J.; Wilson, C.; Newby, G.A.; Raguram, A.; et al. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 2019, 576, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Cox, D.B.T.; Gootenberg, J.S.; Abudayyeh, O.O.; Franklin, B.; Kellner, M.J.; Joung, J.; Zhang, F. RNA editing with CRISPR-Cas13. Science (80-) 2017, 358, 1019–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Hernández, R.J.; Jin, K.; Hernández-Chávez, M.J.; Díaz-Jiménez, D.F.; Trujillo-Esquivel, E.; Clavijo-Giraldo, D.M.; Tamez-Castrellón, A.K.; Franco, B.; Gow, N.A.; Mora-Montes, H.M. Phosphomannosylation and the functional analysis of the extended Candida albicans MNN4-like gene family. Front. Microbiol. 2017, 8, 2156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and Specific Genetic Interference by Double-Stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Zaratiegui, M. RNA Interference Hangs by a Thread. Nature 2015, 520, 162–164. [Google Scholar] [CrossRef]
- Chang, S.; Zhang, Z.; Liu, Y. RNA Interference Pathways in Fungi: Mechanisms and Functions. Annu. Rev. Microbiol. 2012, 66, 305–323. [Google Scholar] [CrossRef] [Green Version]
- Schumann, U.; Ayliffe, M.; Kazan, K.; Wang, M.-B. RNA Silencing in Fungi. Front. Biol. 2010, 5, 478–494. [Google Scholar] [CrossRef]
- Fulci, V.; Macino, G. Quelling: Post-Transcriptional Gene Silencing Guided by Small RNAs in Neurospora Crassa. Curr. Opin. Microbiol. 2007, 10, 199–203. [Google Scholar] [CrossRef]
- Chicas, A.; Cogoni, C.; Macino, G. RNAi-Dependent and RNAi-Independent Mechanisms Contribute to the Silencing of RIPed Sequences in Neurospora Crassa. Nucleic Acids Res. 2004, 32, 4237–4243. [Google Scholar] [CrossRef] [Green Version]
- Song, R.; Zhai, Q.; Sun, L.; Huang, E.; Zhang, Y.; Zhu, Y.; Guo, Q.; Tian, Y. CRISPR/Cas9 Genome Editing Technology in Filamentous Fungi: Progress and Perspective. Appl. Microbiol. Biotechnol. 2019, 103, 6919–6932. [Google Scholar] [CrossRef] [Green Version]
- Martienssen, R.A.; Zaratiegui, M.; Goto, D.B. RNA Interference and Heterochromatin in the Fission Yeast Schizosaccharomyces Pombe. Trends Genet. 2005, 21, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Chang, S.S.; Liu, Y. RNA Interference Pathways in Filamentous Fungi. Cell. Mol. Life Sci. 2010, 67, 3849–3863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, N.; Park, Y.D.; Williamson, P.R. New Technology and Resources for Cryptococcal Research. Physiol. Behav. 2017, 176, 139–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moazeni, M.; Khorramizadeh, M.R.; Teimoori-Toolabi, L.; Noorbakhsh, F.; Fallahi, A.A.; Rezaie, S. Down-Regulation of the ALS3 Gene as a Consequent Effect of RNA-Mediated Silencing of the EFG1 Gene in Candida albicans. Iran. Biomed. J. 2012, 16, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Moazeni, M.; Khoramizadeh, M.R.; Teimoori-Toolabi, L.; Noorbakhsh, F.; Rezaie, S. The Effect of EFG1 Gene Silencing on Down-Regulation of SAP5 Gene, by Use of RNAi Technology. Acta Med. Iran. 2014, 52, 9–14. [Google Scholar] [PubMed]
- De Backer, M.D.; Nelissen, B.; Logghe, M.; Viaene, J.; Loonen, I.; Vandoninck, S.; de Hoogt, R.; Vandoninck, S.; Vanhoof, G. Erratum: An Antisense-Based Functional Genomics Approach for Identification of Genes Critical for Growth of Candida albicans (Nature Biotechnology (2001) 19 (235-241)). Nat. Biotechnol. 2001, 19, 681. [Google Scholar] [CrossRef]
- Janbon, G.; Maeng, S.; Yang, D.H.; Ko, Y.J.; Jung, K.W.; Moyrand, F.; Floyd, A.; Heitman, J.; Bahn, Y.S. Characterizing the Role of RNA Silencing Components in Cryptococcus neoformans. Bone 2011, 23, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Nakayashiki, H.; Nguyen, Q.B. RNA Interference: Roles in Fungal Biology. Curr. Opin. Microbiol. 2008, 11, 494–502. [Google Scholar] [CrossRef]
- Krajaejun, T.; Gauthier, G.M.; Rappleye, C.A.; Sullivan, T.D.; Klein, B.S. Development and Application of a Green Fluorescent Protein Sentinel System for Identification of RNA Interference in Blastomyces dermatitidis Illuminates the Role of Septin in Morphogenesis and Sporulation. Eukaryot. Cell 2007, 6, 1299–1309. [Google Scholar] [CrossRef] [Green Version]
- Chao, H.F.; Yen, Y.F.; Ku, M.S. Characterization of a Salt-Induced DhAHP, a Gene Coding for Alkyl Hydroperoxide Reductase, from the Extremely Halophilic Yeast Debaryomyces hansenii. BMC Microbiol. 2009, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, D.A.; Vyas, V.K.; Weinberg, D.E.; Drinnenberg, I.A.; Bartel, D.P.; Fink, G.R. Candida Albicans Dicer (CaDcr1) Is Required for Efficient Ribosomal and Spliceosomal RNA Maturation. Proc. Natl. Acad. Sci. USA 2012, 109, 523–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Caban, J.; Gonzalez-Velazquez, W.; Perez-Sanchez, L.; Gonzalez-Mendez, R.; Rodriguez-del Valle, N. Calcium/Calmodulin Kinase1 and Its Relation to Thermotolerance and HSP90 in Sporothrix schenckii: An RNAi and Yeast Two-Hybrid Study. BMC Microbiol. 2011, 11, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rappleye, C.A.; Engle, J.T.; Goldman, W.E. RNA Interference in Histoplasma capsulatum Demonstrates a Role for α-(1,3)-Glucan in Virulence. Mol. Microbiol. 2004, 53, 153–165. [Google Scholar] [CrossRef]
- Peres da Silva, R.; Longo, L.G.; da Cunha, J.P.; Sobreira, T.J.; Rodrigues, M.L.; Faoro, H.; Goldenberg, S.; Alves, L.R.; Puccia, R. Comparison of the RNA Content of Extracellular Vesicles Derived from Paracoccidioides brasiliensis and Paracoccidioides lutzii. Cells 2019, 8, 765. [Google Scholar] [CrossRef] [Green Version]
- Skowyra, M.; Doering, T. RNA Interference in Cryptococcus neoformans. Methods Mol. Biol. 2012, 845, 3–17. [Google Scholar] [CrossRef]
- Ishchuk, O.P.; Ahmad, K.M.; Koruza, K.; Bojanovič, K.; Sprenger, M.; Kasper, L.; Brunke, S.; Hube, B.; Säll, T.; Hellmark, T.; et al. RNAi as a Tool to Study Virulence in the Pathogenic Yeast Candida glabrata. Front. Microbiol. 2019, 10, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, I.T.; Ruiz, O.H.; Muñoz, J.F.; Garcia, A.M.; Restrepo, A.; Mcewen, J.G. RNAi Technology Targeting Pb GP 43 and Pb P 27 in Paracoccidioides brasiliensis. Open J. Genet. 2013, 3, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Goes, T.; Bailão, E.F.; Correa, C.R.; Bozzi, A.; Santos, L.I.; Gomes, D.A.; Soares, C.M.; Goes, A.M. New Developments of RNAi in Paracoccidioides brasiliensis: Prospects for High-Throughput, Genome-Wide, Functional Genomics. PLoS Negl. Trop. Dis. 2014, 8. [Google Scholar] [CrossRef] [Green Version]
- Torres-Martínez, S.; Ruiz-Vázquez, R.M. RNAi Pathways in Mucor: A Tale of Proteins, Small RNAs and Functional Diversity. Fungal Genet. Biol. 2016, 90, 44–52. [Google Scholar] [CrossRef] [Green Version]
- Kirkland, T.N.; Muszewska, A.; Stajich, J.E. Analysis of Transposable Elements in Coccidioides Species. J. Fungi 2018, 4, 13. [Google Scholar] [CrossRef] [Green Version]
- Lian, J.; Mishra, S.; Zhao, H. Recent Advances in Metabolic Engineering of Saccharomyces cerevisiae: New Tools and Their Applications. Metab. Eng. 2018, 50, 85–108. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Ding, Q.; Thorpe, J.; Dohmen, R.J.; Keller, J.N. RNA Interference toward UMP1 Induces Proteasome Inhibition in Saccharomyces cerevisiae: Evidence for Protein Oxidation and Autophagic Cell Death. Free Radic. Biol. Med. 2005, 38, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Suk, K.; Choi, J.; Suzuki, Y.; Ozturk, S.B.; Mellor, J.C.; Wong, K.H.; MacKay, J.L.; Gregory, R.I.; Roth, F.P. Reconstitution of Human RNA Interference in Budding Yeast. Nucleic Acids Res. 2011, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, C.; Houseley, J. Endogenous RNA Interference Is Driven by Copy Number. ELife 2014, 2014, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Mercier, R.; Hobman, T.C.; LaPointe, P. Regulation of RNA Interference by Hsp90 Is an Evolutionarily Conserved Process. Biochim. Et Biophys. Acta Mol. Cell Res. 2013, 1833, 2673–2681. [Google Scholar] [CrossRef] [Green Version]
- Kowalik, K.M.; Shimada, Y.; Flury, V.; Stadler, M.B.; Batki, J.; Bühler, M. The Paf1 Complex Represses Small-RNA-Mediated Epigenetic Gene Silencing. Nature 2015, 520, 248–252. [Google Scholar] [CrossRef] [Green Version]
- Si, T.; Luo, Y.; Bao, Z.; Zhao, H. RNAi-Assisted Genome Evolution in Saccharomyces cerevisiae for Complex Phenotype Engineering. ACS Synth. Biol. 2015, 4, 283–291. [Google Scholar] [CrossRef]
- Crook, N.C.; Schmitz, A.C.; Alper, H.S. Alper. Optimization of a Yeast RNA Interference System for Controlling Gene Expression and Enabling Rapid Metabolic Engineering. ACS Synth. Biol. 2014, 3, 307–313. [Google Scholar] [CrossRef]
- Celotto, A.M.; Graveley, B.R. Exon-Specific RNAi: A Tool for Dissecting the Functional Relevance of Alternative Splicing. RNA 2002, 8, 718–724. [Google Scholar] [CrossRef] [Green Version]
- Rappleye, C.A.; Goldman, W.E. Defining Virulence Genes in the Dimporphic Fungi. Annu. Rev. Microbiol. 2006, 60, 281–303. [Google Scholar] [CrossRef]
- Dang, Y.; Yang, Q.; Xue, Z.; Liu, Y. RNA Interference in Fungi: Pathways, Functions, and Applications. Eukaryot. Cell 2011, 10, 1148–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drinnenberg, I.A.; Weinberg, D.E.; Xie, K.T.; Mower, J.P.; Wolfe, K.H.; Fink, G.R.; Bartel, D.P. RNAi in Budding Yeast. Science 2009, 326, 544–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacCallum, D.M.; Castillo, L.; Nather, K.; Munro, C.A.; Brown, A.J.P.; Gow, N.A.R.; Odds, F.C. Property Differences among the Four Major Candida albicans Strain Clades. Eukaryot. Cell 2009, 8, 373–387. [Google Scholar] [CrossRef] [Green Version]
- Pei, Y.; Tuschl, T. On the Art of Identifying Effective and Specific SiRNAs. Nat. Methods 2006, 3, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Kalleda, N.; Naorem, A.; Manchikatla, R.V. Targeting Fungal Genes by Diced SiRNAs: A Rapid Tool to Decipher Gene Function in Aspergillus nidulans. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staab, J.; White, T.; Marr, K. Hairpin DsRNA Does Not Trigger RNA Interference in Candida albicans Cells. Yeast 2011, 28, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, Q.B.; Kadotani, N.; Kasahara, S.; Tosa, Y.; Mayama, S.; Nakayashiki, H. Systematic Functional Analysis of Calcium-Signalling Proteins in the Genome of the Rice-Blast Fungus, Magnaporthe Oryzae, Using a High-Throughput RNA-Silencing System. Mol. Microbiol. 2008, 68, 1348–1365. [Google Scholar] [CrossRef]
- Bourque, G.; Burns, K.H.; Gehring, M.; Gorbunova, V.; Seluanov, A.; Hammell, M.G.; Imbeault, M.; Izsvák, Z.; Levin, H.L.; Macfarlan, T.S.; et al. Ten Things You Should Know about Transposable Elements 06 Biological Sciences 0604 Genetics. Genome Biol. 2018, 19, 1–12. [Google Scholar] [CrossRef]
- Daboussi, M.; Capy, P. Transposable Elements in Filamentous Fungi. Annu. Rev. Microbiol. 2003, 57, 275–299. [Google Scholar] [CrossRef]
- Norman, K.L.; Kumar, A. Mutant Power: Using Mutant Allele Collections for Yeast Functional Genomics. Brief. Funct. Genom. 2016, 15, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Burke, J.E.; Longhurst, A.D.; Natarajan, P.; Rao, B.; Liu, J.; Sales-Lee, J.; Mortensen, Y.; Moresco, J.J.; Diedrich, J.K.; Yates, J.R.; et al. A Non-Dicer RNase III and Four Other Novel Factors Required for RNAi-Mediated Transposon Suppression in the Human Pathogenic Yeast Cryptococcus neoformans. G3 Genes Genomes Genet. 2019, 9, 2235–2244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Zhao, L.; Yan, L.; Zhang, L.; Cao, Y.; Wang, Y.; Jiang, Y.; Yan, T.; Cao, Y. Structural Features and Mechanism of Translocation of Non-LTR Retrotransposons in Candida albicans. Virulence 2014, 5, 245–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segal, E.S.; Gritsenko, V.; Levitan, A.; Yadav, B.; Dror, N.; Steenwyk, J.L.; Silberberg, Y.; Mielich, K.; Rokas, A.; Gow, N.A.R.; et al. Gene Essentiality Analyzed by in Vivo Transposon Mutagenesis and Machine Learning in a Stable Haploid Isolate of Candida albicans. MBio 2018, 9, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Wang, H.; Li, Z.; Wong, A.H.H.; Wang, Y.Z.; Guo, Y.; Lin, X.; Zeng, G.; Wang, Y.; Wang, J. Candida albicans Gains Azole Resistance by Altering Sphingolipid Composition. Nat. Commun. 2018, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.L.; Grahl, N.; Sless, T.; Leach, M.D.; Kim, S.H.; Hogan, D.A.; Robbins, N.; Cowen, L.E. Signaling through Lrg1, Rho1 and Pkc1 Governs Candida albicans Morphogenesis in Response to Diverse Cues. PLoS Genet. 2016, 12, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Barrales, R.R.; Jimenez, J.; Ibeas, J.I. Identification of Novel Activation Mechanisms for FLO11 Regulation in Saccharomyces cerevisiae. Genetics 2008, 178, 145–156. [Google Scholar] [CrossRef] [Green Version]
- Firon, A.; Villalba, F.; Beffa, R.; D’Enfert, C. Identification of Essential Genes in the Human Fungal Pathogen Aspergillus fumigatus by Transposon Mutagenesis. Eukaryot. Cell 2003, 2, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, C.; Hori, Y.; Kashiwagi, Y. Screening and Characterization of Transposon-Insertion Mutants in a Pseudohyphal Strain of Saccharomyces cerevisiae. Yeast 2003, 20, 407–415. [Google Scholar] [CrossRef]
- Uhl, M.A.; Biery, M.; Craig, N.; Johnson, A.D. Haploinsufficiency-Based Large-Scale Forward Genetic Analysis of Filamentous Growth in the Diploid Human Fungal Pathogen C. albicans. EMBO J. 2003, 22, 2668–2678. [Google Scholar] [CrossRef] [Green Version]
- Firon, A.; D’Enfert, C. Identifying Essential Genes in Fungal Pathogens of Humans. Trends Microbiol. 2002, 10, 456–462. [Google Scholar] [CrossRef]
- Xu, T.; Bharucha, N.; Kumar, A. Genome-Wide Transposon Mutagenesis in Saccharomyces cerevisiae and Candida albicans. Methods Mol. Biol. 2011, 765, 207–224. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Seringhaus, M.; Biery, M.C.; Sarnovsky, R.J.; Umansky, L.; Piccirillo, S.; Heidtman, M.; Cheung, K.-H.; Dobry, C.J.; Gerstein, M.B.; et al. Large-Scale Mutagenesis of the Yeast Genome Using a Tn7-Derived Multipurpose Transposon. Genome Res. 2004, 14, 1975–1986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, J.; Fung, E.; Schlecht, U.; Davis, R.W.; Giaever, G.; Onge, R.P.; Deutschbauer, A.; Nislow, C. Gene Annotation and Drug Target Discovery in Candida albicans with a Tagged Transposon Mutant Collection. PLoS Pathog. 2010, 6. [Google Scholar] [CrossRef] [Green Version]
- Horton, B.N.; Kumar, A. Genome-Wide Synthetic Genetic Screening by Transposon Mutagenesis in Candida albicans. Methods Mol. Biol. 2015, 1279, 125–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bharucha, N.; Chabrier-Rosello, Y.; Xu, T.; Johnson, C.; Sobczynski, S.; Song, Q.; Dobry, C.J.; Eckwahl, M.J.; Anderson, C.P.; Benjamin, A.J.; et al. A Large-Scale Complex Haploinsufficiency-Based Genetic Interaction Screen in Candida albicans: Analysis of the Ram Network during Morphogenesis. PLoS Genet. 2011, 7. [Google Scholar] [CrossRef] [Green Version]
- Shoemaker, D.D.; Lashkari, D.A.; Morris, D.; Mittmann, M.; Davis, R.W. Quantitative Phenotypic Analysis of Yeast Deletion Mutants Using a Highly Parallel Molecular Bar-Coding Strategy. Nat. Genet. 1996, 14, 450–456. [Google Scholar] [CrossRef]
- Ross-Macdonald, P.; Coelho, P.S.R.; Roemer, T.; Agarwal, S.; Kumar, A.; Jansen, R.; Cheung, K.-H.; Sheehan, A.; Symoniatis, D.; Umansky, L.; et al. Large-Scale Analysis of the Yeast Genome by Transposon Tagging and Gene Disruption. Nature 1999, 402, 413–418. [Google Scholar] [CrossRef]
- Dias, M.V.; Basso, L.R., Jr.; Coelho, P.S. New Transposons to Generate GFP Protein Fusions in Candida albicans. Gene 2008, 417, 13–18. [Google Scholar] [CrossRef]
- Dufresne, M.; Daboussi, M.J. Development of Impala-Based Transposon Systems for Gene Tagging in Filamentous Fungi. Methods Mol. Biol. 2010, 638, 41–54. [Google Scholar]
- Smith, A.M.; Durbic, T.; Oh, J.; Urbanus, M.; Proctor, M.; Heisler, L.E.; Giaever, G.; Nislow, C. Competitive Genomic Screens of Barcoded Yeast Libraries. J. Vis. Exp. 2011, e2864. [Google Scholar] [CrossRef]
- Nobile, C.J.; Mitchell, A.P. Large-Scale Gene Disruption Using the UAU1 Cassette. Methods Mol. Biol. 2009, 499, 175–194. [Google Scholar] [CrossRef]
- Han, J.S.; Shao, S. Circular Retrotransposition Products Generated by a LINE Retrotransposon. Nucleic Acids Res. 2012, 40, 10866–10877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carr, P.D.; Tuckwell, D.; Hey, P.M.; Simon, L.; d’Enfert, C.; Birch, M.; Oliver, J.D.; Bromley, M.J. The Transposon Impala Is Activated by Low Temperatures: Use of a Controlled Transposition System to Identify Genes Critical for Viability of Aspergillus fumigatus. Eukaryot. Cell 2010, 9, 438–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mielich, K.; Shtifman-Segal, E.; Golz, J.C.; Zeng, G.; Wang, Y.; Berman, J.; Kunze, R. Maize Transposable Elements Ac/Ds as Insertion Mutagenesis Tools in Candida albicans. G3 Genes Genomes Genet. 2018, 8, 1139–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrareze, P.A.; Streit, R.S.; dos Santos, F.M.; Schrank, A.; Kmetzsch, L.; Vainstein, M.H.; Staats, C.C. SRNAs as Possible Regulators of Retrotransposon Activity in Cryptococcus gattii VGII. BMC Genom. 2017, 18, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Wei, D.; Li, Z.; Sun, Z.; Pan, J.; Zhu, X. Cryptococcal Phosphoglucose Isomerase Is Required for Virulence Factor Production, Cell Wall Integrity and Stress Resistance. FEMS Yeast Res. 2015, 15, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Hu, G.; Kronstad, J.W. Gene Disruption in Cryptococcus neoformans and Cryptococcus gattii by in Vitro Transposition. Curr. Genet. 2006, 49, 341–350. [Google Scholar] [CrossRef]
- Batova, M.; Borecka-Melkusova, S.; Simockova, M.; Dzugasova, V.; Goffa, E.; Subik, J. Functional Characterization of the CgPGS1 Gene Reveals a Link between Mitochondrial Phospholipid Homeostasis and Drug Resistance in Candida glabrata. Curr. Genet. 2008, 53, 313–322. [Google Scholar] [CrossRef]
- Juárez-Reyes, A.; De Las Peñas, A.; Castaño, I. Analysis of Subtelomeric Silencing in Candida glabrata. Yeast Genet. Netw. 2011, 734, 279–301. [Google Scholar]
- Page, D.M.; Wittamer, V.; Bertrand, J.Y.; Lewis, K.L.; Pratt, D.N.; Delgado, N.; Schale, S.E.; McGue, C.; Jacobsen, B.H.; Doty, A.; et al. An evolutionarily conserved program of B-cell development and activation in zebrafish. Blood 2013, 122, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Van Der Vaart, M.; Spaink, H.P.; Meijer, A.H. Pathogen recognition and activation of the innate immune response in zebrafish. Adv. Hematol. 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosowski, E.E.; Knox, B.P.; Archambault, L.S.; Huttenlocher, A.; Keller, N.P.; Wheeler, R.T.; Davis, J.M. The zebrafish as a model host for invasive fungal infections. J. Fungi 2018, 4, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosowski, E.E.; Raffa, N.; Knox, B.P.; Golenberg, N.; Keller, N.P.; Huttenlocher, A. Macrophages inhibit Aspergillus fumigatus germination and neutrophil-mediated fungal killing. PLoS Pathog. 2018, 14, 1–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, C.J.; Davis, J.M.; Huttenlocher, A.; Kernien, J.F.; Nett, J.E. Emerging Fungal Pathogen Candida auris Evades Neutrophil Attack. mBio 2018, 21, e01403-18. [Google Scholar]
- Netea, M.G.; Joosten, L.A.B.; Latz, E.; Mills, K.H.G.; Natoli, G.; Stunnenberg, H.G.; O’Neill, L.A.J.; Xavier, R.J. Trained immunity: A program of innate immune memory in health and disease. Science (80-) 2016, 352, 427. [Google Scholar] [CrossRef] [Green Version]
- Kawai, Y.; Mickiewicz, K.; Errington, J. Lysozyme Counteracts β-Lactam Antibiotics by Promoting the Emergence of L-Form Bacteria. Cell 2018, 172, 1038–1049. [Google Scholar] [CrossRef] [Green Version]
- Mickiewicz, K.M.; Kawai, Y.; Drage, L.; Gomes, M.C.; Davison, F.; Pickard, R.; Hall, J.; Mostowy, S.; Aldridge, P.D.; Errington, J. Possible role of L-form switching in recurrent urinary tract infection. Nat. Commun. 2019, 10, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Negus, D.; Moore, C.; Baker, M.; Raghunathan, D.; Tyson, J.; Sockett, R.E. Predator Versus Pathogen: How Does Predatory Bdellovibrio bacteriovorus Interface with the Challenges of Killing Gram-Negative Pathogens in a Host Setting? Annu. Rev. Microbiol. 2017, 71, 441–457. [Google Scholar] [CrossRef]
- Willis, A.R.; Moore, C.; Mazon-Moya, M.; Krokowski, S.; Lambert, C.; Till, R.; Mostowy, S.; Sockett, R.E. Injections of Predatory Bacteria Work Alongside Host Immune Cells to Treat Shigella Infection in Zebrafish Larvae. Curr. Biol. 2016, 26, 3343–3351. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.L.; Upadhyayula, S.; Milkie, D.E.; Singh, V.; Wang, K.; Swinburne, I.A.; Mosaliganti, K.R.; Collins, Z.M.; Hiscock, T.W.; Shea, J.; et al. Observing the cell in its native state: Imaging subcellular dynamics in multicellular organisms. Science (80-) 2018, 360, eaaq1392. [Google Scholar] [CrossRef] [Green Version]
- Ishii, M.; Matsumoto, Y.; Sekimizu, K. Usefulness of silkworm as a host animal for understanding pathogenicity of Cryptococcus neoformans. Drug Discov. Ther. 2016, 10, 9–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaito, C.; Akimitsu, N.; Watanabe, H.; Sekimizu, K. Silkworm larvae as an animal model of bacterial infection pathogenic to humans. Microb. Pathog. 2002, 32, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Kaito, C.; Kurokawa, K.; Matsumoto, Y.; Terao, Y.; Kawabata, S.; Hamada, S.; Sekimizu, K. Silkworm pathogenic bacteria infection model for identification of novel virulence genes. Mol. Microbiol. 2005, 56, 934–944. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, S.; Matsumoto, Y.; Sekimizu, K.; Kaito, C. Evaluation of Staphylococcus aureus virulence factors using a silkworm model. FEMS Microbiol. Lett. 2012, 326, 116–124. [Google Scholar] [CrossRef] [Green Version]
- Hamamoto, H.; Kurokawa, K.; Kaito, C.; Kamura, K.; Razanajatovo, I.M.; Kusuhara, H.; Santa, T.; Sekimizu, K. Quantitative Evaluation of the Therapeutic Effects of Antibiotics Using Silkworms Infected with Human Pathogenic Microorganisms. Antimicrob. Agents Chemother. 2004, 48, 774–779. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, Y.; Sumiya, E.; Sugita, T.; Sekimizu, K. An invertebrate hyperglycemic model for the identification of anti-diabetic drugs. PLoS ONE 2011, 6, e18292. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, Y.; Miyazaki, S.; Fukunaga, D.H.; Shimizu, K.; Kawamoto, S.; Sekimizu, K. Quantitative evaluation of cryptococcal pathogenesis and antifungal drugs using a silkworm infection model with Cryptococcus neoformans. J. Appl. Microbiol. 2012, 112, 138–146. [Google Scholar] [CrossRef]
- Hanaoka, N.; Takano, Y.; Shibuya, K.; Fugo, H.; Uehara, Y.; Niimi, M. Identification of the putative protein phosphatase gene PTC1 as a virulence-related gene using a silkworm model of Candida albicans infection. Eukaryot. Cell 2008, 7, 1640–1648. [Google Scholar] [CrossRef] [Green Version]
- Ueno, K.; Matsumoto, Y.; Uno, J.; Sasamoto, K.; Sekimizu, K.; Kinjo, Y.; Chibana, H. Intestinal resident yeast Candida glabrata requires Cyb2p-Mediated lactate assimilation to adapt in mouse intestine. PLoS ONE 2011, 6, e24759. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Nielsen, K.; Patel, S.; Heitman, J. Impact of mating type, serotype, and ploidy on the virulence of Cryptococcus neoformans. Infect. Immun. 2008, 76, 2923–2938. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, I.; Kanasaki, R.; Yoshikawa, K.; Furukawa, S.; Fujie, A.; Hamamoto, H.; Sekimizu, K. Discovery of a new antifungal agent ASP2397 using a silkworm model of Aspergillus fumigatus infection. J. Antibiot 2017, 70, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Glittenberg, M.T.; Silas, S.; MacCallum, D.M.; Gow, N.A.; Ligoxygakis, P. Wild-type Drosophila melanogaster as an alternative model system for investigating the pathogenicity of Candida albicans. Dis. Model. Mech. 2011, 4, 504–514. [Google Scholar] [CrossRef] [Green Version]
- Pereira, T.C.; De Barros, P.P.; Fugisaki, L.R.D.O.; Rossoni, R.D.; Ribeiro, F.D.C.; De Menezes, R.T.; Junqueira, J.C.; Scorzoni, L. Recent advances in the use of Galleria mellonella model to study immune responses against human pathogens. J. Fungi 2018, 4, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banville, N.; Browne, N.; Kavanagh, K. Effect of nutrient deprivation on the susceptibility of Galleria mellonella larvae to infection. Virulence 2012, 3, 497–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mylonakis, E. Galleria mellonella and the study of fungal pathogenesis: Making the case for another genetically tractable model host. Mycopathologia 2008, 165, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Kavanagh, K.; Reeves, E.P. Exploiting the potential of insects for in vivo pathogenicity testing of microbial pathogens. FEMS Microbiol. Rev. 2004, 28, 101–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotter, G.; Doyle, S.; Kavanagh, K. Development of an insect model for the in vivo pathogenicity testing of yeasts. FEMS Immunol. Med. Microbiol. 2000, 27, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Lionakis, M.S.; Lewis, R.E.; May, G.S.; Wiederhold, N.P.; Albert, N.D.; Halder, G.; Kontoyiannis, D.P. Toll- Deficient Drosophila Flies as a Fast, High-Throughput Model for the Study of Antifungal Drug Efficacy against Invasive Aspergillosis and Aspergillus Virulence. J. Infect. Dis. 2005, 191, 1188–1195. [Google Scholar] [CrossRef] [Green Version]
- Brennan, M.; Thomas, D.Y.; Whiteway, M.; Kavanagh, K. Correlation between virulence of Candida albicans mutants in mice and Galleria mellonella larvae. FEMS Immunol. Med. Microbiol. 2002, 34, 153–157. [Google Scholar] [CrossRef] [Green Version]
- Kavanagh, K.; Sheehan, G. The use of Galleria mellonella larvae to identify novel antimicrobial agents against fungal species of medical interest. J. Fungi 2018, 4, 113. [Google Scholar] [CrossRef] [Green Version]
- Bergin, D.; Murphy, L.; Keenan, J.; Clynes, M.; Kavanagh, K. Pre-exposure to yeast protects larvae of Galleria mellonella from a subsequent lethal infection by Candida albicans and is mediated by the increased expression of antimicrobial peptides. Microbes Infect. 2006, 8, 2105–2112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, G.D. Innate Antifungal Immunity: The Key Role of Phagocytes. Annu. Rev. Immunol. 2011, 29, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sable, C.A.; Strohmaier, K.M.; Chodakewitz, J.A. Recent advances in antifungal therapy. Thorax 1974, 29, 615. [Google Scholar]
- Steele, C.; Wormley, F.L. Immunology of fungal infections: Lessons learned from animal models. Curr. Opin. Microbiol. 2012, 15, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Breschi, A.; Gingeras, T.R.; Guigó, R. Comparative transcriptomics in human and mouse. Nat. Rev. Genet. 2017, 18, 425–440. [Google Scholar] [CrossRef]
- Chinwalla, A.; Cook, L.; Delehaunty, K.; Fewell, G.; Fulton, L.; Fulton, R.; Graves, T.; Hillier, L.; Mardis, E.; McPherson, J. Initial sequencing and comparative analysis of the mouse genome. Nature 2002, 420, 520–562. [Google Scholar]
- Desoubeaux, G.; Cray, C. Animal models of aspergillosis. Comp. Med. 2018, 68, 109–123. [Google Scholar]
- Capilla, J.; Clemons, K.V.; Stevens, D.A. Animal models: An important tool in mycology. Med. Mycol. 2007, 45, 657–684. [Google Scholar] [CrossRef] [Green Version]
- Rauseo, A.M.; Coler-Reilly, A.; Larson, L.; Spec, A. Hope on the Horizon: Novel Fungal Treatments in Development. Open Forum Infect. Dis. 2020, 7, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Pande, K.; Chen, C.; Noble, S.M. Passage through the mammalian gut triggers a phenotypic switch that promotes Candida albicans commensalism. Nat. Genet. 2013, 45, 1088–1091. [Google Scholar] [CrossRef] [Green Version]
- Prieto, D.; Pla, J. Distinct stages during colonization of the mouse gastrointestinal tract by Candida albicans. Front. Microbiol. 2015, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mayfield, J.A.; Rine, J. The genetic basis of variation in susceptibility to infection with Histoplasma capsulatum in the mouse. Genes Immun. 2007, 8, 468–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radovanovic, I.; Mullick, A.; Gros, P. Genetic control of susceptibility to infection with Candida albicans in mice. PLoS ONE 2011, 6, e18957. [Google Scholar] [CrossRef] [Green Version]
- Xin, H.; Mohiuddin, F.; Tran, J.; Adams, A.; Eberle, K. Experimental Mouse Models of Disseminated Candida auris Infection. mSphere 2019, 4, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaragoza, O.; Alvarez, M.; Telzak, A.; Rivera, J.; Casadevall, A. The relative susceptibility of mouse strains to pulmonary Cryptococcus neoformans infection is associated with pleiotropic differences in the immune response. Infect. Immun. 2007, 75, 2729–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balloy, V.; Huerre, M.; Latgé, J.P.; Chignard, M. Differences in patterns of infection and inflammation for corticosteroid treatment and chemotherapy in experimental invasive pulmonary aspergillosis. Infect. Immun. 2005, 73, 494–503. [Google Scholar] [CrossRef] [Green Version]
- Spellberg, B.; Ibrahim, A.S.; Edwards, J.E., Jr.; Filler, S.G. Mice with Disseminated Candidiasis Die of Progressive Sepsis. J. Infect. Dis. 2005, 192, 336–343. [Google Scholar] [CrossRef]
- Ngo, L.Y.; Kasahara, S.; Kumasaka, D.K.; Knoblaugh, S.E.; Jhingran, A.; Hohl, T.M. Inflammatory monocytes mediate early and organ-specific innate defense during systemic candidiasis. J. Infect. Dis. 2014, 209, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Machholz, E.; Mulder, G.; Ruiz, C.; Corning, B.F.; Pritchett-Corning, K.R. Manual restraint and common compound administration routes in mice and rats. J. Vis. Exp. 2012, 26, e2771. [Google Scholar] [CrossRef] [Green Version]
- Da Glória Sousa, M.; Reid, D.M.; Schweighoffer, E.; Tybulewicz, V.; Ruland, J.; Langhorne, J.; Yamasaki, S.; Taylor, P.R.; Almeida, S.R.; Brown, G.D. Restoration of pattern recognition receptor costimulation to treat chromoblastomycosis, a chronic fungal infection of the skin. Cell Host Microbe 2011, 9, 436–443. [Google Scholar] [CrossRef]
- Paisley, D.; Robson, G.D.; Denning, D.W. Correlation between in vitro growth rate and in vivo virulence in Aspergillus fumigatus. Med. Mycol. 2005, 43, 397–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhodes, J.C. Aspergillus fumigatus: Growth and virulence. Med. Mycol. 2006, 44, 77–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marakalala, M.J.; Vautier, S.; Potrykus, J.; Walker, L.A.; Shepardson, K.M.; Hopke, A.; Mora-Montes, H.M.; Kerrigan, A.; Netea, M.G.; Murray, G.I.; et al. Differential Adaptation of Candida albicans in Vivo Modulates Immune Recognition by Dectin-1. PLoS Pathog. 2013, 9, 1–12. [Google Scholar] [CrossRef]
- Nagy, G.; Szebenyi, C.; Csernetics, Á.; Vaz, A.G.; Judit, E.; Vágvölgyi, C.; Papp, T. Development of a plasmid free CRISPR-Cas9 system for the genetic modification of Mucor circinelloides. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Denning, D.; Anderson, M.J.; Turner, G.; Latge, J.P.; Bennett, J. Sequencing the Aspergillus fumigatus genome. Lancet 2002, 2, 251–253. [Google Scholar] [CrossRef]
- Wa, S.; State, W. Improved Reference Genome Sequence of Coccidioides immitis. Microbiol Resour Announc 2019, 8, 18–20. [Google Scholar]
- Cbs, L.L.S. Complete Genome Sequence of a High Lipid-Producing Strain of Mucor circinelloides WJ11 and Comparative Genome Analysis with a low lipid-producing strain CBS 277.49. PLoS ONE 2015, 10, e0137543. [Google Scholar]
- Voorhies, M.; Foo, C.K.; Sil, A. Experimental annotation of the human pathogen Histoplasma capsulatum transcribed regions using high-resolution tiling arrays. BMC Microbiol. 2011, 11, 216. [Google Scholar] [CrossRef] [Green Version]
- Inglis, D.O.; Arnaud, M.B.; Binkley, J.; Shah, P.; Skrzypek, M.S.; Wymore, F.; Binkley, G.; Miyasato, S.R.; Simison, M.; Sherlock, G. The Candida genome database incorporates multiple Candida species: Multispecies search and analysis tools with curated gene and protein information for Candida albicans and Candida glabrata. Nucleic Acids Res. 2012, 40, 667–674. [Google Scholar] [CrossRef]
- Vyas, V.K.; Barrasa, M.I.; Fink, G.R. A Candida albicans CRISPR system permits genetic engineering of essential genes and gene families. Sci. Adv. 2015, 1, e1500248. [Google Scholar] [CrossRef] [Green Version]
- Nicolás, F.E.; Vila, A.; Moxon, S.; Cascales, M.D.; Torres-Martínez, S.; Ruiz-Vázquez, R.M.; Garre, V. The RNAi machinery controls distinct responses to environmental signals in the basal fungus Mucor circinelloides. BMC Genom. 2015, 16, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neafsey, D.E.; Barker, B.M.; Sharpton, T.J.; Stajich, J.E.; Park, D.J.; Whiston, E.; Hung, C.; Mcmahan, C.; White, J.; Sykes, S.; et al. Population genomic sequencing of Coccidioides fungi reveals recent hybridization and transposon control. Genome Res. 2010, 20, 938–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enkler, L.; Richer, D.; Marchand, A.L.; Ferrandon, D. Genome engineering in the yeast pathogen Candida glabrata using the CRISPR-Cas9 system. Sci. Rep. 2016, 6, 35766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, L.; Voorhies, M.; Gilmore, S.; Beyhan, S.; Myint, A.; Id, A.S. Opposing signaling pathways regulate morphology in response to temperature in the fungal pathogen Histoplasma capsulatum. PLoS Biol. 2019, 17, e3000168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, G.; Grace, A.; Szebenyi, C.; Takó, M.; Tóth, E.J.; Csernetics, Á.; Bencsik, O.; Szekeres, A.; Homa, M.; Ayaydin, F. CRISPR-Cas9-mediated disruption of the HMG-CoA reductase genes of Mucor circinelloides and subcellular localization of the encoded enzymes. Fungal Genet. Biol. 2019, 129, 30–39. [Google Scholar] [CrossRef]
- Mouyna, I.; Henry, C.; Doering, T.L.; Latge, J. Gene silencing with RNA interference in the human pathogenic fungus Aspergillus fumigatus. FEMS Microbiol. Lett. 2004, 237, 317–324. [Google Scholar] [CrossRef] [Green Version]
- Loftus, B.J.; Fung, E.; Roncaglia, P.; Rowley, D.; Amedeo, P.; Vamathevan, J.; Miranda, M.; Anderson, I.J.; Fraser, J.A.; Allen, J.E.; et al. The Genome of the Basidiomycetous Yeast and Human pathogen Cryptococcus neoformans. Science 2012, 307, 1321–1324. [Google Scholar] [CrossRef] [Green Version]
- Wüthrich, M.; Klein, S. CRISPR/Cas9-Mediated Gene Disruption Reveals the Fungal Pathogen Blastomyces dermatitidis. MBio 2018, 9, e00412-18. [Google Scholar]
- Tsai, H.; Bard, M.; Izumikawa, K.; Krol, A.A.; Sturm, A.M.; Culbertson, N.T.; Pierson, C.A.; Bennett, J.E. Candida glabrata erg1 Mutant with Increased Sensitivity to Azoles and to Low Oxygen Tension. Antimicrob. Agents Chemother. 2004, 48, 2483–2489. [Google Scholar] [CrossRef] [Green Version]
- Mu, J.F.; Gauthier, G.M.; Desjardins, C.A.; Gallo, J.E.; Holder, J.; Sullivan, T.D.; Marty, A.J.; Carmen, J.C.; Chen, Z.; Ding, L.; et al. The Dynamic Genome and Transcriptome of the Human Fungal Pathogen Blastomyces and Close Relative Emmonsia. PLoS Genet. 2015, 11, e1005493. [Google Scholar]
- Min, K.; Ichikawa, Y.; Woolford, C.A.; Mitchell, A.P. Candida albicans Gene Deletion with a Transient CRISPR-Cas9 System. mSphere 2016, 1, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nosanchuk, J.D.; Steenbergen, J.N.; Shi, L.; Deepe, G.S.; Casadevall, A. Antibodies to a cell surface histone-like protein protect against Histoplasma capsulatum. J. Clin. Invesigt. 2003, 112, 1164–1175. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, K.N.; Clemons, K.V.; Stevens, D.A. Murine models of blastomycosis, coccidioidomycosis, and histoplasmosis. Mycopathologia 1999, 146, 53–65. [Google Scholar] [CrossRef]
- Hamilos, G.; Samonis, G.; Kontoyiannis, D.P. Recent advances in the use of Drosophila melanogaster as a model to study immunopathogenesis of medically important filamentous fungi. Int. J. Microbiol. 2012, 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apidianakis, Y.; Rahme, L.G.; Heitman, J.; Ausubel, F.M.; Calderwood, S.B.; Mylonakis, E. Challenge of Drosophila melanogaster with Cryptococcus neoformans and Role of the Innate Immune Response. Eukaryot. Cell 2004, 3, 413–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ames, L.; Duxbury, S.; Pawlowska, B.; Ho, H.L.; Haynes, K.; Bates, S. Galleria mellonella as a host model to study Candida glabrata virulence and antifungal efficacy. Virulence 2017, 8, 1909–1917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trevijano-Contador, N.; Zaragoza, O. Immune response of Galleria mellonella against human fungal pathogens. J. Fungi 2019, 5, 3. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, I.D. Animal models to study mucormycosis. J. Fungi 2019, 5, 27. [Google Scholar] [CrossRef] [Green Version]
- Quintin, J.; Asmar, J.; Matskevich, A.A.; Lafarge, M.-C.; Ferrandon, D. The Drosophila Toll Pathway Controls but Does Not Clear Candida glabrata Infections. J. Immunol. 2013, 190, 2818–2827. [Google Scholar] [CrossRef] [Green Version]
- Clemons, K.V.; Capilla, J.; Stevens, D.A. Experimental animal models of coccidioidomycosis. Ann. N. Y. Acad. Sci. 2007, 1111, 208–224. [Google Scholar] [CrossRef]
- Peleg, A.Y.; Jara, S.; Monga, D.; Eliopoulos, G.M.; Moellering, R.C.; Mylonakis, E. Galleria mellonella as a model system to study Acinetobacter baumannii pathogenesis and therapeutics. Antimicrob. Agents Chemother. 2009, 53, 2605–2609. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Tools and Resources | Candida albicans | Candida glabrata | Aspergillus fumigatus | Cryptococcus neoformans | Histoplasma capsulatum | Coccidiodes immitis | Blastomyces dermatitidis | Mucor circillenoides |
|---|---|---|---|---|---|---|---|---|
| Sequenced and annotated genome | [194] | [194] | [195] | [196] | [197] | [198] | [199] | [193] |
| CRISPR | [195,200,201] | [202] | [16] | [18] | [203,204] | [205] | [206] | [207,208] |
| RNAi | [75,80] | [85] | [209] | [21,76,84] | [82] | [89] | [78] | [201] |
| Transposon mutagenesis | [112,123] | [210] | [116,132] | [110,136] | X | [211] | X | X |
| Animal Models | ||||||||
| Zebrafish | [140,145] | [141] | [141,142] | [143] | [212] | [212,213] | [16] | X |
| Bombyx mori—the silkworm | [157] | [155,159] | [160] | [150,156] | X | X | X | X |
| Galleria mellonella—the wax moth | [168,170,214] | [215,216] | [165,214] | [214,217] | [214,217] | X | X | [218] |
| Drosophila melanogaster | [161] | [219] | [167] | [220] | X | X | X | [90] |
| Murine model | [168,179,180,182,186] | [173,177,186] | [176,185] | [184] | [221] | [213] | X | [90] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malavia, D.; Gow, N.A.R.; Usher, J. Advances in Molecular Tools and In Vivo Models for the Study of Human Fungal Pathogenesis. Microorganisms 2020, 8, 803. https://doi.org/10.3390/microorganisms8060803
Malavia D, Gow NAR, Usher J. Advances in Molecular Tools and In Vivo Models for the Study of Human Fungal Pathogenesis. Microorganisms. 2020; 8(6):803. https://doi.org/10.3390/microorganisms8060803
Chicago/Turabian StyleMalavia, Dhara, Neil A. R. Gow, and Jane Usher. 2020. "Advances in Molecular Tools and In Vivo Models for the Study of Human Fungal Pathogenesis" Microorganisms 8, no. 6: 803. https://doi.org/10.3390/microorganisms8060803
APA StyleMalavia, D., Gow, N. A. R., & Usher, J. (2020). Advances in Molecular Tools and In Vivo Models for the Study of Human Fungal Pathogenesis. Microorganisms, 8(6), 803. https://doi.org/10.3390/microorganisms8060803